
Feeding Postures and Substrate Use of François’ Langurs (Trachypithecus francoisi) in the Limestone Forest of Southwest China

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Subjects
2.2. Data Collection and Analysis
3. Results
3.1. Dietary Composition and Their Seasonal Variation
3.2. Feeding Postures and Their Seasonal Variation
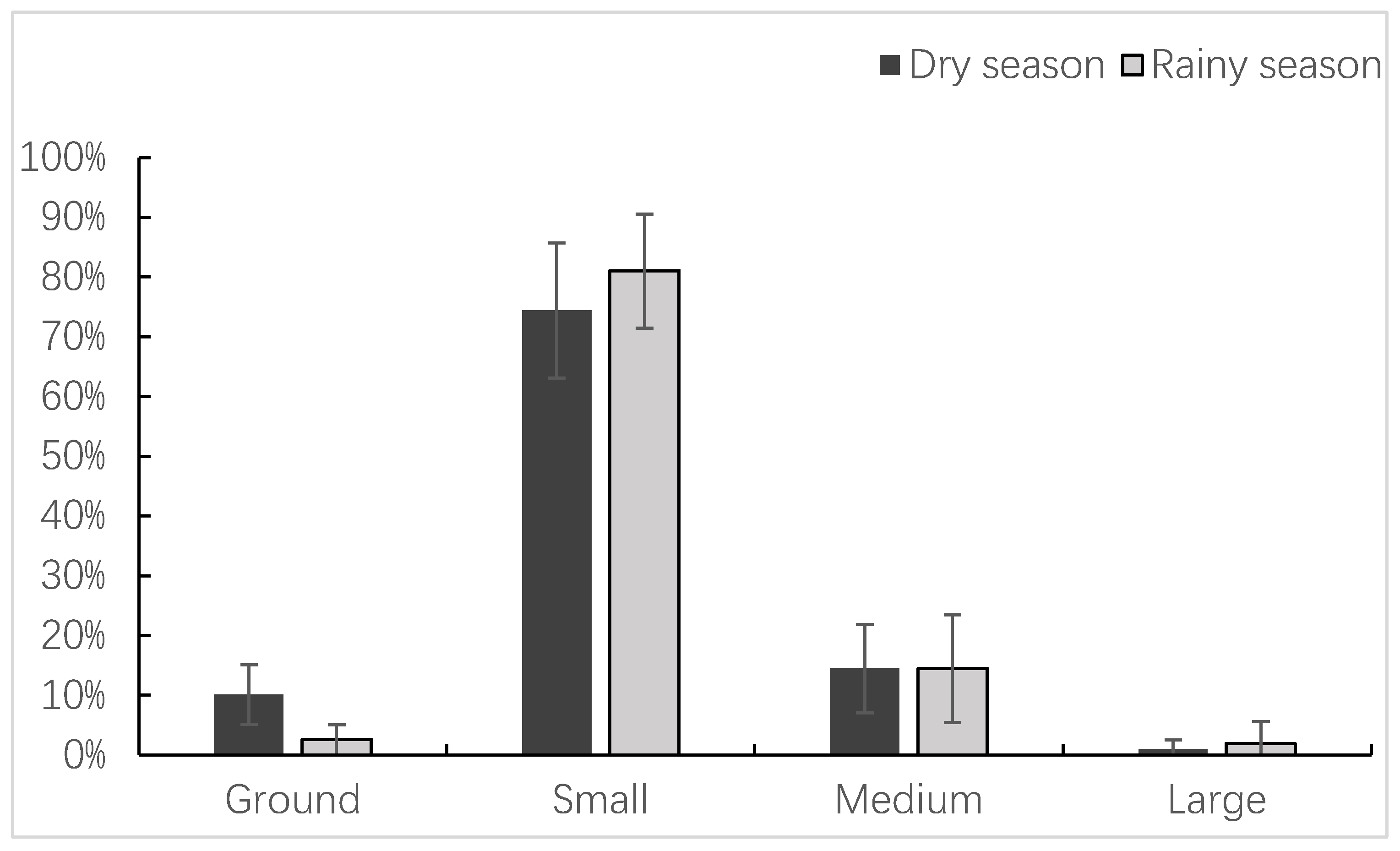
3.3. Forest Strata Use by François’ Langurs during Feeding
4. Discussions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Workman, C.; Schmitt, D. Positional behavior of Delacour’s langurs (Trachypithecus delacouri) in Northern Vietnam. Int. J. Primatol. 2012, 33, 19–37. [Google Scholar] [CrossRef]
- Fleagle, J.G. Primate Adaptation and Evolution, 3rd ed.; Academic Press: London, UK, 2013. [Google Scholar]
- Mekonnen, A.; Fashing, P.J.; Sargis, E.J.; Venkataraman, V.V.; Bekele, A.; Hernandez-Aguilar, R.A.; Rueness, E.K.; Stenseth, N.C. Flexibility in positional behavior, strata use, and substrate utilization among Bale monkeys (Chlorocebus djamdjamensis) in response to habitat fragmentation and degradation. Am. J. Primatol. 2018, 80, e22760. [Google Scholar] [CrossRef]
- Richard, A.F. Primates in Nature; W. H. Freeman: New York, NY, USA, 1985. [Google Scholar]
- Cant, J.G.H. Positional behavior and body size of arboreal primates: A theoretical framework for field studies and an illustration of its application. Am. J. Phys. Anthropol. 1992, 88, 273–283. [Google Scholar] [CrossRef]
- Hunt, K.D. Social rank and body size as determinants of positional behavior in Pan troglodytes. Primates 1992, 33, 347–357. [Google Scholar] [CrossRef]
- Granatosky, M.C. A review of locomotor diversity in mammals with analyses exploring the influence of substrate use, body mass and intermembral index in primates. J. Zool. 2018, 306, 207–216. [Google Scholar] [CrossRef]
- Garber, P.A.; Pruetz, J.D. Positional behavior in moustached tamarin monkeys: Effects of habitat on locomotor variability and locomotor stability. J. Hum. Evol. 1995, 28, 411–426. [Google Scholar] [CrossRef]
- Zhu, W.W.; Garber, P.A.; Bezanson, M.; Qi, X.G.; Li, B.G. Age- and sex-based patterns of positional behavior and substrate utilization in the golden snub-nosed monkey (Rhinopithecus roxellana). Am. J. Primatol. 2015, 77, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Dunham, N.T. Ontogeny of positional behavior and support use among Colobus angolensis palliatus of the Diani Forest, Kenya. Primates 2015, 56, 183–192. [Google Scholar] [CrossRef]
- Dagosto, M.; Yamashita, N. Effect of habitat structure on positional behavior and support use in three species of lemur. Primates 1998, 39, 459–472. [Google Scholar] [CrossRef]
- Prates, H.M.; Bicca-Marques, J.C. Age-sex analysis of activity budget, diet, and positional behavior in Alouatta caraya in an orchard forest. Int. J. Primatol. 2008, 29, 703–715. [Google Scholar] [CrossRef]
- Chen, T.; Huang, Z.; Huang, C.; Wei, H.; Zhou, Q. Positional behaviours of François’ langur (Trachypithecus francoisi) in the limestone forest of Nonggang, Guangxi, South-West China. Folia Primatol. 2020, 91, 170–187. [Google Scholar] [CrossRef]
- Gebo, D.L.; Chapman, C.A. Habitat, annual, and seasonal effects on positional behavior in red colobus monkeys. Am. J. Phys. Anthropol. 1995, 96, 73–82. [Google Scholar] [CrossRef]
- McGraw, W.S. Positional behavior of Cercopithecus petaurista. Int. J. Primatol. 2000, 21, 157–182. [Google Scholar] [CrossRef]
- Li, Y.; Huang, X.; Huang, Z. Behavioral adjustments and support use of Francois’ langur in limestone habitat in Fusui, China: Implications for behavioral thermoregulation. Ecol. Evol. 2020, 10, 4956–4967. [Google Scholar] [CrossRef]
- Bezanson, M. Primate positional behavior development and evolution. Annu. Rev. Anthropol. 2017, 46, 279–298. [Google Scholar] [CrossRef]
- Youlatos, D. Seasonal variation in the positional behavior of red howling monkeys (Alouatta seniculus). Primates 1998, 39, 449–457. [Google Scholar] [CrossRef]
- Huang, C.; Li, Y. How does the white-headed langur (Trachypithecus leucocephalus) adapt locomotor behavior to its unique limestone hill habitat? Primates 2005, 46, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, Y.; Hanya, G.; Grueter, C.C. Feeding strategies of primates in temperate and alpine forests: Comparison of Asian macaques and colobines. Primates 2013, 54, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Aujard, F.; Séguy, M.; Terrien, J.; Botalla, R.; Blanc, S.; Perret, M. Behavioral thermoregulation in a non human primate: Effects of age and photoperiod on temperature selection. Exp. Gerontol. 2006, 41, 784–792. [Google Scholar] [CrossRef]
- Hanya, G.; Kiyono, M.; Hayaishi, S. Behavioral thermoregulation of wild Japanese macaques: Comparisons between two subpopulations. Am. J. Primatol. 2007, 69, 802–815. [Google Scholar] [CrossRef] [PubMed]
- Gestich, C.C.; Caselli, C.B.; Setz, E.Z.F. Behavioural thermoregulation in a small Neotropical primate. Ethology 2014, 120, 331–339. [Google Scholar] [CrossRef]
- Kelley, E.A.; Jablonski, N.G.; ChaPlin, G.; Sussman, R.W.; Kamilar, J.M. Behavioral thermoregulation in Lemur catta: The significance of sunning and huddling behaviors. Am. J. Primatol. 2016, 78, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Campos, F.A.; Fedigan, L.M. Behavioral adaptations to heat stress and water scarcity in white-faced capuchins (Cebus capucinus) in Santa Rosa national park, Costa Rica. Am. J. Phys. Anthropol. 2009, 138, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zhang, K.; Liang, J.; Li, Y.; Huang, Z. Food availability, temperature, and day length drive seasonal variations in the positional behavior of white-headed langurs in the limestone forests of Southwest Guangxi, China. Ecol. Evol. 2021, 11, 14857–14872. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wu, H.; Zhou, Q.; Li, Y.; Cai, X. Feeding strategy of Francois’ langur and white-headed langur at Fusui, China. Am. J. Primatol. 2008, 70, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Huang, C. Advances in ecological research on the limestone langurs in China. Acta Theriol. Sin. 2021, 41, 59–70. [Google Scholar]
- Lu, S.; Chen, T.; Huang, Z.; Li, Y.; Lu, C. Interannual variation in food choice of white-headed langur inhabiting limestone forests in Fusui, southwest Guangxi, China. Ecol. Evol. 2021, 11, 9349–9360. [Google Scholar] [CrossRef] [PubMed]
- Altmann, J. Observational study of behavior: Sampling methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef]
- Hunt, K.D.; Cant, J.G.H.; Gebo, D.L.; Rose, M.D.; Walker, S.E.; Youlatos, D. Standardized descriptions of primate locomotor and postural modes. Primates 1996, 37, 363–387. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 18 May 2021).
- McGraw, W.S. Posture and support use of old world monkeys (Cercopithecidae): The influence of foraging strategies, activity patterns, and the spatial distribution of preferred food items. Am. J. Primatol. 1998, 46, 229–250. [Google Scholar] [CrossRef]
- Clutton-Brock, T.H. Feeding levels and feeding sites of red colobus (Colobus badius tephrosceles) in the Gombe national park. Folia Primatol. 1973, 19, 368–379. [Google Scholar] [CrossRef]
- Youlatos, D. Positinal behavior of black spider monkeys (Ateles paniscus) in French Guiana. Int. J. Primatol. 2002, 23, 1071–1093. [Google Scholar] [CrossRef]
- Huang, Z.; Huang, C.; Wei, H.; Tang, H.; Krzton, A.; Ma, G.; Zhou, Q. Factors influencing positional behavior and habitat use of sympatric macaques in the limestone habitat of Nonggang, China. Int. J. Primatol. 2015, 36, 95–112. [Google Scholar] [CrossRef]
- Li, Y.; Ding, P.; Huang, C.; Jiang, P.; Wood, C. Dietary response of a group of François’ langur Trachypithecus francoisi in a fragmented habitat in the county of Fusui, China: Implications for conservation. Wildl. Biol. 2009, 15, 137–146. [Google Scholar] [CrossRef]
- Dagosto, M. Seasonal variation in positional behavior of Malagasy lemurs. Int. J. Primatol. 1995, 16, 807–833. [Google Scholar] [CrossRef]
- Zhou, Q.; Wei, F.; Huang, C.; Li, M.; Ren, B.; Luo, B. Seasonal variation in the activity patterns and time budgets of Trachypithecus francoisi in the Nonggang nature Reserve, China. Int. J. Primatol. 2007, 28, 657–671. [Google Scholar] [CrossRef]
- Zhou, Q.; Wei, F.; Li, M.; Huang, C.; Luo, B. Diet and food choice of Trachypithecus françoisi in the Nonggang nature Reserve, China. Int. J. Primatol. 2006, 27, 1441–1460. [Google Scholar] [CrossRef]
- Huang, Z.; Huang, C.; Zhou, Q.; Wei, H.; Meng, Y. Diet and the seasonal changes of the François’ langur (Trachypithecus francoisi). Acta Ecol. Sin. 2010, 30, 5501–5508. [Google Scholar]
- Bicca-Marques, J.C.; Calegaro-Marques, C. Behavioral thermoregulation in a sexually and developmentally dichromatic Neotropical primate, the black-and-gold howling monkey (Alouatta caraya). Am. J. Phys. Anthropol. 1998, 106, 533–546. [Google Scholar] [CrossRef]
- Donati, G.; Ricci, E.; Baldi, N.; Morelli, V.; Borgognini-Tarli, S.M. Behavioral thermoregulation in a gregarious lemur, Eulemur collaris: Effects of climatic and dietary-related factors. Am. J. Phys. Anthropol. 2011, 144, 355–364. [Google Scholar] [CrossRef]
- Huang, C. White-Headed Langurs in China; Guangxi Normal University Press: Guilin, China, 2002. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Term | Definition |
|---|---|
| Activities | |
| Resting | The unaltered position of the langur individual |
| Feeding | The foraging, picking, ingestion, and chewing of food |
| Moving | Including quadrupedal walking, leaping, vertical climbing, and quadrupedal running |
| Social grooming | Mutual grooming behavior among the individuals |
| Postural modes | |
| Sitting | An orthograde posture in which the ischia bears a substantial portion of the body weight |
| Bipedal standing | Standing on the hind limbs with no significant support from any other body part. |
| Quadrupedal standing | Four-limbed standing on horizontal or subhorizontal supports; the elbow and knee are relatively extended and the trunk is near horizontal |
| Suspending | A part of the body hangs on the support, including the hind limb. |
| Forest strata height | |
| Ground | On the bare rock |
| Low | Height of ≤5 m of strata |
| Middle | Height of >5 m and ≤10 m |
| Upper | Height of >10 m |
| Substrate size | |
| Small | Substrate diameter of ≤2 cm |
| Medium | Substrate diameter of >2 cm and ≤5 cm |
| Large | Substrate diameter of >5 cm |
| Month | Total Sampling Days | Full Observation Days | Scan Records | Scan Individual | Feeding Records | Feeding Postures (%) | ||
|---|---|---|---|---|---|---|---|---|
| Sitting | Bipedal Standing | Other Postures | ||||||
| 1 | 6 | 5 | 194 | 1029 | 342 | 85.13 | 14.29 | 0.58 |
| 2 | 5 | 4 | 191 | 1345 | 414 | 81.82 | 15.15 | 3.03 |
| 3 | 5 | 5 | 202 | 1415 | 496 | 85.91 | 10.15 | 0.38 |
| 4 | 5 | 5 | 222 | 1584 | 521 | 91.11 | 7.27 | 1.62 |
| 5 | 6 | 5 | 221 | 1284 | 404 | 91.25 | 8.00 | 0.75 |
| 6 | 4 | 4 | 156 | 804 | 337 | 92.04 | 6.78 | 1.18 |
| 7 | 4 | 4 | 144 | 813 | 320 | 89.09 | 10.30 | 0.61 |
| 8 | 5 | 5 | 168 | 1022 | 342 | 89.12 | 9.41 | 1.47 |
| 9 | 2 | 2 | 65 | 370 | 138 | 83.33 | 10.87 | 5.80 |
| 10 | 4 | 4 | 139 | 1054 | 358 | 70.13 | 29.44 | 0.43 |
| 11 | 5 | 4 | 166 | 989 | 393 | 84.93 | 14.87 | 0.20 |
| 12 | 4 | 3 | 135 | 876 | 322 | 87.99 | 11.36 | 0.65 |
| Mean | 4.58 | 4.17 | 166.92 | 1048.75 | 365.58 | 85.99 | 12.33 | 1.39 |
| SD | 1.08 | 0.94 | 44.04 | 326.76 | 97.00 | 5.97 | 6.08 | 1.59 |
| Posture | Statistics (n = 12) | Dietary Composition | ||||
|---|---|---|---|---|---|---|
| Young Leaves | Mature Leaves | Flowers | Fruits | Others | ||
| Sitting | r | 0.510 | −0.580 | −0.126 | 0.133 | −0.399 |
| Adjusted p | 0.225 | 0.225 | 0.697 | 0.697 | 0.332 | |
| Bipedal standing | r | −0.636 | 0.713 | 0.175 | −0.168 | 0.510 |
| Adjusted p | 0.065 | 0.045 | 0.602 | 0.602 | 0.150 | |
| Other postures | r | 0.196 | −0.147 | −0.217 | 0.357 | −0.007 |
| Adjusted p | 0.811 | 0.811 | 0.811 | 0.811 | 0.983 | |
| Month | Forest Strata Heights (%) | Substrate Sizes (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| Ground | Low | Middle | Upper | Ground | Small | Medium | Large | |
| 1 | 16.33 | 60.06 | 20.70 | 2.92 | 19.24 | 53.06 | 23.62 | 4.08 |
| 2 | 9.09 | 66.67 | 18.18 | 6.06 | 9.09 | 75.76 | 15.15 | 0 |
| 3 | 3.81 | 53.43 | 37.94 | 4.82 | 4.31 | 71.95 | 23.10 | 0.63 |
| 4 | 2.58 | 72.54 | 20.84 | 4.04 | 3.39 | 67.21 | 29.24 | 0.16 |
| 5 | 2.75 | 51.50 | 40.25 | 5.50 | 1.25 | 82.50 | 15.25 | 1.00 |
| 6 | 1.18 | 35.99 | 53.69 | 9.14 | 1.77 | 84.07 | 13.57 | 0.59 |
| 7 | 7.58 | 56.97 | 30.00 | 5.45 | 7.27 | 73.94 | 18.48 | 0.30 |
| 8 | 10.00 | 36.76 | 52.94 | 0.29 | 1.76 | 83.53 | 5.29 | 9.41 |
| 9 | 0 | 23.91 | 73.19 | 2.90 | 0 | 94.93 | 5.07 | 0 |
| 10 | 7.79 | 76.62 | 15.15 | 0.43 | 7.79 | 83.55 | 8.66 | 0 |
| 11 | 9.98 | 72.41 | 17.61 | 0 | 9.59 | 79.65 | 9.59 | 1.17 |
| 12 | 10.39 | 64.29 | 23.05 | 2.27 | 10.71 | 82.47 | 6.82 | 0 |
| Mean | 6.79 | 55.93 | 33.63 | 3.65 | 6.35 | 77.72 | 14.49 | 1.45 |
| SD | 4.78 | 16.50 | 18.33 | 2.73 | 5.46 | 10.55 | 7.88 | 2.75 |
| Forest Strata Heights | Statistics (n = 12) | Dietary Composition | ||||
|---|---|---|---|---|---|---|
| Young Leaves | Mature Leaves | Flowers | Fruits | Others | ||
| Ground | r | −0.448 | 0.545 | 0.175 | −0.245 | 0.392 |
| Adjusted p | 0.347 | 0.335 | 0.587 | 0.554 | 0.347 | |
| Low | r | −0.741 | 0.392 | 0.070 | 0.007 | 0.406 |
| Adjusted p | 0.030 | 0.347 | 0.983 | 0.983 | 0.347 | |
| Middle | r | 0.713 | −0.517 | −0.077 | 0.077 | −0.406 |
| Adjusted p | 0.045 | 0.213 | 0.812 | 0.812 | 0.318 | |
| Upper | r | 0.434 | −0.531 | 0.056 | 0.287 | −0.580 |
| Adjusted p | 0.265 | 0.188 | 0.863 | 0.458 | 0.188 | |
| Strata Sizes | Statistics (n = 12) | Dietary Composition | ||||
|---|---|---|---|---|---|---|
| Young Leaves | Mature Leaves | Flowers | Fruits | Others | ||
| Ground | r | −0.650 | 0.524 | 0.084 | −0.049 | 0.329 |
| Adjusted p | 0.110 | 0.200 | 0.880 | 0.880 | 0.495 | |
| Small | r | 0.252 | 0.105 | 0.133 | −0.462 | 0.455 |
| Adjusted p | 0.717 | 0.746 | 0.746 | 0.345 | 0.345 | |
| Medium | r | −0.014 | −0.329 | −0.224 | 0.469 | −0.678 |
| Adjusted p | 0.966 | 0.495 | 0.605 | 0.310 | 0.075 | |
| Large | r | 0.413 | −0.142 | −0.185 | −0.164 | −0.317 |
| Adjusted p | 0.659 | 0.659 | 0.659 | 0.659 | 0.659 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, S.; Lin, N.; Huang, A.; Tong, D.; Liang, Y.; Li, Y.; Lu, C. Feeding Postures and Substrate Use of François’ Langurs (Trachypithecus francoisi) in the Limestone Forest of Southwest China. Animals 2024, 14, 565. https://doi.org/10.3390/ani14040565
Lu S, Lin N, Huang A, Tong D, Liang Y, Li Y, Lu C. Feeding Postures and Substrate Use of François’ Langurs (Trachypithecus francoisi) in the Limestone Forest of Southwest China. Animals. 2024; 14(4):565. https://doi.org/10.3390/ani14040565
Chicago/Turabian StyleLu, Shiyi, Nanxin Lin, Anshu Huang, Dewen Tong, Yongyan Liang, Youbang Li, and Changhu Lu. 2024. "Feeding Postures and Substrate Use of François’ Langurs (Trachypithecus francoisi) in the Limestone Forest of Southwest China" Animals 14, no. 4: 565. https://doi.org/10.3390/ani14040565
APA StyleLu, S., Lin, N., Huang, A., Tong, D., Liang, Y., Li, Y., & Lu, C. (2024). Feeding Postures and Substrate Use of François’ Langurs (Trachypithecus francoisi) in the Limestone Forest of Southwest China. Animals, 14(4), 565. https://doi.org/10.3390/ani14040565