
Reproductive Seasonality, Estrous Cycle, Pregnancy, and the Recurrence of Postpartum Estrus Based on Long-Term Profiles of Fecal Sex Steroid Hormone Metabolites regarding Zoo-Housed Female Golden Takins (Budorcas taxicolor bedfordi)

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
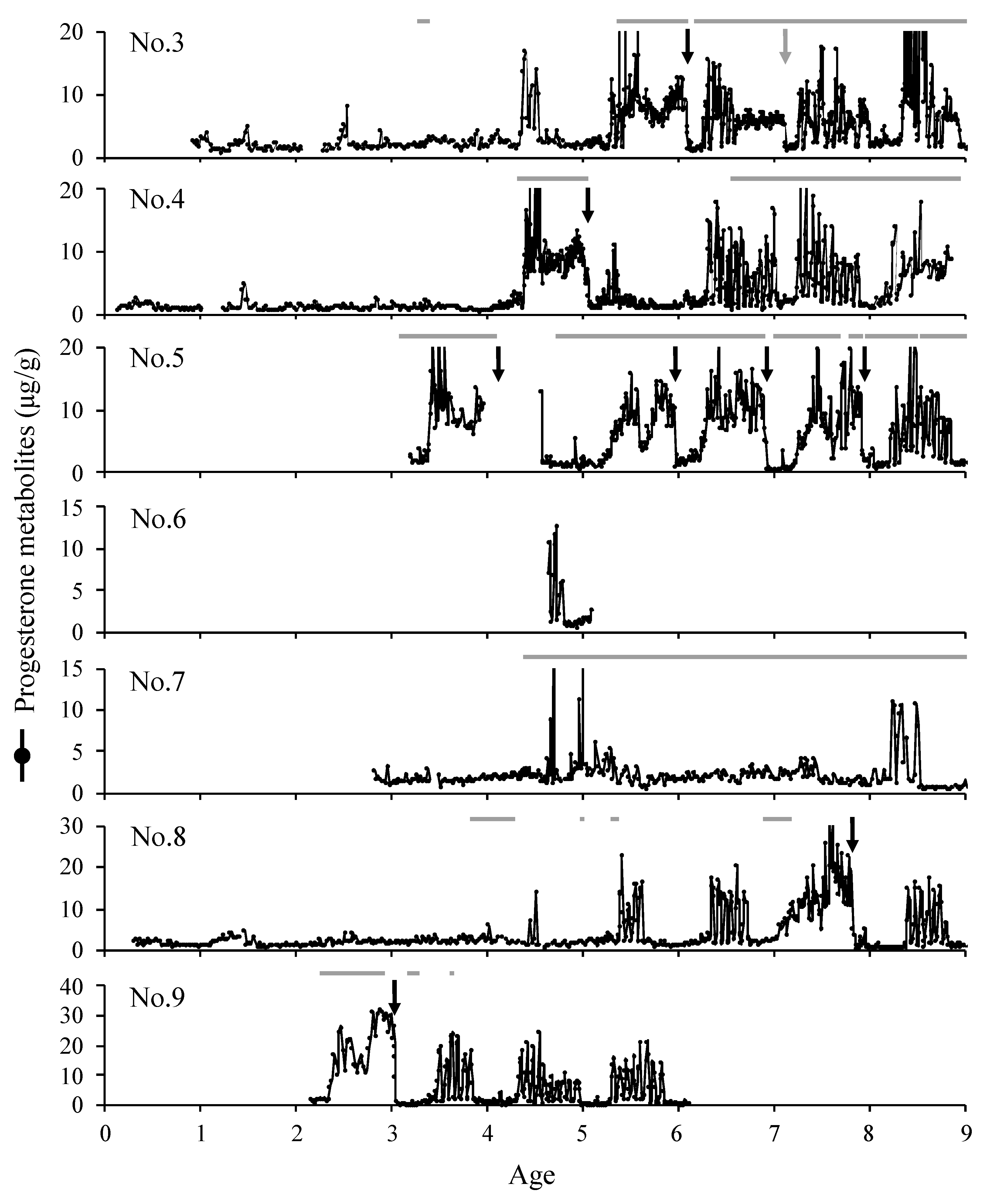
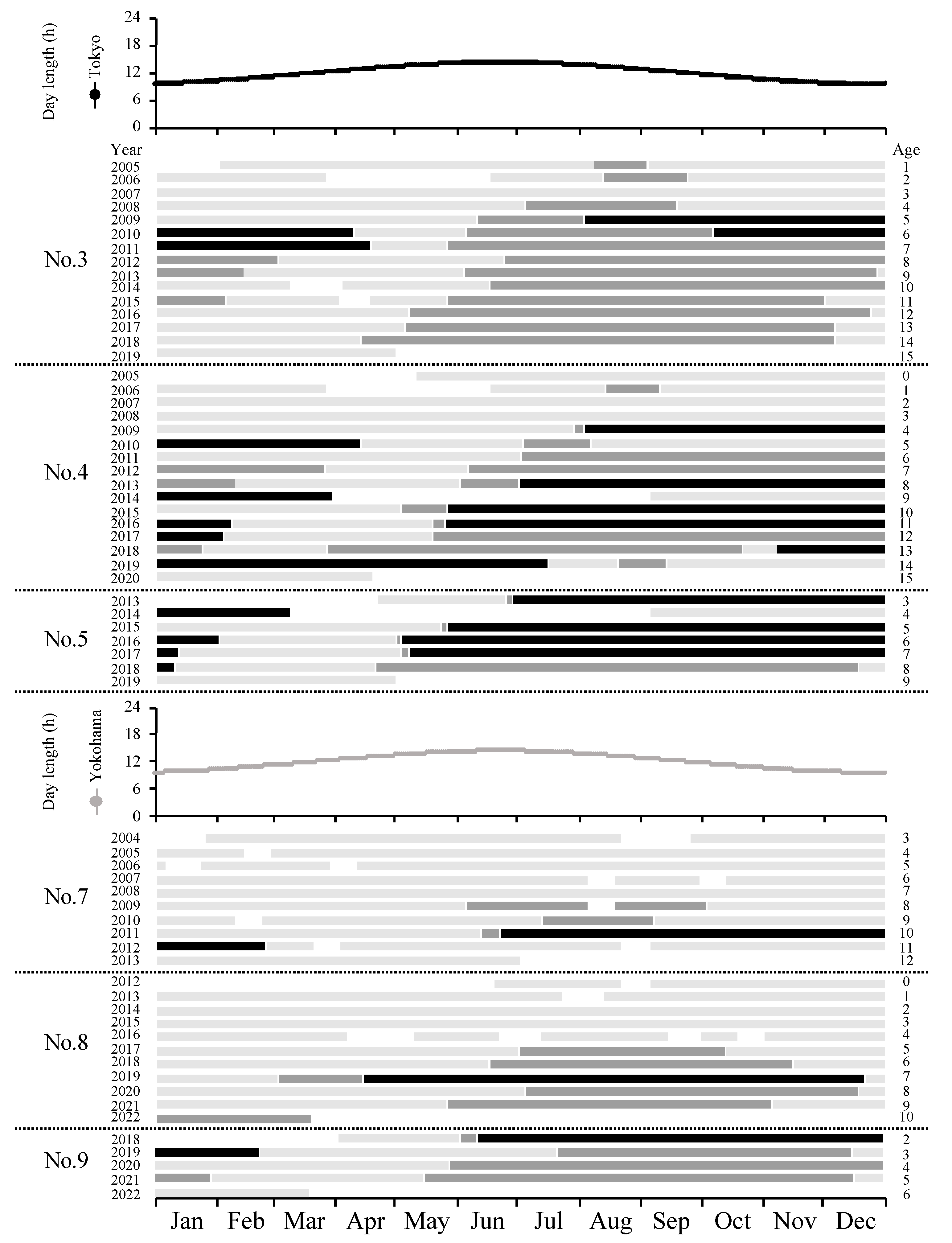
3. Results
3.1. Sexual Maturity
3.2. Breeding Season
3.3. The Estrous Cycle and Reproductive Behavior
3.4. Pregnancy and Recurrence of Postpartum Estrus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neas, J.F.; Hoffmann, R.S. Budorcas taxicolor. Mamm. Species 1987, 277, 1–7. [Google Scholar] [CrossRef]
- Song, Y.-L.; Smith, A.T.; MacKinnon, J. Budorcas taxicolor. The IUCN Red List of Threatened Species 2008: E.T3160A9643719. Available online: https://www.iucnredlist.org/species/3160/9643719 (accessed on 31 October 2023).
- CITES. Appendices I, II and III Valid 22 June 2022. Convention on International Trade in Endangered Species of Wild Fauna and Flora. Available online: https://cites.org/sites/default/files/eng/app/2022/E-Appendices-2022-06-22.pdf (accessed on 31 October 2023).
- Species 360. Zoological Information Management System. Available online: https://species360.org/ (accessed on 19 October 2023).
- EAZA. EAZA Ex Situ Programme (EEP). Available online: https://www.eaza.net/conservation/programmes/eep-pages/takin-eep/ (accessed on 31 October 2023).
- Schwarzenberger, F. The many uses of non-invasive faecal steroid monitoring in zoo and wildlife species. Int. Zoo Yb. 2007, 41, 52–74. [Google Scholar] [CrossRef]
- Zeng, Z.; Skidmore, A.K.; Song, Y.; Wang, T.; Gong, H. Seasonal altitudinal movements of Golden takin in the Qinling Mountains of China. J. Wildl. Manag. 2008, 72, 611–617. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, X.; Guo, S.; Li, B.; Meng, S. Reproductive biology in Golden takin. Chin. J. Zool. 2009, 44, 6–8. (In Chinese) [Google Scholar]
- Adkin, A.; Bernier, D.; Santymire, R.M. Characterizing the behavior and reproductive biology of zoo-housed Sichuan takin (Budorcas taxicolor tibetana) using non-invasive techniques. Theriogenology 2012, 78, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Santymire, R.M.; Adkin, A.; Bernier, D.; Hill, B. Validating the use of fecal glucocorticoid metabolite analysis to assess the adrenocortical activity of the zoo-housed Sichuan takin (Budorcas taxicolor tibetana). Zoo Biol. 2021, 40, 479–484. [Google Scholar] [CrossRef]
- National Astronomical Observatory of Japan. Local Calendar. Available online: https://eco.mtk.nao.ac.jp/koyomi/dni/ (accessed on 8 August 2023).
- Yoshida, T.; Kusuda, S.; Shimokawa, Y.; Orita, T. Reproductive seasonality and pregnancy based on fecal steroid steroid hormone profiles in zoo-housed chamois (Rupicapra rupicapra). Jpn. J. Zoo Wild Med. 2023, 28, 25–34, (In Japanese with English summary). [Google Scholar] [CrossRef]
- Kusuda, S.; Morikaku, K.; Kawada, K.; Ishiwada, K.; Doi, O. Excretion patterns of fecal progestagens, androgen, and estrogens during pregnancy, parturition and postpartum in okapi (Okapia johnstoni). J. Reprod. Dev. 2007, 53, 143–150. [Google Scholar] [CrossRef]
- Brown, J.L.; Lehnhardt, J. Serum and urinary hormones during pregnancy and the peri- and postpartum period in an Asian elephant (Elephas, maximus). Zoo Biol. 1995, 14, 555–564. [Google Scholar] [CrossRef]
- Patton, M.L.; Swaisgood, R.R.; Czekala, N.M.; White, A.M.; Fetter, G.A.; Montagne, J.P.; Rieches, R.G.; Lance, V.A. Reproductive cycle length and pregnancy in the southern white rhinoceros (Ceratotherium simum simum) as determined by fecal pregnane analysis and observations of mating behavior. Zoo Biol. 1999, 18, 111–127. [Google Scholar] [CrossRef]
- Schillo, K.K. (Furusawa, T. transration). Puberty chapter. In The Reproductive Physiology of Mammals: From Farm to Field and Beyond; Schillo, K.K., Sasada, H., Kosaka, T., Hashizume, K., Eds.; Kodansha: Tokyo, Japan, 2008; pp. 166–188. [Google Scholar]
- Boulanouar, B.; Ahmed, M.; Klopfenstein, T.; Brink, D.; Kinder, J. Dietary protein or energy restriction influences age and weight at puberty in ewe lambs. Anim. Reprod. Sci. 1995, 40, 229–238. [Google Scholar] [CrossRef]
- Togashi, M.; Tsujimoto, T.; Yamauchi, K.; Deguchi, Y.; Hashizume, K.; Kizaki, K.; Honjou, S.; Izaike, Y.; Osawa, T. Plasma and fecal sex steroid steroid hormone profile during the estrous cycle in a Japanese serow (Capricornis crispus). J. Reprod. Dev. 2009, 55, 412–417. [Google Scholar] [CrossRef][Green Version]
- Zeng, Z.-G.; Beck, S.A.P.; Wang, T.-J.; Skidmore, A.K.; Song, Y.-L.; Gong, H.-S.; Prins, H.H.T. Effects of plant phenology and solar radiation on seasonal movement of golden takin in the Qinling Mountains, China. J. Mammal. 2010, 91, 92–100. [Google Scholar] [CrossRef][Green Version]
- Kanomata, K.; Izawa, M. Some observations on estrus in female Japanese serows, Capricornis crispus. J. Jpn. Assoc. Zool. Gard. Aquar. 1982, 24, 61–63. [Google Scholar]
- Schwarzenberger, F.; Walzer, C.; Tomasova, K.; Vahala, J.; Meister, J.; Goodrowe, K.L.; Zima, J.; Strauß, G.; Lynch, M. Faecal progesterone metabolite analysis for non-invasive monitoring of reproductive function in the white rhinoceros (Ceratotherium simum). Anim. Reprod. Sci. 1998, 53, 173–190. [Google Scholar] [CrossRef] [PubMed]
- Rowell, J.E.; Flood, P.F. Progesterone, oestradiol-17β and LH during the oestrous cycle of muskoxen (Ovibos moschatus). J. Reprod. Fert. 1988, 84, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenberger, F.; Möstl, E.; Palme, R.; Bamberg, E. Faecal steroid analysis for non-invasive monitoring of reproductive status in farm, wild and zoo animals. Anim. Reprod. Sci. 1996, 42, 515–526. [Google Scholar] [CrossRef]
- Hindle, J.E.; Hodges, J.K. Metabolism of oestradiol-17β and progesterone in the white rhinoceros (Ceratotherium simum simum). J. Reprod. Fert. 1990, 90, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Palme, R.; Fischer, P.; Schildorfer, H.; Ismail, M.N. Excretion of infused 14C-steroid steroid hormones via faeces and urine in domestic livestock. Anim. Reprod. Sci. 1996, 43, 43–63. [Google Scholar] [CrossRef]
- Wasser, S.K.; Papageorge, S.; Foley, C.; Brown, J.L. Excretory fate of estradiol and progesterone in the African elephant (Loxodonta africana) and patterns of fecal steroid concentrations throughout the estrous cycle. Gen. Comp. Endocrinol. 1996, 102, 255–262. [Google Scholar] [CrossRef]
- Heistermann, M.; Agil, M.; Büthe, A.; Hodges, J.K. Metabolism and excretion of oestradiol-17β and progesterone in the Sumatran rhinoceros (Dicerorhinus sumatrensis). Anim. Reprod. Sci. 1998, 53, 157–172. [Google Scholar] [CrossRef]
- Abáigar, T.; Domené, M.A.; Cassinello, J. Characterization of the estrous cycle and reproductive traits of the aoudad (Ammotragus lervia) in captivity. Theriogenology 2012, 77, 1759–1766. [Google Scholar] [CrossRef]
- Khonmee, J.; Brown, J.L.; Taya, K.; Rojanasthien, S.; Punyapornwithaya, V.; Thumasanukul, D.; Kongphoemphun, A.; Siriaroonrat, B.; Tipkantha, W.; Pongpiachan, P.; et al. Assessment of ovarian activity in captive goral (Naemorhedus griseus) using noninvasive fecal steroid monitoring. Theriogenology 2014, 82, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- O’Hanlon, J.; Mogey, R.; Walker, S.L.; Roffe, S.; Clarke, H.; Edwards, K.L. Utilising routine non-invasive faecal samples for the detection of oestrus and early gestation in okapi (Okapi johnstoni). Theriogenol. Wild 2023, 3, 100047. [Google Scholar] [CrossRef]
- Sarda, I.R.; Robertson, H.A.; Smeaton, T.C. Sequential changes in plasma progesterone levels in the ewe during the estrous cycle, and during pregnancy in intact and ovariectomized sheep. Can. J. Anim. Sci. 1973, 53, 25–34. [Google Scholar] [CrossRef]
- Capezzuto, A.; Chelini, M.O.M.; Felippe, E.C.G.; Oliveira, C.A. Correlation between serum and fecal concentrations of reproductive steroids throughout gestation in goats. Anim. Reprod. Sci. 2008, 103, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Cowie, A.T.; Daniel, P.M.; Prichard, M.M.L. Hypophysectomy in pregnant goats, and section of the pituitary stalk in pregnant goats and sheep. J. Endocrin. 1963, 28, 93–102. [Google Scholar] [CrossRef]
- Al-Gubory, K.H.; Solari, A.; Mirman, B. Effects of luteectomy on the maintenance of pregnancy, circulating progesterone concentrations and lambing performance in sheep. Reprod. Fertil. Dev. 1999, 11, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Edgerton, L.A. Effect of lactation upon the postpartum interval. J. Anim. Sci. 1980, 51, 40–52. [Google Scholar] [PubMed]
- Lawson, J.L.; Forrest, D.W.; Shelton, M. Reproductive response to suckling manipulation in Spanish goats. Theriogenology 1984, 21, 747–755. [Google Scholar] [CrossRef]
- Short, R.E.; Bellows, R.A.; Moody, E.L.; Howland, B.E. Effects of suckling and mastectomy on bovine postpartum reproduction. J. Anim. Sci. 1972, 34, 70–74. [Google Scholar] [CrossRef] [PubMed]
- LaVoie, V.; Han, D.K.; Foster, D.B.; Moody, E.L. Suckling effect on estrus and blood plasma progesterone in postpartum beef cows. J. Anim. Sic. 1981, 52, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Pickard, A.R.; Abáigar, T.; Green, D.I.; Holt, W.V. Hormonal characterization of the reproductive cycle and pregnancy in the female Mohor gazelle (Gazella dama mhorr). Reproduction 2001, 122, 571–580. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Individual No. (Name) | Birth Date | Facility | Sample Collection Period | Age of Analysis Period | Death Date |
|---|---|---|---|---|---|
| No. 1 (Houla) | March 1990 * | TZP ** | 7 January 2004~26 January 2005 | 13.9~14.9 | 31 January 2005 |
| No. 2 (Ohi) | 31 March 1991 * | 7 January 2004~28 February 2013 | 12.8~21.9 | 11 November 2013 | |
| No. 3 (Fuuka) | 4 March 2004 | 2 February 2005~30 April 2019 | 0.9~15.2 | − | |
| No. 4 (Ouki) | 20 March 2005 | 11 May 2005~19 April 2020 | 0.1~15.1 | 23 December 2022 | |
| No. 5 (Hoi) | 10 February 2010 | 22 April 2013~30 April 2019 | 3.2~9.2 | − | |
| No. 6 (Fuku) | 3 February 2017 | 28 September 2021~13 March 2022 | 4.7~5.1 | − | |
| No. 7 (Turco) | 27 March 2001 | YZG ** | 25 January 2004~2 July 2013 | 2.8~12.3 | 18 December 2014 |
| No. 8 (Hinata) | 24 February 2012 | 19 June 2012~19 March 2022 | 0.3~10.1 | − | |
| No. 9 (Mei) | 7 February 2016 | 3 April 2018~19 March 2022 | 2.2~6.1 | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshida, T.; Shimokawa, Y.; Ohta, M.; Takayanagi, M.; Kusuda, S. Reproductive Seasonality, Estrous Cycle, Pregnancy, and the Recurrence of Postpartum Estrus Based on Long-Term Profiles of Fecal Sex Steroid Hormone Metabolites regarding Zoo-Housed Female Golden Takins (Budorcas taxicolor bedfordi). Animals 2024, 14, 571. https://doi.org/10.3390/ani14040571
Yoshida T, Shimokawa Y, Ohta M, Takayanagi M, Kusuda S. Reproductive Seasonality, Estrous Cycle, Pregnancy, and the Recurrence of Postpartum Estrus Based on Long-Term Profiles of Fecal Sex Steroid Hormone Metabolites regarding Zoo-Housed Female Golden Takins (Budorcas taxicolor bedfordi). Animals. 2024; 14(4):571. https://doi.org/10.3390/ani14040571
Chicago/Turabian StyleYoshida, Tomoki, Yuki Shimokawa, Makoto Ohta, Mayo Takayanagi, and Satoshi Kusuda. 2024. "Reproductive Seasonality, Estrous Cycle, Pregnancy, and the Recurrence of Postpartum Estrus Based on Long-Term Profiles of Fecal Sex Steroid Hormone Metabolites regarding Zoo-Housed Female Golden Takins (Budorcas taxicolor bedfordi)" Animals 14, no. 4: 571. https://doi.org/10.3390/ani14040571
APA StyleYoshida, T., Shimokawa, Y., Ohta, M., Takayanagi, M., & Kusuda, S. (2024). Reproductive Seasonality, Estrous Cycle, Pregnancy, and the Recurrence of Postpartum Estrus Based on Long-Term Profiles of Fecal Sex Steroid Hormone Metabolites regarding Zoo-Housed Female Golden Takins (Budorcas taxicolor bedfordi). Animals, 14(4), 571. https://doi.org/10.3390/ani14040571