

Dietary Hydroxytyrosol Supplementation on Growth Performance, Gut Morphometry, and Oxidative and Inflammatory Status in LPS-Challenged Broilers
 , , , ,
, , , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Experimental Design, and Diets
2.2. Growth Performance
2.3. Samples Collection
2.3.1. Relative Organ Weights
2.3.2. Intestinal Morphometry
2.3.3. mRNA Extraction and Gene Expression
2.4. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Relative Organ Weight
3.3. Intestinal Morphometry
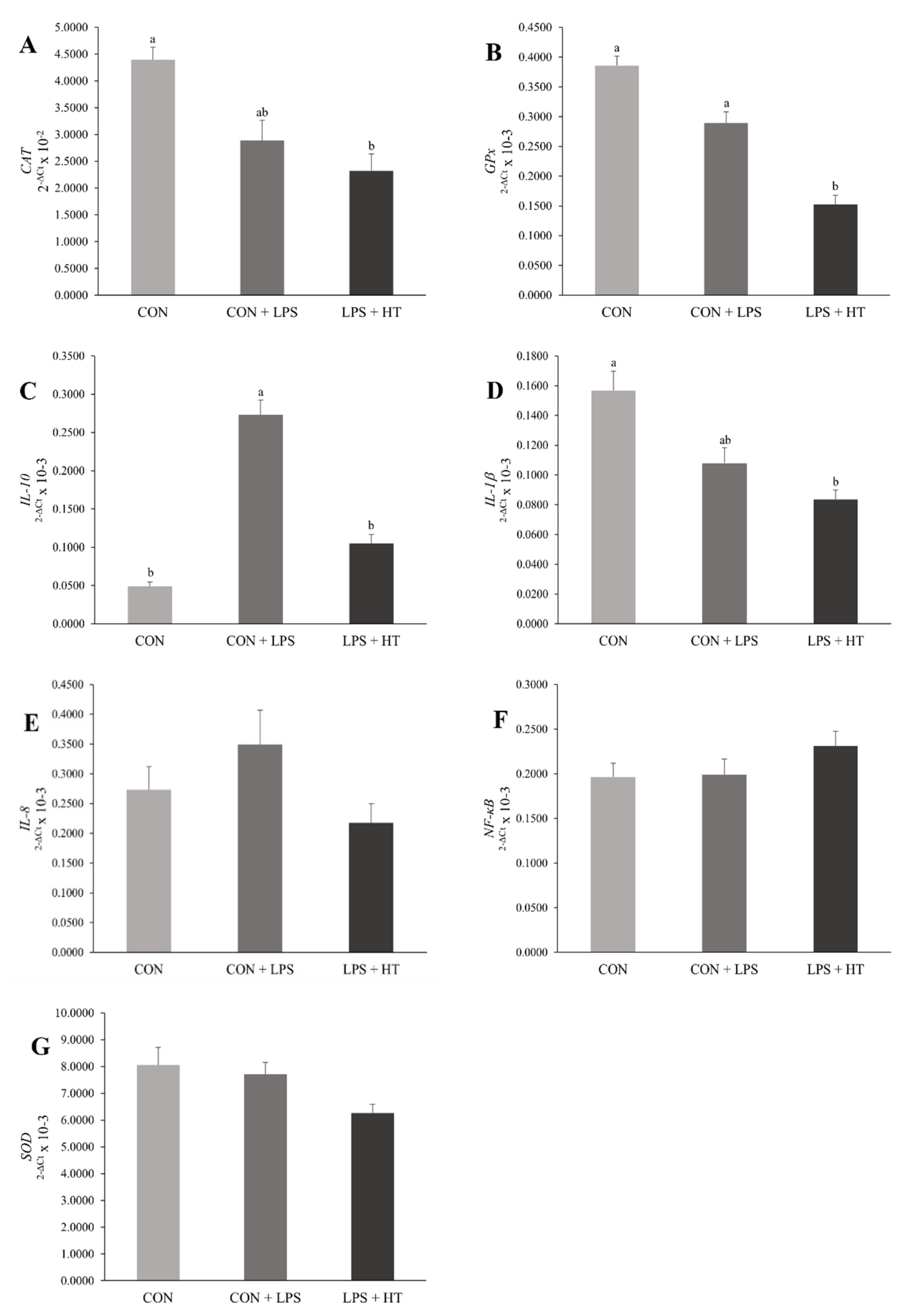
3.4. mRNA Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, J.; Wang, H.; Wu, Z.; Gu, H.; Li, C.; Wang, S.; Liu, G. Effects of 5-aminolevulinic acid on the inflammatory responses and antioxidative capacity in broiler chickens challenged with lipopolysaccharide. Animal 2022, 16, 100575. [Google Scholar] [CrossRef]
- Kong, L.; Wang, Z.; Xiao, C.; Zhu, Q.; Song, Z. Glycerol monolaurate attenuated immunological stress and intestinal mucosal injury by regulating the gut microbiota and activating AMPK/Nrf2 signaling pathway in lipopolysaccharide-challenged broilers. Anim. Nutr. 2022, 10, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ye, J.; Zhang, S.; Chen, Z.; Fan, Q.; Jiang, S. Dietary supplementation with anthocyanin attenuates lipopolysaccharide-induced intestinal damage through antioxidant effects in yellow-feathered broiler chicks. Poult. Sci. 2022, 102, 102325. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, G.; Liang, X.; Wang, M.; Zhu, X.; Luo, Y.; Shang, Y.; Yang, J.-Q.; Zhou, P.; Gu, X.-L. Effects of berberine on the growth performance, antioxidative capacity and immune response to lipopolysaccharide challenge in broilers. Anim. Sci. J. 2019, 90, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Han, H.; Zhang, L.; Wang, T. Dietary bisdemethoxycurcumin supplementation attenuates lipopolysaccharide-induced damages on intestinal redox potential and redox status of broilers. Poult. Sci. 2021, 100, 101061. [Google Scholar] [CrossRef] [PubMed]
- Bi, S.; Shao, J.; Qu, Y.; Hu, W.; Ma, Y.; Cao, L. Hepatic transcriptomics and metabolomics indicated pathways associated with immune stress of broilers induced by lipopolysaccharide. Poult. Sci. 2022, 101, 102199. [Google Scholar] [CrossRef] [PubMed]
- Shan, C.; Miao, F. Immunomodulatory and antioxidant effects of hydroxytyrosol in cyclophosphamide-induced immunosuppressed broilers. Poult. Sci. 2022, 101, 101516. [Google Scholar] [CrossRef] [PubMed]
- Britton, J.; Davis, R.; O’Connor, K.E. Chemical, physical and biotechnological approaches to the production of the potent antioxidant hydroxytyrosol. Appl. Microbiol. Biotechnol. 2019, 103, 5957–5974. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sun, X.; Chai, X.; Jiao, Y.; Sun, J.; Wang, S.; Wang, S.; Yu, H.; Feng, X. Curcumin mitigates oxidative damage in broiler liver and ileum caused by Aflatoxin B1-contaminated feed through Nrf2 signaling pathway. Animals 2024, 14, 409. [Google Scholar] [CrossRef]
- Yang, H.; Wang, Y.; Liu, M.; Liu, X.; Jiao, Y.; Jin, S.; Shan, A.; Feng, X. Effects of dietary resveratrol supplementation on growth performance and anti-inflammatory ability in ducks (Anas platyrhynchos) through the Nrf2/HO-1 and TLR4/NF-κB signaling pathways. Animals 2021, 11, 3588. [Google Scholar] [CrossRef]
- Yang, H.; Wang, Y.; Jin, S.; Pang, Q.; Shan, A.; Feng, X. Dietary resveratrol alleviated lipopolysaccharide-inducted ileitis through Nrf2 and NK-κB signalling pathways in ducks (Anas platyrhynchos). J. Anim. Physiol. Anim. Nutr. 2022, 106, 1306–1320. [Google Scholar] [CrossRef]
- Bertelli, M.; Kiani, A.K.; Paolacci, S.; Manara, E.; Kurti, D.; Dhuli, K.; Bushati, V.; Miertus, J.; Pangallo, D.; Baglivo, M.; et al. Hydroxytyrosol: A natural compound with promising pharmacological activities. J. Biotechnol. 2020, 309, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Zrelli, H.; Matsuoka, M.; Kitazaki, S.; Araki, M.; Kusunoki, M.; Zarrouk, M.; Miyazaki, H. Hydroxytyrosol induces proliferation and cytoprotection against oxidative injury in vascular endothelial cells: Role of Nrf2 activation and HO-1 induction. J. Agric. Food Chem. 2011, 59, 4473–4482. [Google Scholar] [CrossRef] [PubMed]
- Robles-Almazan, M.; Pulido-Moran, M.; Moreno-Fernandez, J.; Ramirez-Tortosa, C.; Rodriguez-Garcia, C.; Quiles, J.L.; Ramirez-Tortosa, M.C. Hydroxytyrosol: Bioavailability, toxicity, and clinical applications. Int. Food Res. J. 2018, 105, 654–667. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, W.; Tian, X.; He, Y.; Xiao, Z.; Zhao, X.; Fan, L.; Du, R.; Yang, G.; Yu, T. Hydroxytyrosol effectively improves the quality of pig sperm at 17 °C. Theriogenology 2022, 177, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Branciari, R.; Galarini, R.; Giusepponi, D.; Trabalza-Marinucci, M.; Forte, C.; Roila, R.; Miraglia, D.; Servili, M.; Acuti, G.; Valiani, A. Oxidative status and presence of bioactive compounds in meat from chickens fed polyphenols extracted from olive oil industry waste. Sustainability 2017, 9, 1566. [Google Scholar] [CrossRef]
- Sabino, M.; Cappelli, K.; Capomaccio, S.; Pascucci, L.; Biasato, I.; Verini-Supplizi, A.; Valiani, A.; Trabalza-Marinucci, M. Dietary supplementation with olive mill wastewaters induces modifications on chicken jejunum epithelial cell transcriptome and modulates jejunum morphology. BMC Genom. 2018, 19, 576. [Google Scholar] [CrossRef]
- Pappas, A.C.; Tsiplakou, E.; Papadomichelakis, G.; Mitsiopoulou, C.; Sotirakoglou, K.; Mpekelis, V.; Haroutounian, S.A.; Fegeros, K.; Zervas, G. Effects of olive pulp addition to broiler diets on performance, selected biochemical parameters and antioxidant enzymes. J. Hellenic Vet. Med. Soc. 2019, 70, 1687–1696. [Google Scholar] [CrossRef]
- Papadomichelakis, G.; Pappas, A.C.; Tsiplakou, E.; Symeon, G.K.; Sotirakoglou, K.; Mpekelis, V.; Fegeros, K.; Zervas, G. Effects of dietary dried olive pulp inclusion on growth performance and meat quality of broiler chickens. Livest. Sci. 2019, 221, 115–122. [Google Scholar] [CrossRef]
- Dias, K.M.M.; Oliveira, C.H.; Calderano, A.A.; Rostagno, H.S.; O’Connor, K.E.; Davis, R.; Walsh, M.; Britton, J.; Altieri, E.A.; Albino, L.F.T. Effects of hydroxytyrosol supplementation on performance, meat quality and blood parameters of broiler chickens. Animals 2023, 14, 119. [Google Scholar] [CrossRef]
- Rostagno, H.S.; Albino, L.F.T.; Hannas, M.I.; Donzele, J.L.; Sakomura, N.K.; Perazzo, F.G.; Saraiva, A.; de Abreu, M.L.T.; Rodrigues, P.B.; de Oliveira, R.F.; et al. Brazilian Tables for Poultry and Swine: Composition of Feedstuffs and Nutritional Requirements, 4th ed.; Federal University of Viçosa: Viçosa, Brazil, 2017; 482p. [Google Scholar]
- Gava, M.S. Metodologia de Morfometria Intestinal em Frango de Corte. Master’s Thesis, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil, 2012. [Google Scholar]
- Luna, G.L. Manual of Histologic Staining Methods of the Armed Forces Institute of Pathology, 3rd ed.; Mc Graw-Hill Book Company: London, UK, 1968. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2011, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, Z.; Ren, W.; Yue, Y.; Guo, Y. Effects of live yeast supplementation on lipopolysaccharide-induced inflammatory responses in broilers. Poult. Sci. 2016, 95, 2557–2564. [Google Scholar] [CrossRef]
- Zhu, C.; Nie, X.; He, Z.; Xiong, T.; Li, Y.; Bai, Y.; Zhang, H. Research Note: Dietary resveratrol supplementation improves the hepatic antioxidant capacity and attenuates lipopolysaccharide-induced inflammation in yellow-feathered broilers. Poult. Sci. 2023, 102, 102370. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Qi, L.; Wei, Q.; Shi, F. Maternal stevioside supplementation ameliorates intestinal mucosal damage and modulates gut microbiota in the chick offspring challenged with lipopolysaccharide. Food Funct. 2021, 12, 6014–6028. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cheng, Y.; Wang, W.; Wang, A.; Zhou, Y. Protective effects of dietary supplementation with a silicate clay mineral (palygorskite) in lipopolysaccharide-challenged broiler chickens at an early age. Anim. Feed. Sci. Techonol. 2020, 263, 114459. [Google Scholar] [CrossRef]
- Lin, J.-Y.; Lai, Y.-S.; Liu, C.-J.; Wu, A.-R. Effects of lotus plumule supplementation before and following systemic administration of lipopolysaccharide on the splenocyte responses of BALB/c mice. Food Chem. Toxicol. 2007, 45, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, G.; Zhu, X.; Luo, Y.; Shang, Y.; Gu, X.-L. The anti-inflammatory and antioxidant effects of leonurine hydrochloride after lipopolysaccharide challenge in broiler chicks. Poult. Sci. 2019, 98, 1648–1657. [Google Scholar] [CrossRef]
- Nunes, R.A.; Duarte, M.S.; Campos, P.H.R.F.; Oliveira, L.L.; Silva, F.F.; Kreuz, B.S.; Mirabile, C.G.; Borges, S.O.; Calderano, A.A. Active vitamin D3-glycoside preserves weight gain and modulates the inflammatory response in broiler chickens challenged with lipopolysaccharide. Anim. Feed Sci. Technol. 2020, 270, 114704. [Google Scholar] [CrossRef]
- Jiang, J.; Qi, L.; Lv, Z.; Jin, S.; Wei, X.; Shi, F. Dietary stevioside supplementation alleviates lipopolysaccharide-induced intestinal mucosal damage through anti-inflammatory and antioxidant effects in broiler chickens. Antioxidants 2019, 8, 575. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, X.; Eicher, S.D.; Ajuwon, K.M.; Applegate, T.J. Effect of threonine on secretory immune system using a chicken intestinal ex vivo model with lipopolysaccharide challenge. Poult. Sci. 2017, 96, 3043–3051. [Google Scholar] [CrossRef]
- Chang, Y.; Yuan, L.; Liu, J.; Muhammad, I.; Cao, C.; Shi, C.; Zhang, Y.; Li, R.; Li, C.; Liu, F. Dihydromyricetin attenuates Escherichia coli lipopolysaccharide-induced ileum injury in chickens by inhibiting NLRP3 inflammasome and TLR4/NF-κB signalling pathway. Vet. Res. 2020, 51, 72. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, J.; Hou, Y.; Zhu, H.; Zhao, S.; Ding, B.; Yin, Y.; Yi, G.; Shi, J.; Fan, W. Dietary arginine supplementation alleviates intestinal mucosal disruption induced by Escherichia coli lipopolysaccharide in weaned pigs. Br. J. Nutr. 2008, 100, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, H.; Chen, Y.P.; Yang, M.X.; Zhang, L.L.; Lu, Z.X.; Zhou, Y.M.; Wang, T. Bacillus amyloliquefaciens supplementation alleviates immunological stress and intestinal damage in lipopolysaccharide-challenged broilers. Anim. Feed Sci. Technol. 2015, 208, 119–131. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, H.J.; Yu, S.H.; Wu, S.G.; Yoon, I.; Quigley, J.; Gao, Y.P.; Qi, G.H. Effects of yeast culture in broiler diets on performance and immunomodulatory functions. Poult. Sci. 2008, 87, 1377–1384. [Google Scholar] [CrossRef] [PubMed]
- Montagne, L.; Pluske, J.R.; Hampson, D.J. A review of interactions between dietary fiber and the intestinal mucosa, and their consequences on digestive health in young non-ruminant animals. Anim. Feed Sci. Technol. 2003, 108, 95–117. [Google Scholar] [CrossRef]
- Yang, L.; Liu, G.; Lian, K.; Qiao, Y.; Zhang, B.; Zhu, X.; Luo, Y.; Shang, Y.; Gu, X.-L. Dietary leonurine hydrochloride supplementation attenuates lipopolysaccharide challenge-induced intestinal inflammation and barrier dysfunction by inhibiting the NF-κB/MAPK signaling pathway in broilers. J. Anim. Sci. 2019, 97, 1679–1692. [Google Scholar] [CrossRef]
- Wang, M.-Y.; Zhang, Y.; Tong, Y.-X.; Guo, P.-T.; Zhang, J.; Wang, C.-K.; Gao, Y.-Y. Effects of lutein on jejunal mucosal barrier function and inflammatory responses in lipopolysaccharide-challenged yellow-feather broilers. Poult. Sci. 2022, 101, 102191. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhou, J.; Chen, M.; Huang, X.; Xie, X.; Li, W.; Cao, Q.; Kan, H.; Xu, Y.; Ying, Z. Exposure to concentrated ambient PM2.5 alters the composition of gut microbiota in a murine model. Part. Fibre Toxicol. 2018, 15, 17. [Google Scholar] [CrossRef] [PubMed]
- Keestra, A.M.; de Zoete, M.R.; Bouwman, L.I.; Vaezirad, M.M.; van Putten, J.P.M. Unique features of chicken Toll-like receptors. Dev. Comp. Immunol. 2013, 41, 316–323. [Google Scholar] [CrossRef]
- Li, W.; Guo, F.; Jiang, X.; Li, Y.; Li, X.; Yu, Z. Compound ammonium glycyrrhizin protects hepatocytes from injury induced by lipopolysaccharide/florfenicol through oxidative stress and a MAPK pathway. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2019, 225, 108585. [Google Scholar]
- Liu, S.D.; Song, M.H.; Yun, W.; Lee, J.H.; Kim, H.B.; Cho, J.H. Effect of carvacrol essential oils on immune response and inflammation-related genes expression in broilers challenged by lipopolysaccharide. Poult. Sci. 2019, 98, 2026–2033. [Google Scholar] [CrossRef]
- French, C.E.; Sales, M.A.; Rochell, S.J.; Rodriguez, A.; Erf, G.F. Local and systemic inflammatory responses to lipopolysaccharide in broilers: New insights using a two-window approach. Poult. Sci. 2022, 99, 6593–6605. [Google Scholar] [CrossRef] [PubMed]
- Arendt, M.; Elissa, J.; Schimdt, N.; Michael, E.; Potter, N.; Cook, M.; Knoll, L. Investigating the role of interleukin 10 on Eimeria intestinal pathogenesis in broiler chickens. Vet. Immunol. 2019, 218, 109934. [Google Scholar] [CrossRef]
- Kaiser, M.G.; Block, S.S.; Ciraci, C.; Fang, W.; Sifri, M.; Lamont, S.J. Effects of dietary vitamin E type and level on lipopolysaccharide-induced cytokine mRNA expression in broiler chicks. Poult. Sci. 2012, 91, 1893–1898. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhong, X.; Zhou, Y.; Wang, G.; Du, H.; Wang, T. Dietary RRR-a-tocopherol succinate attenuates lipopolysaccharide-induced inflammatory cytokines secretion in broiler chicks. Br. J. Nutr. 2010, 104, 1796–1805. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.C.; Wu, Q.J.; Song, Z.H.; Zhang, H.; Zhang, J.F.; Zhang, L.L.; Zhang, T.Y.; Wang, C.; Wang, T. Effects of Oridonin on growth performance and oxidative stress in broilers challenged with lipopolysaccharide. Poult. Sci. 2016, 95, 2281–2289. [Google Scholar] [CrossRef]
- Fukai, T.; Ushio-Fukai, M. Superoxide dismutases: Role in redox signaling, vascular function, and diseases. Antioxid. Redox Signal. 2011, 15, 1583–1606. [Google Scholar] [CrossRef] [PubMed]
- Satta, S.; Mahmoud, A.M.; Wilkinson, F.L.; Yvonne Alexander, M.; White, S.J. The role of Nrf2 in cardiovascular function and disease. Oxid. Med. Cell. Longev. 2017, 2017, 9237263. [Google Scholar] [CrossRef] [PubMed]
- Althunibat, O.Y.; Abduh, M.S.; Abukhalil, M.H.; Aladaileh, S.H.; Haineh, H.; Mahmoud, A.M. Umbelliferone prevents isoproterenol-induced myocardial injury by upregulating Nrf2/HO-1 signaling, and attenuating oxidative stress, inflammation, and cell death in rats. Biomed. Pharmacother. 2022, 149, 112900. [Google Scholar] [CrossRef]
- Wei, H.; Li, T.; Zhang, Y.; Liu, X.; Gong, R.; Bao, J.; Li, J. Cold stimulation causes oxidative stress, inflammatory response and apoptosis in broiler heart via regulating Nrf2/HO-1 and NF-κB pathway. J. Therm. Biol. 2023, 116, 103658. [Google Scholar] [CrossRef]

{kind=link}
| Ingredient | 8 to 20 Days, % |
|---|---|
| Corn, 7.88% | 50.38 |
| Soybean meal, 45.0% | 41.12 |
| Soybean oil | 4.58 |
| Dicalcium phosphate | 1.67 |
| Limestone | 0.84 |
| Salt | 0.52 |
| DL-Methionine, 99% | 0.32 |
| L-Lysine, 78% | 0.15 |
| L-Threonine, 98.5% | 0.05 |
| Choline chloride, 60% | 0.10 |
| Phytase | 0.01 |
| Antioxidant (BHT) 3 | 0.01 |
| Mineral Premix 1 | 0.13 |
| Vitamin Premix 2 | 0.13 |
| Calculated nutritional composition 4 | |
| Crude Protein, % | 22.3 |
| Metabolizable Energy, kcal/kg | 3000 |
| Calcium, % | 0.980 |
| Available Phosphorus, % | 0.480 |
| Sodium, % | 0.220 |
| Digestible Lysine, % | 1.240 |
| Digestible Methionine, % | 0.470 |
| Digestible Methionine + Cysteine, % | 0.929 |
| Digestible Threonine, % | 0.810 |
| Digestible Tryptophan, % | 0.267 |
| Gene | GenBank | Sequence |
|---|---|---|
| NF-κB | NM_205129.1 | F: 5′-GTGTGAAGAAACGGGAACTG-3′ |
| R: 5′-GGCACGCTTGTCATAGATGG-3′ | ||
| CAT | NM_001031215.2 | F: 5′-ACTGCAAGGCGAAAGTGTTT-3′ |
| R: 5′-GGCTATGGATGAAGGATGGA-3′ | ||
| GPX | NM_001277853.2 | F: 5′-GACCAACCCGCAGTACATCA-3′ |
| R: 5′-GAGGTGCGGGCTTTCCTTTA-3′ | ||
| SOD1 | NM_205064.1 | F: 5′-AGGGGGTCATCCACTTCC-3′ |
| R: 5′-CCCATTTGTGTTGTCTCCAA-3′ | ||
| IL-10 | NM_001004414.2 | F: 5′-CATGCTGCTGGGCCTGAA-3′ |
| R: 5′-CGTCTCCTTGATCTGCTTGATG-3′ | ||
| IL-1β | NM_204524.1 | F: 5′-GCTCTACATGTCGTGTGTGATGAG-3′ |
| R: 5′-TGTCGATGTCCCGCATGA-3′ | ||
| IL-8 | HM179639.1 | F: 5′-GGCTTGCTAGGGGAAATGA-3′ |
| R: 5′-AGCTGACTCTGACTAGGAAACTGT-3′ | ||
| β-ACT | NM_205518.1 | F: 5′-TGCTGTGTTCCCATCTATCG-3′ |
| R: 5′-TTGGTGACAATACCGTGTTCA-3′ |
| CON | CON + LPS | HT + LPS | p Value | SEM | |
|---|---|---|---|---|---|
| FI, kg/bird | 0.934 | 0.926 | 0.910 | 0.09 | 0.005 |
| BWG, kg/bird | 0.741 a | 0.704 b | 0.736 a | <0.001 | 0.005 |
| BW, kg/bird | 0.973 a | 0.932 b | 0.967 a | <0.001 | 0.005 |
| FCR, kg/kg | 1.262 a | 1.317 b | 1.238 a | <0.001 | 0.010 |
| CON | CON + LPS | HT + LPS | p Value | SEM | |
|---|---|---|---|---|---|
| Liver, % | 2.494 b | 3.038 a | 2.853 a | 0.001 | 0.067 |
| Spleen, % | 0.099 b | 0.167 a | 0.150 a | <0.0001 | 0.007 |
| Bursa, % | 0.214 | 0.213 | 0.213 | 0.997 | 0.009 |
| CON | CON + LPS | HT + LPS | p Value | SEM | |
|---|---|---|---|---|---|
| VH, µm | 2100.85 | 1972.55 | 2108.57 | 0.324 | 40.91 |
| CD, µm | 477.77 a | 582.77 b | 448.81 a | <0.001 | 15.28 |
| VH:CD | 4.453 a | 3.417 b | 4.879 a | 0.004 | 0.198 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dias, K.M.M.; Oliveira, C.H.; Calderano, A.A.; Rostagno, H.S.; Gomes, K.M.; O’Connor, K.E.; Davis, R.; Walsh, M.; Britton, J.; Altieri, E.A.; et al. Dietary Hydroxytyrosol Supplementation on Growth Performance, Gut Morphometry, and Oxidative and Inflammatory Status in LPS-Challenged Broilers. Animals 2024, 14, 871. https://doi.org/10.3390/ani14060871
Dias KMM, Oliveira CH, Calderano AA, Rostagno HS, Gomes KM, O’Connor KE, Davis R, Walsh M, Britton J, Altieri EA, et al. Dietary Hydroxytyrosol Supplementation on Growth Performance, Gut Morphometry, and Oxidative and Inflammatory Status in LPS-Challenged Broilers. Animals. 2024; 14(6):871. https://doi.org/10.3390/ani14060871
Chicago/Turabian StyleDias, Kelly M. M., Carlos H. Oliveira, Arele A. Calderano, Horacio S. Rostagno, Kaique M. Gomes, Kevin E. O’Connor, Reeta Davis, Meg Walsh, James Britton, Enrico A. Altieri, and et al. 2024. "Dietary Hydroxytyrosol Supplementation on Growth Performance, Gut Morphometry, and Oxidative and Inflammatory Status in LPS-Challenged Broilers" Animals 14, no. 6: 871. https://doi.org/10.3390/ani14060871
APA StyleDias, K. M. M., Oliveira, C. H., Calderano, A. A., Rostagno, H. S., Gomes, K. M., O’Connor, K. E., Davis, R., Walsh, M., Britton, J., Altieri, E. A., & Albino, L. F. T. (2024). Dietary Hydroxytyrosol Supplementation on Growth Performance, Gut Morphometry, and Oxidative and Inflammatory Status in LPS-Challenged Broilers. Animals, 14(6), 871. https://doi.org/10.3390/ani14060871