

Olive Mill Waste-Water Extract Enriched in Hydroxytyrosol and Tyrosol Modulates Host–Pathogen Interaction in IPEC-J2 Cells
 , , ,
, , ,  ,
,  , , and
, , and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Olive Mill Waste-Water Extract and Composition
2.2. Cell Cultures
2.2.1. Cell Viability
2.2.2. Bacterial Invasion
2.2.3. Modulation of the Immune Response
2.3. RNA Extraction and Reverse Transcription Quantitative PCR (RT-qPCR)
2.4. Cytokine Quantification
2.5. Statistical Analyses
3. Results
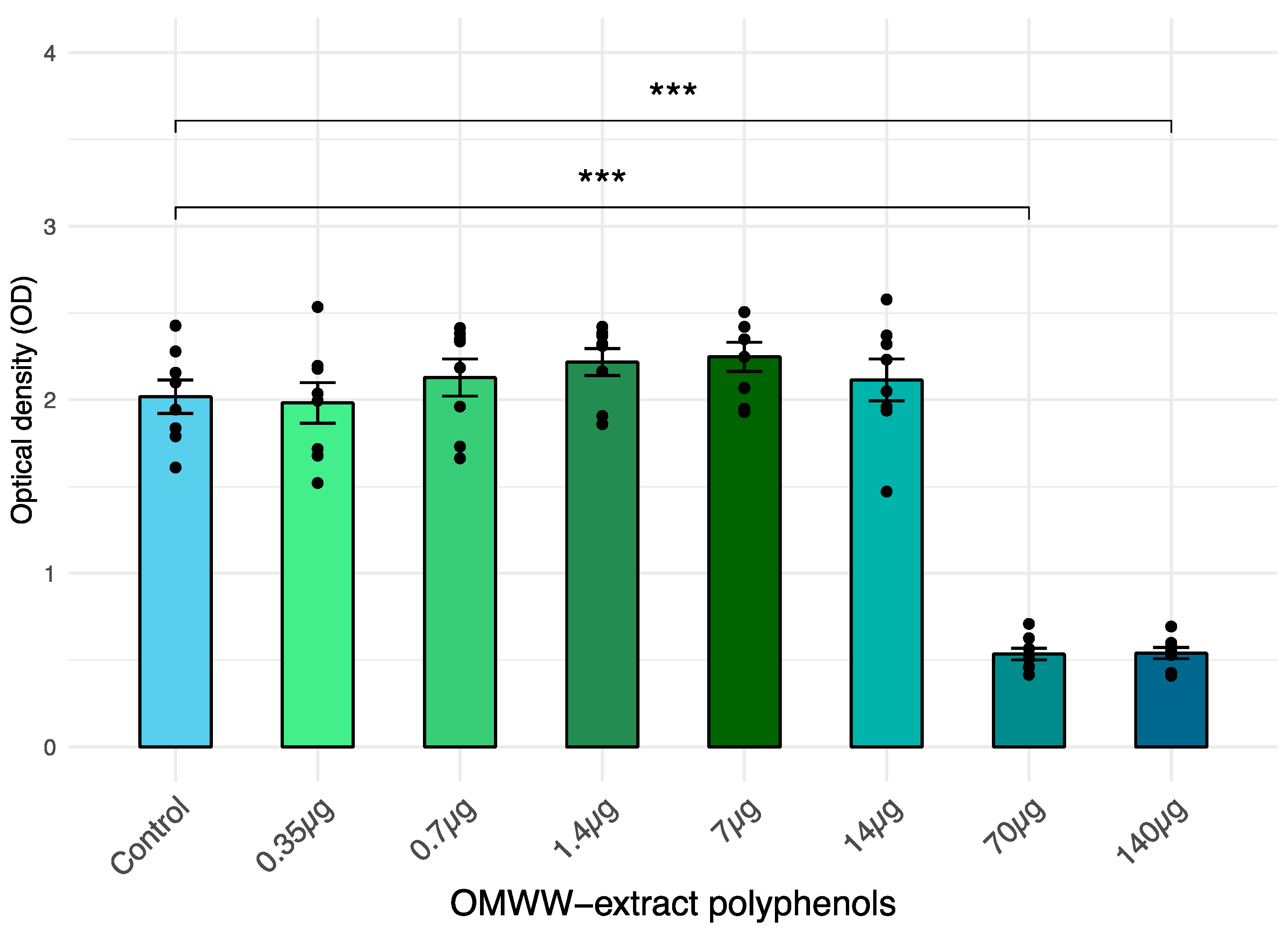
3.1. Cell Viability
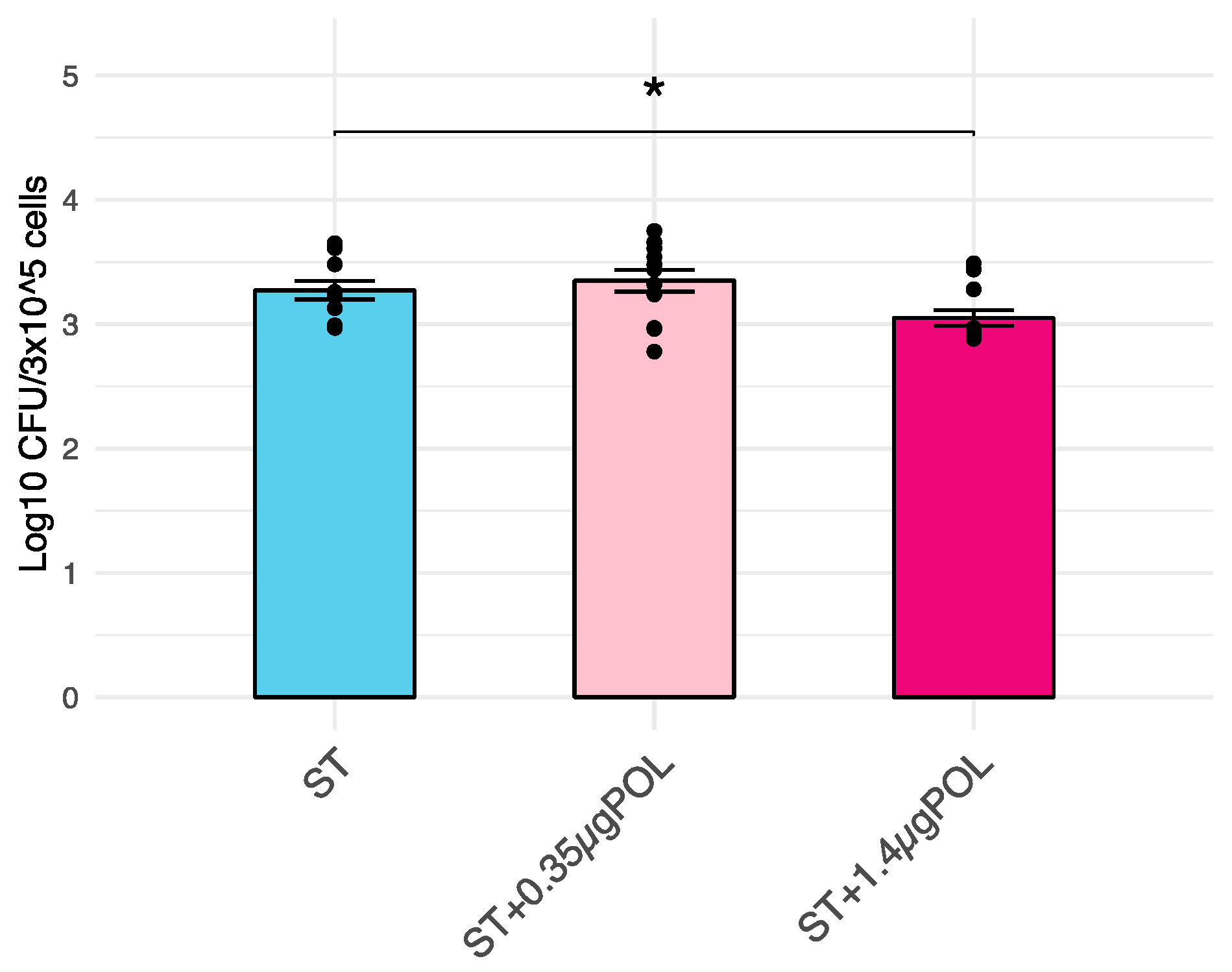
3.2. Salmonella Typhimurium Invasiveness
3.3. Modulation of Immune Response
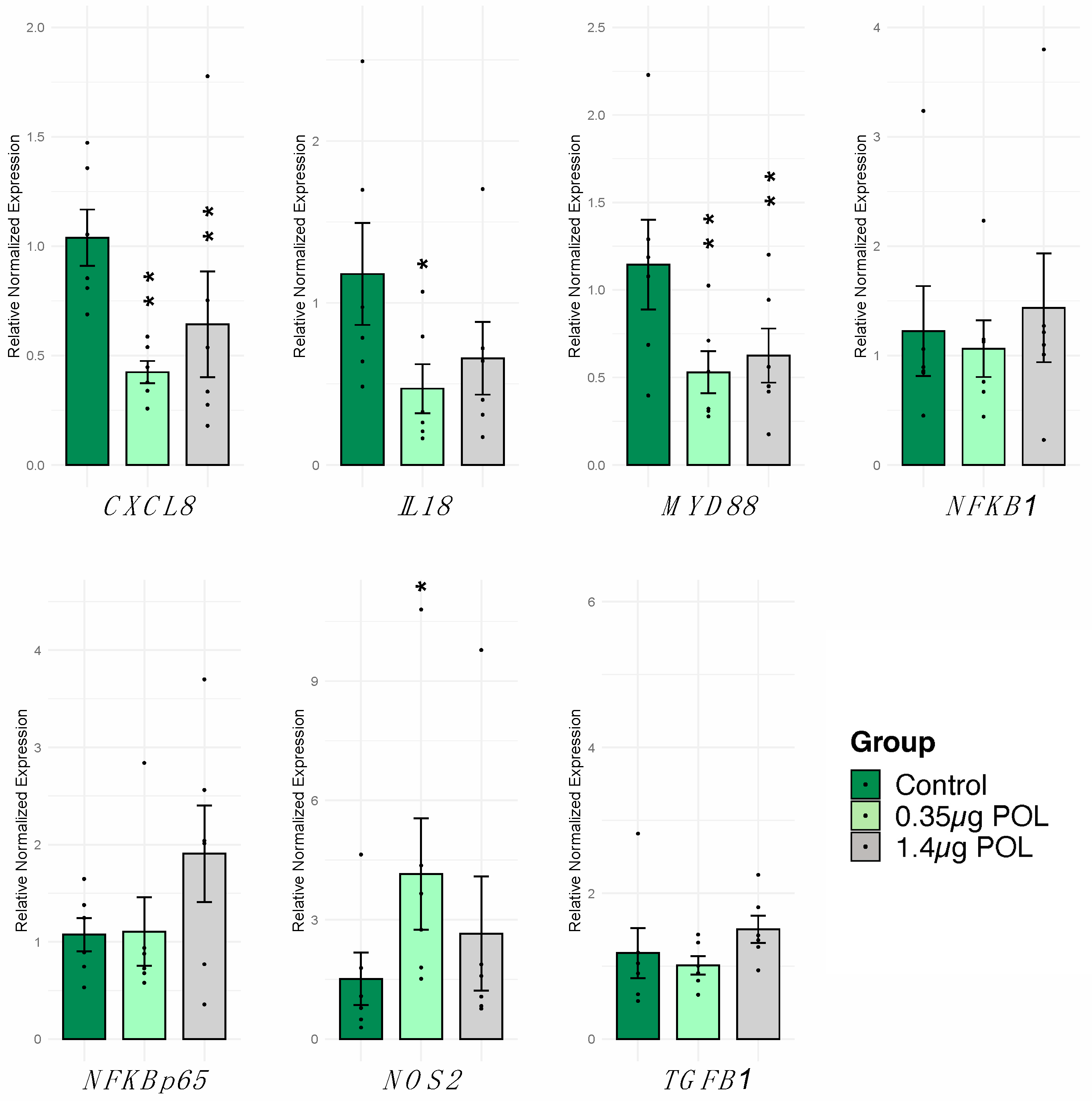
3.3.1. OMWW-Extract Polyphenols’ Effect on IPEC-J2 Gene Expression and Cytokine Release
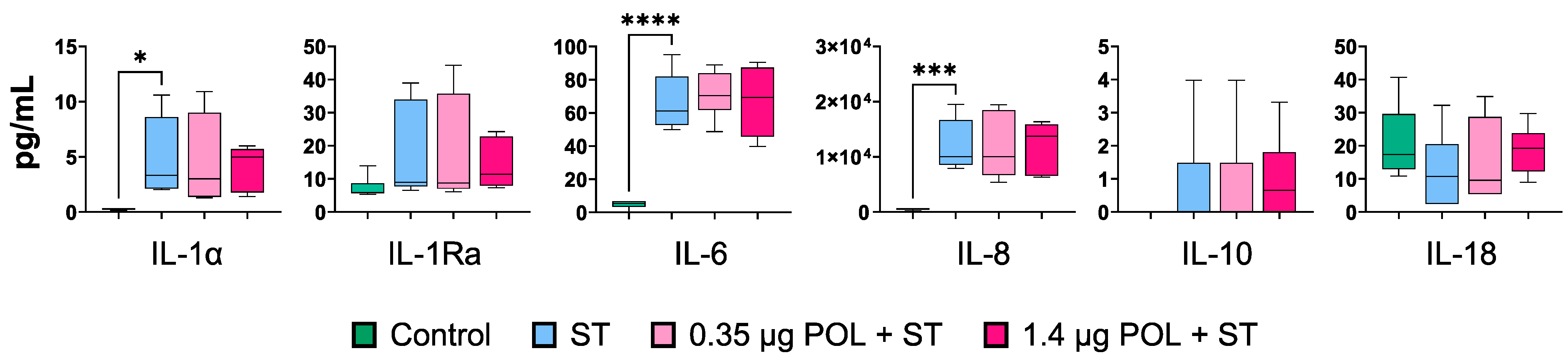
3.3.2. OMWW-Extract Polyphenols’ Effect on IPEC-J2 Cytokine Release
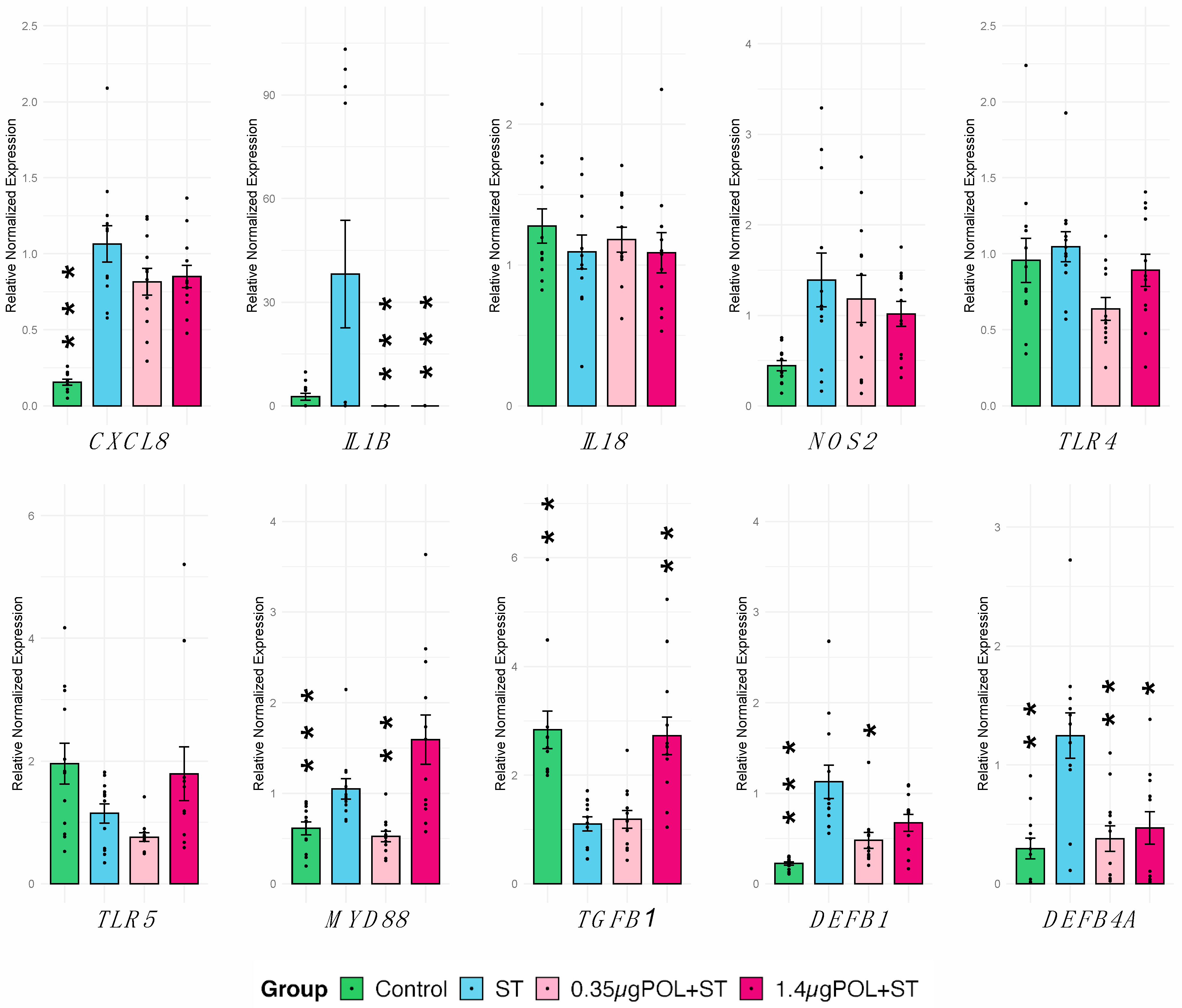
3.3.3. OMWW-Extract Polyphenols’ and S. typhimurium Infection Effects on IPEC-J2 Gene Expression
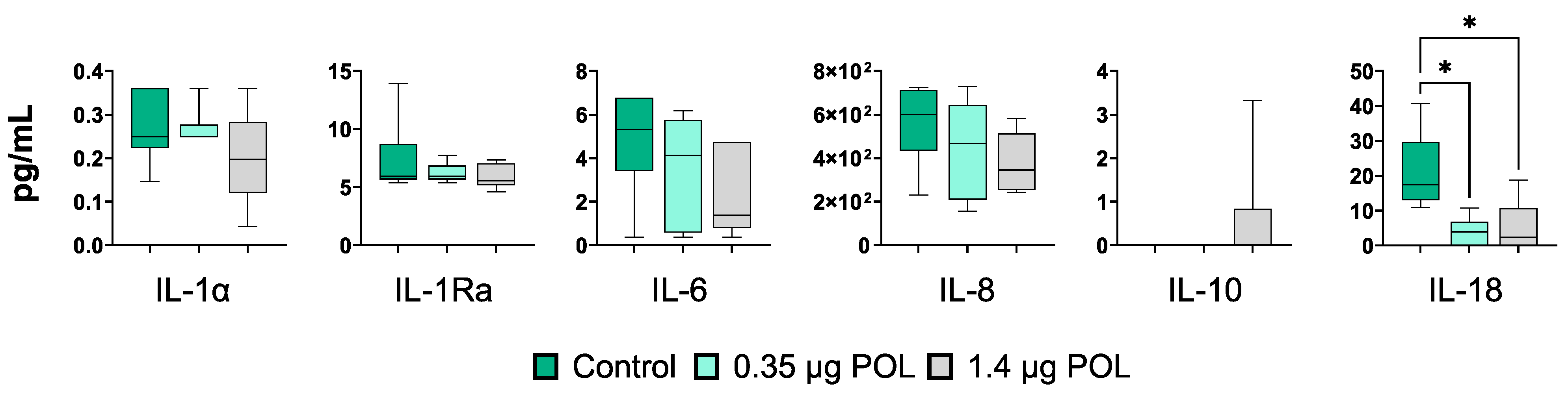
3.3.4. OMWW-Extract Polyphenols’ and S. typhimurium Infection Effects on IPEC-J2 Cytokine Production
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Capasso, R.; Cristinzio, G.; Evidente, A.; Scognamiglio, F. Isolation, Spectroscopy and Selective Phytotoxic Effects of Polyphenols from Vegetable Waste Waters. Phytochemistry 1992, 31, 4125–4128. [Google Scholar] [CrossRef]
- Mekki, A.; Dhouib, A.; Sayadi, S. Changes in Microbial and Soil Properties Following Amendment with Treated and Untreated Olive Mill Wastewater. Microbiol. Res. 2006, 161, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Ben Sassi, A.; Boularbah, A.; Jaouad, A.; Walker, G.; Boussaid, A. A Comparison of Olive Oil Mill Wastewaters (OMW) from Three Different Processes in Morocco. Process Biochem. 2006, 41, 74–78. [Google Scholar] [CrossRef]
- Casa, R.; D’Annibale, A.; Pieruccetti, F.; Stazi, S.R.; Giovannozzi Sermanni, G.; Lo Cascio, B. Reduction of the Phenolic Components in Olive-Mill Wastewater by an Enzymatic Treatment and Its Impact on Durum Wheat (Triticum durum Desf.) Germinability. Chemosphere 2003, 50, 959–966. [Google Scholar] [CrossRef] [PubMed]
- El Hadrami, A.; Belaqziz, M.; El Hassni, M.; Hanifi, S.; Abbad, A.; Capasso, R.; Gianfreda, L.; El Hadrami, I. Physico-chemical characterization and effects of olive oil mill wastewaters fertirrigation on the growth of some Mediterranean crops. J. Agron. 2004, 3, 247–254. [Google Scholar] [CrossRef]
- Quaratino, D.; D’Annibale, A.; Federici, F.; Cereti, C.F.; Rossini, F.; Fenice, M. Enzyme and Fungal Treatments and a Combination Thereof Reduce Olive Mill Wastewater Phytotoxicity on Zea mays L. Seeds. Chemosphere 2007, 66, 1627–1633. [Google Scholar] [CrossRef] [PubMed]
- Rodis, P.S.; Karathanos, V.T.; Mantzavinou, A. Partitioning of Olive Oil Antioxidants between Oil and Water Phases. J. Agric. Food Chem. 2002, 50, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Bertin, L.; Ferri, F.; Scoma, A.; Marchetti, L.; Fava, F. Recovery of High Added Value Natural Polyphenols from Actual Olive Mill Wastewater through Solid Phase Extraction. Chem. Eng. J. 2011, 171, 1287. [Google Scholar] [CrossRef]
- De Marco, E.; Savarese, M.; Paduano, A.; Sacchi, R. Characterization and Fractionation of Phenolic Compounds Extracted from Olive Oil Mill Wastewaters. Food Chem. 2007, 104, 858–867. [Google Scholar] [CrossRef]
- Varricchio, E.; Coccia, E.; Orso, G.; Lombardi, V.; Imperatore, R.; Vito, P.; Paolucci, M. Influence of Polyphenols from Olive Mill Wastewater on the Gastrointestinal Tract, Alveolar Macrophages and Blood Leukocytes of Pigs. Ital. J. Anim. Sci. 2019, 18, 574–586. [Google Scholar] [CrossRef]
- Branciari, R.; Ranucci, D.; Ortenzi, R.; Roila, R.; Trabalza-Marinucci, M.; Servili, M.; Papa, P.; Galarini, R.; Valiani, A. Dietary Administration of Olive Mill Wastewater Extract Reduces Campylobacter Spp. Prevalence in Broiler Chickens. Sustainability 2016, 8, 837. [Google Scholar] [CrossRef]
- Hume, D.A.; Whitelaw, C.B.A.; Archibald, A.L. The Future of Animal Production: Improving Productivity and Sustainability. J. Agric. Sci. 2011, 149, 9–16. [Google Scholar] [CrossRef]
- Obied, H.K.; Bedgood, D.R.; Prenzler, P.D.; Robards, K. Bioscreening of Australian Olive Mill Waste Extracts: Biophenol Content, Antioxidant, Antimicrobial and Molluscicidal Activities. Food Chem. Toxicol. 2007, 45, 1238–1248. [Google Scholar] [CrossRef] [PubMed]
- Servili, M.; Sordini, B.; Esposto, S.; Urbani, S.; Veneziani, G.; Di Maio, I.; Selvaggini, R.; Taticchi, A. Biological Activities of Phenolic Compounds of Extra Virgin Olive Oil. Antioxidants 2013, 3, 1–23. [Google Scholar] [CrossRef]
- Veneziani, G.; Novelli, E.; Esposto, S.; Taticchi, A.; Servili, M. Applications of Recovered Bioactive Compounds in Food Products. In Olive Mill Waste: Recent Advances for Sustainable Management; Academic Press: Cambridge, MA, USA, 2017; pp. 231–253. ISBN 978-0-12-805314-0. [Google Scholar]
- Salomone, R.; Ioppolo, G. Environmental Impacts of Olive Oil Production: A Life Cycle Assessment Case Study in the Province of Messina (Sicily). J. Clean. Prod. 2012, 28, 88–100. [Google Scholar] [CrossRef]
- Vastolo, A.; Calabrò, S.; Cutrignelli, M.I. A Review on the Use of Agro-Industrial CO-Products in Animals’ Diets. Ital. J. Anim. Sci. 2022, 21, 577–594. [Google Scholar] [CrossRef]
- Branciari, R.; Galarini, R.; Giusepponi, D.; Trabalza-Marinucci, M.; Forte, C.; Roila, R.; Miraglia, D.; Servili, M.; Acuti, G.; Valiani, A. Oxidative Status and Presence of Bioactive Compounds in Meat from Chickens Fed Polyphenols Extracted from Olive Oil Industry Waste. Sustainability 2017, 9, 1566. [Google Scholar] [CrossRef]
- Sabino, M.; Cappelli, K.; Capomaccio, S.; Pascucci, L.; Biasato, I.; Verini-Supplizi, A.; Valiani, A.; Trabalza-Marinucci, M. Dietary Supplementation with Olive Mill Wastewaters Induces Modifications on Chicken Jejunum Epithelial Cell Transcriptome and Modulates Jejunum Morphology. BMC Genom. 2018, 19, 576. [Google Scholar] [CrossRef]
- Ferlisi, F.; Tang, J.; Cappelli, K.; Trabalza-Marinucci, M. Dietary Supplementation with Olive Oil Co-Products Rich in Polyphenols: A Novel Nutraceutical Approach in Monogastric Animal Nutrition. Front. Vet. Sci. 2023, 10, 1272274. [Google Scholar] [CrossRef]
- Maranesi, M.; Dall’Aglio, C.; Acuti, G.; Cappelli, K.; Trabalza Marinucci, M.; Galarini, R.; Suvieri, C.; Zerani, M. Effects of Dietary Polyphenols from Olive Mill Waste Waters on Inflammatory and Apoptotic Effectors in Rabbit Ovary. Animals 2021, 11, 1727. [Google Scholar] [CrossRef]
- Cappelli, K.; Ferlisi, F.; Mecocci, S.; Maranesi, M.; Trabalza-Marinucci, M.; Zerani, M.; Dal Bosco, A.; Acuti, G. Dietary Supplementation of Olive Mill Waste Water Polyphenols in Rabbits: Evaluation of the Potential Effects on Hepatic Apoptosis, Inflammation and Metabolism through RT-qPCR Approach. Animals 2021, 11, 2932. [Google Scholar] [CrossRef] [PubMed]
- Lessard, M.; Talbot, G.; Bergeron, N.; Verso, L.L.; Morissette, B.; Yergeau, É.; Matte, J.J.; Bissonnette, N.; Blais, M.; Gong, J.; et al. Weaning Diet Supplemented with Health-Promoting Feed Additives Influences Microbiota and Immune Response in Piglets Challenged with Salmonella. Vet. Immunol. Immunopathol. 2023, 255, 110533. [Google Scholar] [CrossRef] [PubMed]
- Lian, S.; Lin, X.; Zhan, F.; Shen, X.; Liang, Y.; Li, C. Transcriptome Analysis Reveals the Multiple Functions of pBD2 in IPEC-J2 Cells against E. coli. Int. J. Mol. Sci. 2022, 23, 9754. [Google Scholar] [CrossRef]
- Van den Bogaard, A.E.; Stobberingh, E.E. Antibiotic Usage in Animals. Drugs 1999, 58, 589–607. [Google Scholar] [CrossRef] [PubMed]
- Veldhuizen, E.J.A.; Koomen, I.; Ultee, T.; van Dijk, A.; Haagsman, H.P. Salmonella Serovar Specific Upregulation of Porcine Defensins 1 and 2 in a Jejunal Epithelial Cell Line. Vet. Microbiol. 2009, 136, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Farzan, A.; Friendship, R.M. A Clinical Field Trial to Evaluate the Efficacy of Vaccination in Controlling Salmonella Infection and the Association of Salmonella-Shedding and Weight Gain in Pigs. Can. J. Vet. Res. 2010, 74, 258–263. [Google Scholar] [PubMed]
- Naberhaus, S.A.; Krull, A.C.; Arruda, B.L.; Arruda, P.; Sahin, O.; Schwartz, K.J.; Burrough, E.R.; Magstadt, D.R.; Matias Ferreyra, F.; Gatto, I.R.H.; et al. Pathogenicity and Competitive Fitness of Salmonella enterica Serovar 4,[5],12:i:- Compared to Salmonella Typhimurium and Salmonella Derby in Swine. Front. Vet. Sci. 2019, 6, 502. [Google Scholar] [CrossRef]
- Scott, M.B.; Styring, A.K.; McCullagh, J.S.O. Polyphenols: Bioavailability, Microbiome Interactions and Cellular Effects on Health in Humans and Animals. Pathogens 2022, 11, 770. [Google Scholar] [CrossRef]
- Romier, B.; Schneider, Y.-J.; Larondelle, Y.; During, A. Dietary Polyphenols Can Modulate the Intestinal Inflammatory Response. Nutr. Rev. 2009, 67, 363–378. [Google Scholar] [CrossRef]
- Zhang, Y.; Mu, T.; Deng, X.; Guo, R.; Xia, B.; Jiang, L.; Wu, Z.; Liu, M. New Insights of Biological Functions of Natural Polyphenols in Inflammatory Intestinal Diseases. Int. J. Mol. Sci. 2023, 24, 9581. [Google Scholar] [CrossRef]
- Axling, U.; Olsson, C.; Xu, J.; Fernandez, C.; Larsson, S.; Ström, K.; Ahrné, S.; Holm, C.; Molin, G.; Berger, K. Green Tea Powder and Lactobacillus plantarum Affect Gut Microbiota, Lipid Metabolism and Inflammation in High-Fat Fed C57BL/6J Mice. Nutr. Metab. 2012, 9, 105. [Google Scholar] [CrossRef] [PubMed]
- Tassou, C.C.; Nychas, G.J.; Board, R.G. Effect of Phenolic Compounds and Oleuropein on the Germination of Bacillus cereus T Spores. Biotechnol. Appl. Biochem. 1991, 13, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Anhê, F.F.; Pilon, G.; Roy, D.; Desjardins, Y.; Levy, E.; Marette, A. Triggering Akkermansia with Dietary Polyphenols: A New Weapon to Combat the Metabolic Syndrome? Gut Microbes 2016, 7, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Dueñas, M.; Muñoz-González, I.; Cueva, C.; Jiménez-Girón, A.; Sánchez-Patán, F.; Santos-Buelga, C.; Moreno-Arribas, M.V.; Bartolomé, B. A Survey of Modulation of Gut Microbiota by Dietary Polyphenols. BioMed Res. Int. 2015, 2015, e850902. [Google Scholar] [CrossRef] [PubMed]
- Hara, H.; Orita, N.; Hatano, S.; Ichikawa, H.; Hara, Y.; Matsumoto, N.; Kimura, Y.; Terada, A.; Mitsuoka, T. Effect of Tea Polyphenols on Fecal Flora and Fecal Metabolic Products of Pigs. J. Vet. Med. Sci. 1995, 57, 45–49. [Google Scholar] [CrossRef]
- Ishihara, N.; Chu, D.-C.; Akachi, S.; Juneja, L.R. Improvement of Intestinal Microflora Balance and Prevention of Digestive and Respiratory Organ Diseases in Calves by Green Tea Extracts. Livest. Prod. Sci. 2001, 68, 217–229. [Google Scholar] [CrossRef]
- Liu, Y. Assessment of the Antimicrobial Activity of Olive Leaf Extract Against Foodborne Bacterial Pathogens. Front. Microbiol. 2017, 8, 113. [Google Scholar] [CrossRef]
- Sudjana, A.N.; D’Orazio, C.; Ryan, V.; Rasool, N.; Ng, J.; Islam, N.; Riley, T.V.; Hammer, K.A. Antimicrobial Activity of Commercial Olea europaea (Olive) Leaf Extract. Int. J. Antimicrob. Agents 2009, 33, 461–463. [Google Scholar] [CrossRef]
- Chen, J.; Yu, B.; Chen, D.; Huang, Z.; Mao, X.; Zheng, P.; Yu, J.; Luo, J.; He, J. Chlorogenic Acid Improves Intestinal Barrier Functions by Suppressing Mucosa Inflammation and Improving Antioxidant Capacity in Weaned Pigs. J. Nutr. Biochem. 2018, 59, 84–92. [Google Scholar] [CrossRef]
- Chen, X.; Zeng, Z.; Huang, Z.; Chen, D.; He, J.; Chen, H.; Yu, B.; Yu, J.; Luo, J.; Luo, Y.; et al. Effects of Dietary Resveratrol Supplementation on Immunity, Antioxidative Capacity and Intestinal Barrier Function in Weaning Piglets. Anim. Biotechnol. 2021, 32, 240–245. [Google Scholar] [CrossRef]
- Ahasan, A.S.M.L.; Invernizzi, G.; Farina, G.; Pilotto, A.; Barbé, F.; Bontempo, V.; Rossi, R.; Bellagamba, F.; Lecchi, C.; Savoini, G.; et al. The Effects of Superoxide Dismutase-Rich Melon Pulp Concentrate on Inflammation, Antioxidant Status and Growth Performance of Challenged Post-Weaning Piglets. Animal 2019, 13, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Jiang, X.R.; Mantovani, G.; Lumbreras, A.E.V.; Comi, M.; Alborali, G.; Savoini, G.; Dell’Orto, V.; Bontempo, V. Modulation of Plasma Antioxidant Activity in Weaned Piglets by Plant Polyphenols; Taylor & Francis: Abingdon, UK, 2014. [Google Scholar]
- Fiesel, A.; Gessner, D.K.; Most, E.; Eder, K. Effects of Dietary Polyphenol-Rich Plant Products from Grape or Hop on Pro-Inflammatory Gene Expression in the Intestine, Nutrient Digestibility and Faecal Microbiota of Weaned Pigs. BMC Vet. Res. 2014, 10, 196. [Google Scholar] [CrossRef] [PubMed]
- Gessner, D.K.; Ringseis, R.; Eder, K. Potential of Plant Polyphenols to Combat Oxidative Stress and Inflammatory Processes in Farm Animals. J. Anim. Physiol. Anim. Nutr. 2017, 101, 605–628. [Google Scholar] [CrossRef] [PubMed]
- Mahfuz, S.; Shang, Q.; Piao, X. Phenolic Compounds as Natural Feed Additives in Poultry and Swine Diets: A Review. J. Anim. Sci. Biotechnol. 2021, 12, 48. [Google Scholar] [CrossRef] [PubMed]
- Mosele, J.I.; Macià, A.; Motilva, M.-J. Metabolic and Microbial Modulation of the Large Intestine Ecosystem by Non-Absorbed Diet Phenolic Compounds: A Review. Molecules 2015, 20, 17429–17468. [Google Scholar] [CrossRef] [PubMed]
- Mecocci, S.; De Paolis, L.; Fruscione, F.; Pietrucci, D.; De Ciucis, C.G.; Giudici, S.D.; Franzoni, G.; Chillemi, G.; Cappelli, K.; Razzuoli, E. In Vitro Evaluation of Immunomodulatory Activities of Goat Milk Extracellular Vesicles (mEVs) in a Model of Gut Inflammation. Res. Vet. Sci. 2022, 152, 546–556. [Google Scholar] [CrossRef]
- Razzuoli, E.; Amadori, M.; Lazzara, F.; Bilato, D.; Ferraris, M.; Vito, G.; Ferrari, A. Salmonella Serovar-Specific Interaction with Jejunal Epithelial Cells. Vet. Microbiol. 2017, 207, 219–225. [Google Scholar] [CrossRef]
- Razzuoli, E.; Villa, R.; Amadori, M. IPEC-J2 Cells as Reporter System of the Anti-Inflammatory Control Actions of Interferon-Alpha. J. Interferon Cytokine Res. 2013, 33, 597–605. [Google Scholar] [CrossRef]
- Sonntag, A.-K.; Bielaszewska, M.; Mellmann, A.; Dierksen, N.; Schierack, P.; Wieler, L.H.; Schmidt, M.A.; Karch, H. Shiga Toxin 2e-Producing Escherichia coli Isolates from Humans and Pigs Differ in Their Virulence Profiles and Interactions with Intestinal Epithelial Cells. Appl. Environ. Microbiol. 2005, 71, 8855–8863. [Google Scholar] [CrossRef]
- De Ciucis, C.G.; Fruscione, F.; De Paolis, L.; Mecocci, S.; Zinellu, S.; Guardone, L.; Franzoni, G.; Cappelli, K.; Razzuoli, E. Toll-like Receptors and Cytokine Modulation by Goat Milk Extracellular Vesicles in a Model of Intestinal Inflammation. Int. J. Mol. Sci. 2023, 24, 11096. [Google Scholar] [CrossRef]
- Mariani, V.; Palermo, S.; Fiorentini, S.; Lanubile, A.; Giuffra, E. Gene Expression Study of Two Widely Used Pig Intestinal Epithelial Cell Lines: IPEC-J2 and IPI-2I. Vet. Immunol. Immunopathol. 2009, 131, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Schierack, P.; Nordhoff, M.; Pollmann, M.; Weyrauch, K.D.; Amasheh, S.; Lodemann, U.; Jores, J.; Tachu, B.; Kleta, S.; Blikslager, A.; et al. Characterization of a Porcine Intestinal Epithelial Cell Line for in Vitro Studies of Microbial Pathogenesis in Swine. Histochem. Cell Biol. 2006, 125, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.A.; Artis, D. Intestinal Bacteria and the Regulation of Immune Cell Homeostasis. Annu. Rev. Immunol. 2010, 28, 623–667. [Google Scholar] [CrossRef] [PubMed]
- Dejban, P.; Nikravangolsefid, N.; Chamanara, M.; Dehpour, A.; Rashidian, A. The Role of Medicinal Products in the Treatment of Inflammatory Bowel Diseases (IBD) through Inhibition of TLR4/NF-kappaB Pathway. Phytother. Res. PTR 2021, 35, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Okumura, R.; Takeda, K. Roles of Intestinal Epithelial Cells in the Maintenance of Gut Homeostasis. Exp. Mol. Med. 2017, 49, e338. [Google Scholar] [CrossRef] [PubMed]
- Scharek, L.; Tedin, K. The Porcine Immune System—Differences Compared to Man and Mouse and Possible Consequences for Infections by Salmonella Serovars. Berl. Munch. Tierarztl. Wochenschr. 2007, 120, 347–354. [Google Scholar] [CrossRef]
- Skjolaas, K.A.; Burkey, T.E.; Dritz, S.S.; Minton, J.E. Effects of Salmonella enterica Serovars Typhimurium (ST) and Choleraesuis (SC) on Chemokine and Cytokine Expression in Swine Ileum and Jejunal Epithelial Cells. Vet. Immunol. Immunopathol. 2006, 111, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Støy, A.C.F.; Heegaard, P.M.H.; Sangild, P.T.; Østergaard, M.V.; Skovgaard, K. Gene Expression Analysis of the IPEC-J2 Cell Line: A Simple Model for the Inflammation-Sensitive Preterm Intestine. Int. Sch. Res. Not. 2013, 2013, 980651. [Google Scholar] [CrossRef][Green Version]
- Marks, H.; Grześkowiak, Ł.; Martinez-Vallespin, B.; Dietz, H.; Zentek, J. Porcine and Chicken Intestinal Epithelial Cell Models for Screening Phytogenic Feed Additives-Chances and Limitations in Use as Alternatives to Feeding Trials. Microorganisms 2022, 10, 629. [Google Scholar] [CrossRef]
- Hejna, M.; Kovanda, L.; Rossi, L.; Liu, Y. Mint Oils: In Vitro Ability to Perform Anti-Inflammatory, Antioxidant, and Antimicrobial Activities and to Enhance Intestinal Barrier Integrity. Antioxidants 2021, 10, 1004. [Google Scholar] [CrossRef]
- Li, E.; Horn, N.; Ajuwon, K.M. Mechanisms of Deoxynivalenol-Induced Endocytosis and Degradation of Tight Junction Proteins in Jejunal IPEC-J2 Cells Involve Selective Activation of the MAPK Pathways. Arch. Toxicol. 2021, 95, 2065–2079. [Google Scholar] [CrossRef]
- Ling, K.-H.; Wan, M.L.Y.; El-Nezami, H.; Wang, M. Protective Capacity of Resveratrol, a Natural Polyphenolic Compound, against Deoxynivalenol-Induced Intestinal Barrier Dysfunction and Bacterial Translocation. Chem. Res. Toxicol. 2016, 29, 823–833. [Google Scholar] [CrossRef]
- Pomothy, J.M.; Barna, R.F.; Pászti, E.A.; Babiczky, Á.; Szóládi, Á.; Jerzsele, Á.; Gere, E.P. Beneficial Effects of Rosmarinic Acid on IPEC-J2 Cells Exposed to the Combination of Deoxynivalenol and T-2 Toxin. Mediat. Inflamm. 2020, 2020, 8880651. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- International Olive Council. Determination of Biophenols in Olive Oils by HPLC. Available online: https://www.internationaloliveoil.org/wp-content/uploads/2022/10/COI-T20-Doc.-29-REV-1-2017-Eng.pdf (accessed on 21 December 2023).
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Razzuoli, E.; Vencia, W.; Modesto, P.; Franzoni, G.; Giudici, S.D.; Parisi, E.; Ferrari, A.; Amadori, M. Yersinia enterocolitica-Specific Modulation of Innate Immune Responses in Jejunal Epithelial Cells. Vet. Microbiol. 2020, 242, 108596. [Google Scholar] [CrossRef] [PubMed]
- Pistol, G.C.; Bulgaru, C.V.; Marin, D.E.; Oancea, A.G.; Taranu, I. Dietary Grape Seed Meal Bioactive Compounds Alleviate Epithelial Dysfunctions and Attenuates Inflammation in Colon of DSS-Treated Piglets. Foods 2021, 10, 530. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Li, F.; Liu, X.; Liu, J.; Li, D. Synergistic Anti-Inflammatory Effects of Quercetin and Catechin via Inhibiting Activation of TLR4–MyD88-Mediated NF-κB and MAPK Signaling Pathways. Phytother. Res. 2019, 33, 756–767. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, M.; Ceballos-Olvera, I.; del Barrio, L.; Re, F. Role of the Inflammasome, IL-1β, and IL-18 in Bacterial Infections. Sci. World J. 2011, 11, 2037–2050. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Overview of the IL-1 Family in Innate Inflammation and Acquired Immunity. Immunol. Rev. 2018, 281, 8–27. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, N.; Kurata, M.; Yamamoto, T.; Morikawa, S.; Masumoto, J. The Role of Interleukin-1 in General Pathology. Inflamm. Regen. 2019, 39, 12. [Google Scholar] [CrossRef]
- Olcum, M.; Tastan, B.; Ercan, I.; Eltutan, I.B.; Genc, S. Inhibitory Effects of Phytochemicals on NLRP3 Inflammasome Activation: A Review. Phytomedicine 2020, 75, 153238. [Google Scholar] [CrossRef]
- Serrano-López, J.; Martín-Antonio, B. Inflammaging, an Imbalanced Immune Response that Needs to Be Restored for Cancer Prevention and Treatment in the Elderly. Cells 2021, 10, 2562. [Google Scholar] [CrossRef]
- Wang, T.; Xu, H.; Dong, R.; Wu, S.; Guo, Y.; Wang, D. Effectiveness of Targeting the NLRP3 Inflammasome by Using Natural Polyphenols: A Systematic Review of Implications on Health Effects. Food Res. Int. 2023, 165, 112567. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.-T.; Yan, W.-H.; Cao, Y.; Yan, J.-K.; Cai, W. Neutralization of IL-6 and TNF-α Ameliorates Intestinal Permeability in DSS-Induced Colitis. Cytokine 2016, 83, 189–192. [Google Scholar] [CrossRef] [PubMed]
- Franchi, L. Role of Inflammasomes in Salmonella Infection. Front. Microbiol. 2011, 2, 8. [Google Scholar] [CrossRef]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [PubMed]
- Gessner, D.K.; Ringseis, R.; Siebers, M.; Keller, J.; Kloster, J.; Wen, G.; Eder, K. Inhibition of the Pro-Inflammatory NF-κB Pathway by a Grape Seed and Grape Marc Meal Extract in Intestinal Epithelial Cells. J. Anim. Physiol. Anim. Nutr. 2012, 96, 1074–1083. [Google Scholar] [CrossRef] [PubMed]
- González, R.; Ballester, I.; López-Posadas, R.; Suárez, M.D.; Zarzuelo, A.; Martínez-Augustin, O.; Medina, F.S.D. Effects of Flavonoids and Other Polyphenols on Inflammation. Crit. Rev. Food Sci. Nutr. 2011, 51, 331–362. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Tyagi, A.K.; Deshmukh-Taskar, P.; Hinojosa, M.; Prasad, S.; Aggarwal, B.B. Downregulation of Tumor Necrosis Factor and Other Proinflammatory Biomarkers by Polyphenols. Arch. Biochem. Biophys. 2014, 559, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Martín, A.R.; Villegas, I.; La Casa, C.; de la Lastra, C.A. Resveratrol, a Polyphenol Found in Grapes, Suppresses Oxidative Damage and Stimulates Apoptosis during Early Colonic Inflammation in Rats. Biochem. Pharmacol. 2004, 67, 1399–1410. [Google Scholar] [CrossRef] [PubMed]
- Camuesco, D.; Comalada, M.; Concha, A.; Nieto, A.; Sierra, S.; Xaus, J.; Zarzuelo, A.; Gálvez, J. Intestinal Anti-Inflammatory Activity of Combined Quercitrin and Dietary Olive Oil Supplemented with Fish Oil, Rich in EPA and DHA (n-3) Polyunsaturated Fatty Acids, in Rats with DSS-Induced Colitis. Clin. Nutr. 2006, 25, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Salh, B.; Assi, K.; Templeman, V.; Parhar, K.; Owen, D.; Gómez-Muñoz, A.; Jacobson, K. Curcumin Attenuates DNB-Induced Murine Colitis. Am. J. Physiol.-Gastrointest. Liver Physiol. 2003, 285, G235–G243. [Google Scholar] [CrossRef] [PubMed]
- Taranu, I.; Habeanu, M.; Gras, M.A.; Pistol, G.C.; Lefter, N.; Palade, M.; Ropota, M.; Chedea, V.S.; Marin, D.E. Assessment of the Effect of Grape Seed Cake Inclusion in the Diet of Healthy Fattening-Finishing Pigs. J. Anim. Physiol. Anim. Nutr. 2018, 102, e30–e42. [Google Scholar] [CrossRef]
- Britti, D.; Impellizzeri, D.; Procopio, A.; Cuzzocrea, S.; Britti, D.; Impellizzeri, D.; Procopio, A.; Cuzzocrea, S. Oleuropein an Olive Oil Compound in Acute and Chronic Inflammation Models: Facts and Perspectives. In Olive Germplasm—The Olive Cultivation, Table Olive and Olive Oil Industry in Italy; IntechOpen: London, UK, 2012; ISBN 978-953-51-0883-2. [Google Scholar]
- Letterio, J.J.; Roberts, A.B. Regulation of Immune Responses by TGF-Beta. Annu. Rev. Immunol. 1998, 16, 137–161. [Google Scholar] [CrossRef]
- Qin, G.; Zhao, Y.; Gan, Y.; Yu, X.; Zhao, Y.; Peng, H.; Fang, S. Alterations in Gene Expressions of Caco-2 Cell Responses to LPS and Ploy(I:C) Stimulation. PeerJ 2023, 11, e15459. [Google Scholar] [CrossRef]
- Huang, T.; Huang, X.; Shi, B.; Wang, F.; Feng, W.; Yao, M. Regulators of Salmonella-Host Interaction Identified by Peripheral Blood Transcriptome Profiling: Roles of TGFB1 and TRP53 in Intracellular Salmonella Replication in Pigs. Vet. Res. 2018, 49, 121. [Google Scholar] [CrossRef]
- Howe, K.L.; Reardon, C.; Wang, A.; Nazli, A.; McKay, D.M. Transforming Growth Factor-β Regulation of Epithelial Tight Junction Proteins Enhances Barrier Function and Blocks Enterohemorrhagic Escherichia coli O157:H7-Induced Increased Permeability. Am. J. Pathol. 2005, 167, 1587–1597. [Google Scholar] [CrossRef]
- Nallathambi, R.; Poulev, A.; Zuk, J.B.; Raskin, I. Proanthocyanidin-Rich Grape Seed Extract Reduces Inflammation and Oxidative Stress and Restores Tight Junction Barrier Function in Caco-2 Colon Cells. Nutrients 2020, 12, 1623. [Google Scholar] [CrossRef]
- Laurindo, L.F.; Santos, A.R.d.O.d.; Carvalho, A.C.A.d.; Bechara, M.D.; Guiguer, E.L.; Goulart, R.d.A.; Vargas Sinatora, R.; Araújo, A.C.; Barbalho, S.M. Phytochemicals and Regulation of NF-kB in Inflammatory Bowel Diseases: An Overview of In Vitro and In Vivo Effects. Metabolites 2023, 13, 96. [Google Scholar] [CrossRef] [PubMed]
- Mecocci, S.; De Paolis, L.; Zoccola, R.; Fruscione, F.; De Ciucis, C.G.; Chiaradia, E.; Moccia, V.; Tognoloni, A.; Pascucci, L.; Zoppi, S.; et al. Antimicrobial and Immunomodulatory Potential of Cow Colostrum Extracellular Vesicles (ColosEVs) in an Intestinal In Vitro Model. Biomedicines 2022, 10, 3264. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences | Amplicon Length | Source |
| IL18 | For-5′-CGTGTTTGAGGATATGCCTGATT-3′ Rev-5′-TGGTTACTGCCAGACCTCTAGTGA-3′ | 106 | [48] |
| IL1B | For-5′-AATTCGAGTCTGCCCTGTACCC-3′ Rev-5′-TGGTGAAGTCGGTTATATCTTGGC-3′ | 110 | [49] |
| NOS2 | For-5′-CGTTATGCCACCAACAATGG-3′ Rev-5′-AGACCCGGAAGTCGTGCTT-3′ | 84 | [48] |
| TGFB1 | For-5′-CGCGTGCTAATGGTGGAAAG-3′ Rev-5′-CCGACGTGTTGAACAGCATA-3′ | 87 | [48] |
| CXCL8 | For-5′-TTCGATGCCAGTGCATAAATA-3′ Rev-5′-CTGTACAACCTTCTGCACCCA-3′ | 175 | [69] |
| MYD88 | For-5′-GCAGCTGGAACAGACCAACT-3′ Rev-5′-GTGCCAGGCAGGACATCT-3′ | 62 | [69] |
| NFKB1 | For-5′-CCCATGTAGACAGCACCACCTATGAT-3′ Rev-5′-ACAGAGGCTCAAAGTTCTCCACCA-3′ | 131 | [69] |
| NFKB/p65 | For-5′-CGAGAGGAGCACGGATACCA-3′ Rev-5′-GCCCCGTGTAGCCATTGA-3′ | 61 | [69] |
| DEFB1 | For-5′-CTGTTAGCTGCTTAAGGAATAAAGGC-3′ Rev-5′-TGCCACAGGTGCCGATCT-3′ | 80 | [48] |
| DEFB4A | For-5′-CCAGAGGTCCGACCACTA-3′ Rev-5′-GGTCCCTTCAATCCTGTT-3′ | 87 | [48] |
| TLR4 | For-5′-TGGCAGTTTCTGAGGAGTCATG-3′ Rev.–5′ –CCGCAGCAGGGACTTCTC-3′ | 71 | [48] |
| TLR5 | For-5′-TCAAAGATCCTGACCATCACA-3′ Rev.-5′ –CCAGCTGTATCAGGGAGCTT-3′ | 59 | [48] |
| GAPDH | For-5′-ATGGTGAAGGTCGGAGTGAA-3′ Rev-5′AGTGGAGGTCAATGAAGGGG-3′ | 61 | [48] |
| HPRT1 | For-5′-AACCTTGCTTTCCTTGGTCA-3′ Rev-5′-TCAAGGGCATAGCCTACCAC-3′ | 150 | [48] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferlisi, F.; De Ciucis, C.G.; Trabalza-Marinucci, M.; Fruscione, F.; Mecocci, S.; Franzoni, G.; Zinellu, S.; Galarini, R.; Razzuoli, E.; Cappelli, K. Olive Mill Waste-Water Extract Enriched in Hydroxytyrosol and Tyrosol Modulates Host–Pathogen Interaction in IPEC-J2 Cells. Animals 2024, 14, 564. https://doi.org/10.3390/ani14040564
Ferlisi F, De Ciucis CG, Trabalza-Marinucci M, Fruscione F, Mecocci S, Franzoni G, Zinellu S, Galarini R, Razzuoli E, Cappelli K. Olive Mill Waste-Water Extract Enriched in Hydroxytyrosol and Tyrosol Modulates Host–Pathogen Interaction in IPEC-J2 Cells. Animals. 2024; 14(4):564. https://doi.org/10.3390/ani14040564
Chicago/Turabian StyleFerlisi, Flavia, Chiara Grazia De Ciucis, Massimo Trabalza-Marinucci, Floriana Fruscione, Samanta Mecocci, Giulia Franzoni, Susanna Zinellu, Roberta Galarini, Elisabetta Razzuoli, and Katia Cappelli. 2024. "Olive Mill Waste-Water Extract Enriched in Hydroxytyrosol and Tyrosol Modulates Host–Pathogen Interaction in IPEC-J2 Cells" Animals 14, no. 4: 564. https://doi.org/10.3390/ani14040564
APA StyleFerlisi, F., De Ciucis, C. G., Trabalza-Marinucci, M., Fruscione, F., Mecocci, S., Franzoni, G., Zinellu, S., Galarini, R., Razzuoli, E., & Cappelli, K. (2024). Olive Mill Waste-Water Extract Enriched in Hydroxytyrosol and Tyrosol Modulates Host–Pathogen Interaction in IPEC-J2 Cells. Animals, 14(4), 564. https://doi.org/10.3390/ani14040564