
Optimization of Fermented Maize Stover for the Fattening Phase of Geese: Effect on Production Performance and Gut Microflora
 , , , , , , , , , and
, , , , , , , , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Diet and Experimental Design
2.2. Preparation of Fermented Maize Stover and Determination of Fermentation Quality
2.3. Sample Collection
2.4. Growth Performance Parameter Measurements
2.5. Slaughter Performance Parameter Measurements
2.6. Histological Observation of Jejunum and Ileum
2.7. DNA Extraction and 16S RNA Gene Amplicon Sequencing
2.8. Data Analysis
3. Results
3.1. Growth Performance and Slaughter Performance
3.2. Intestinal Morphology of Jejunum and Ileum
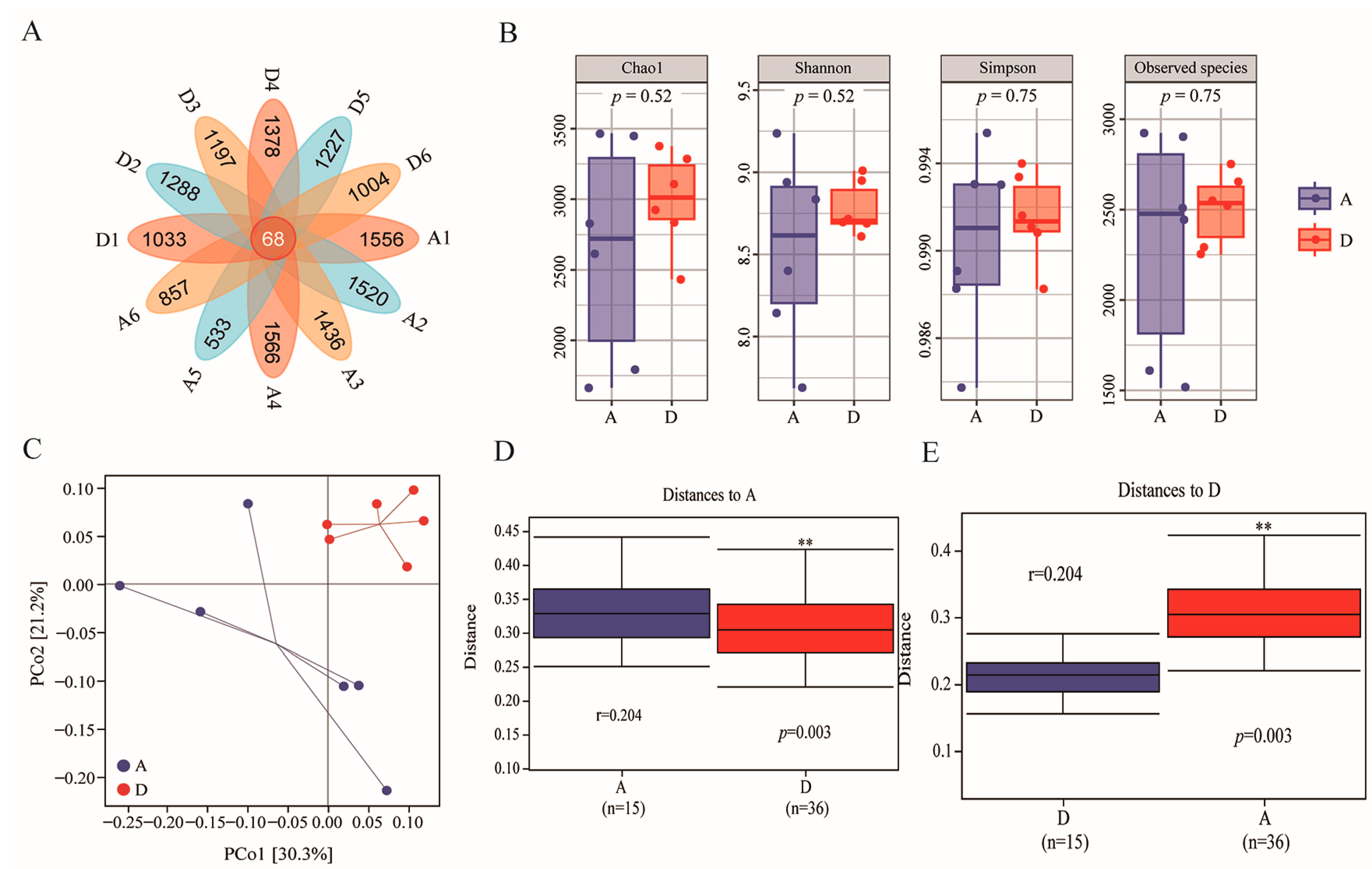
3.3. ASV Composition and Alpha and Beta Diversity Analysis of Caecum Flora
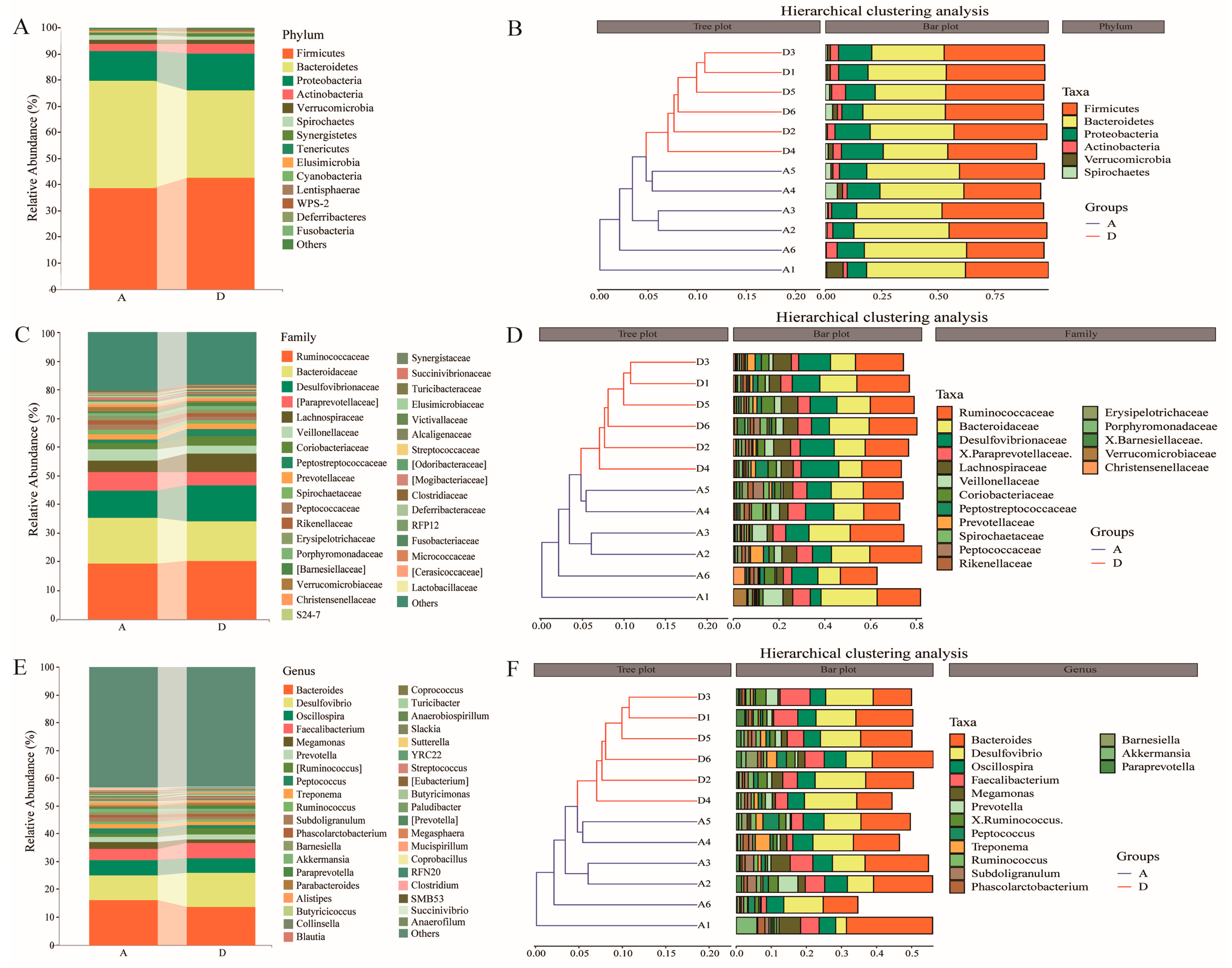
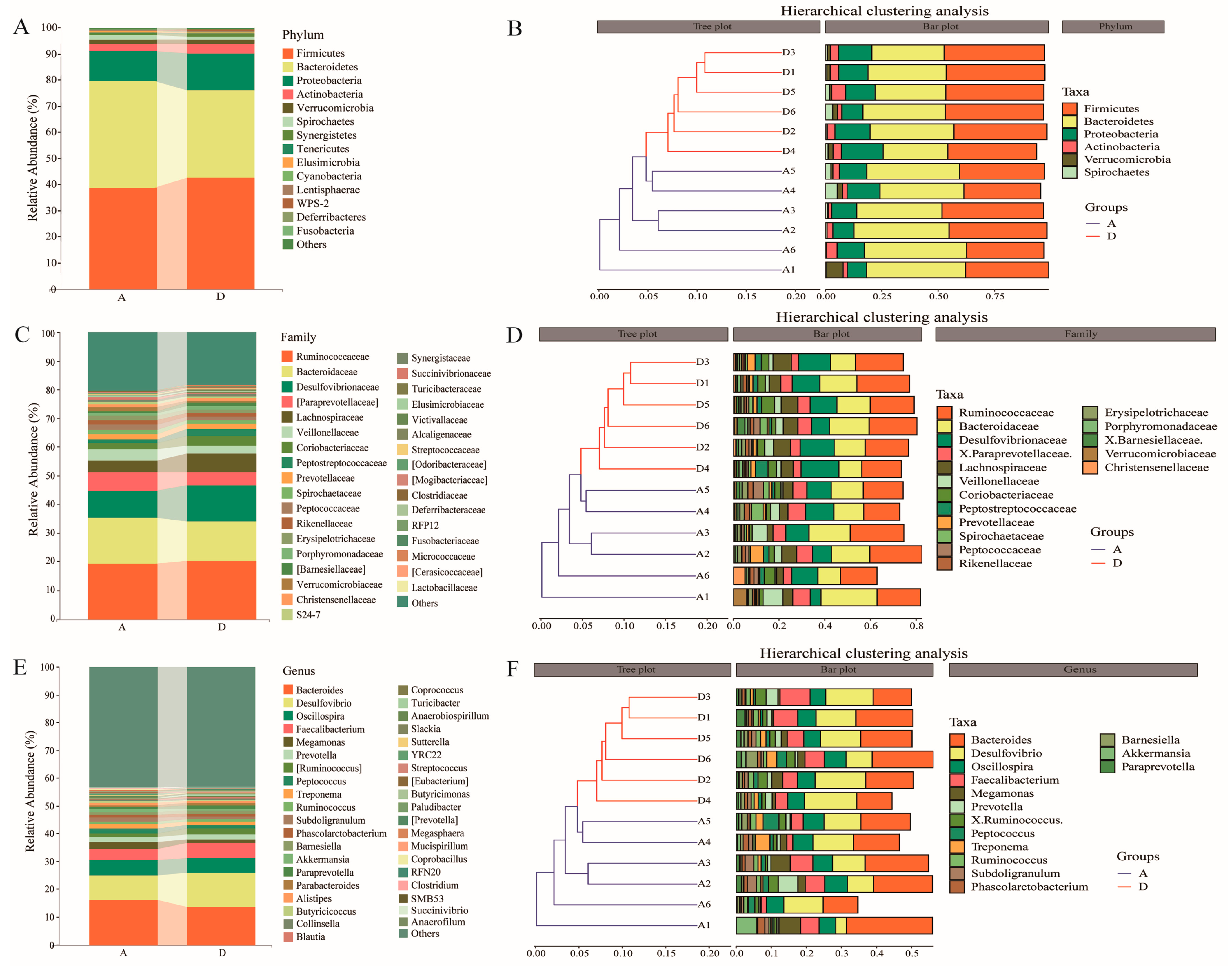
3.4. Taxonomic Composition and Clustering of Intestinal Flora
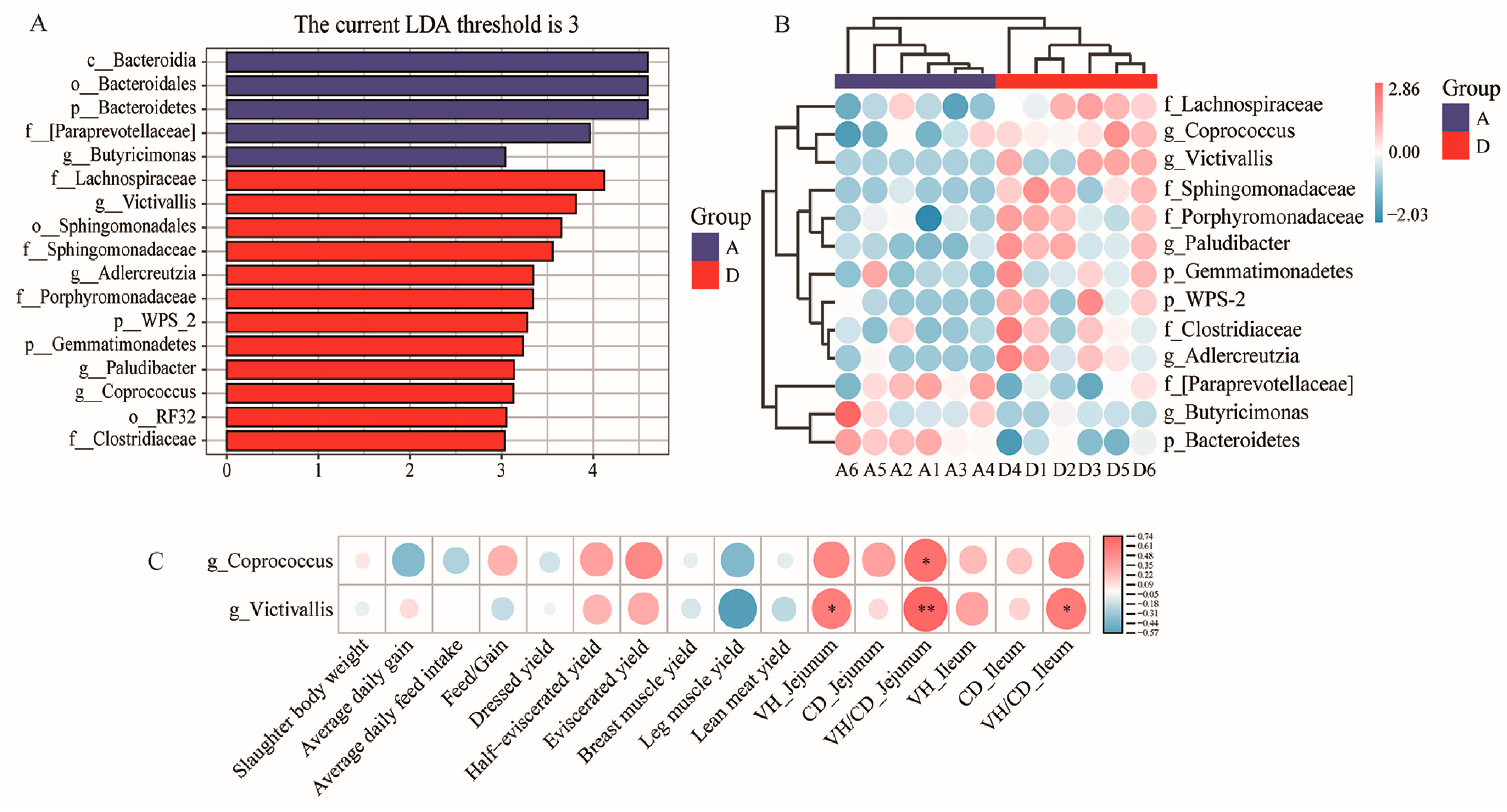
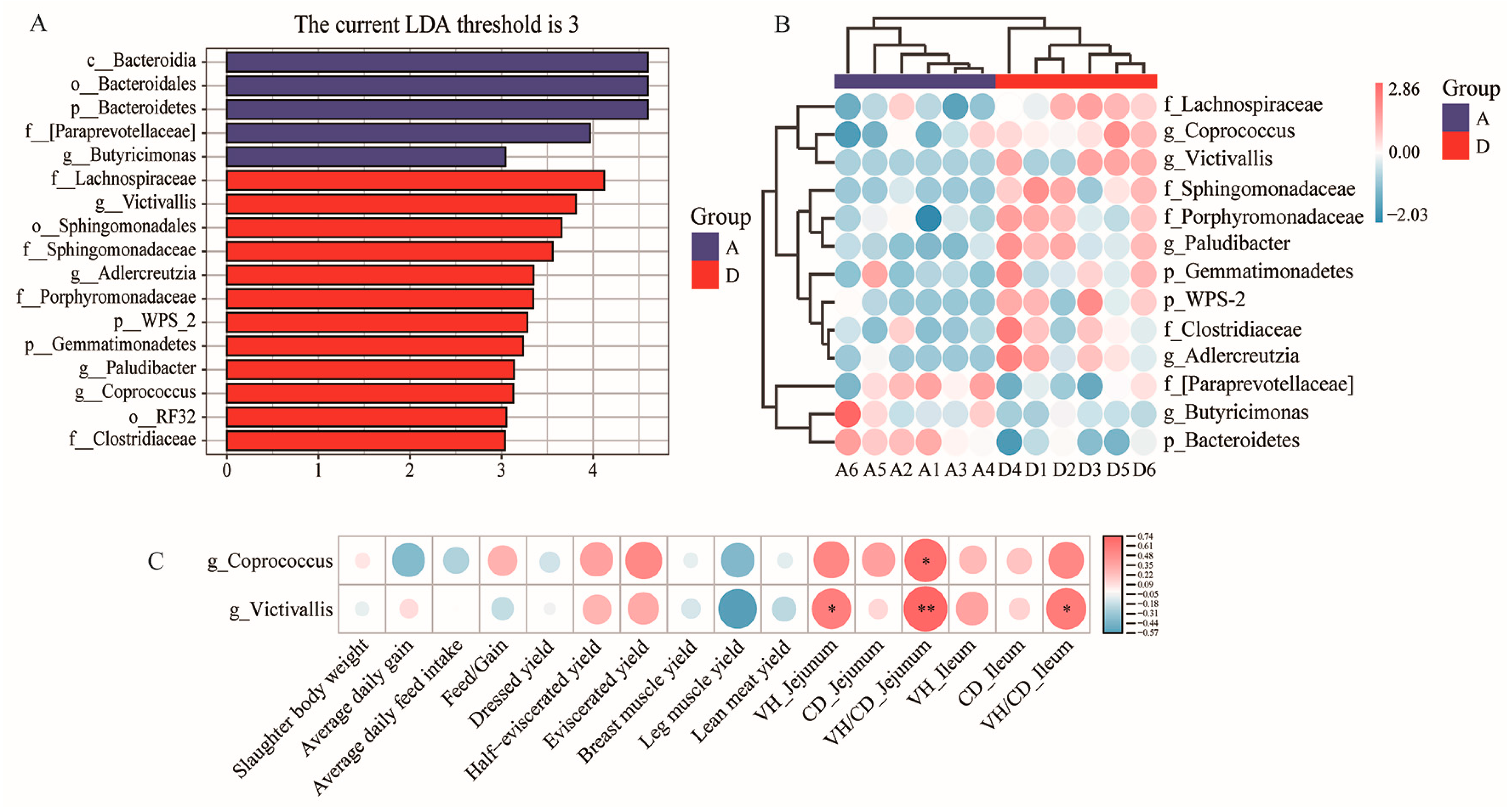
3.5. LDA Effect Size (LEfSe) and Correlation Analysis of Phenotypic Parameters and Differentially Enriched Microbes
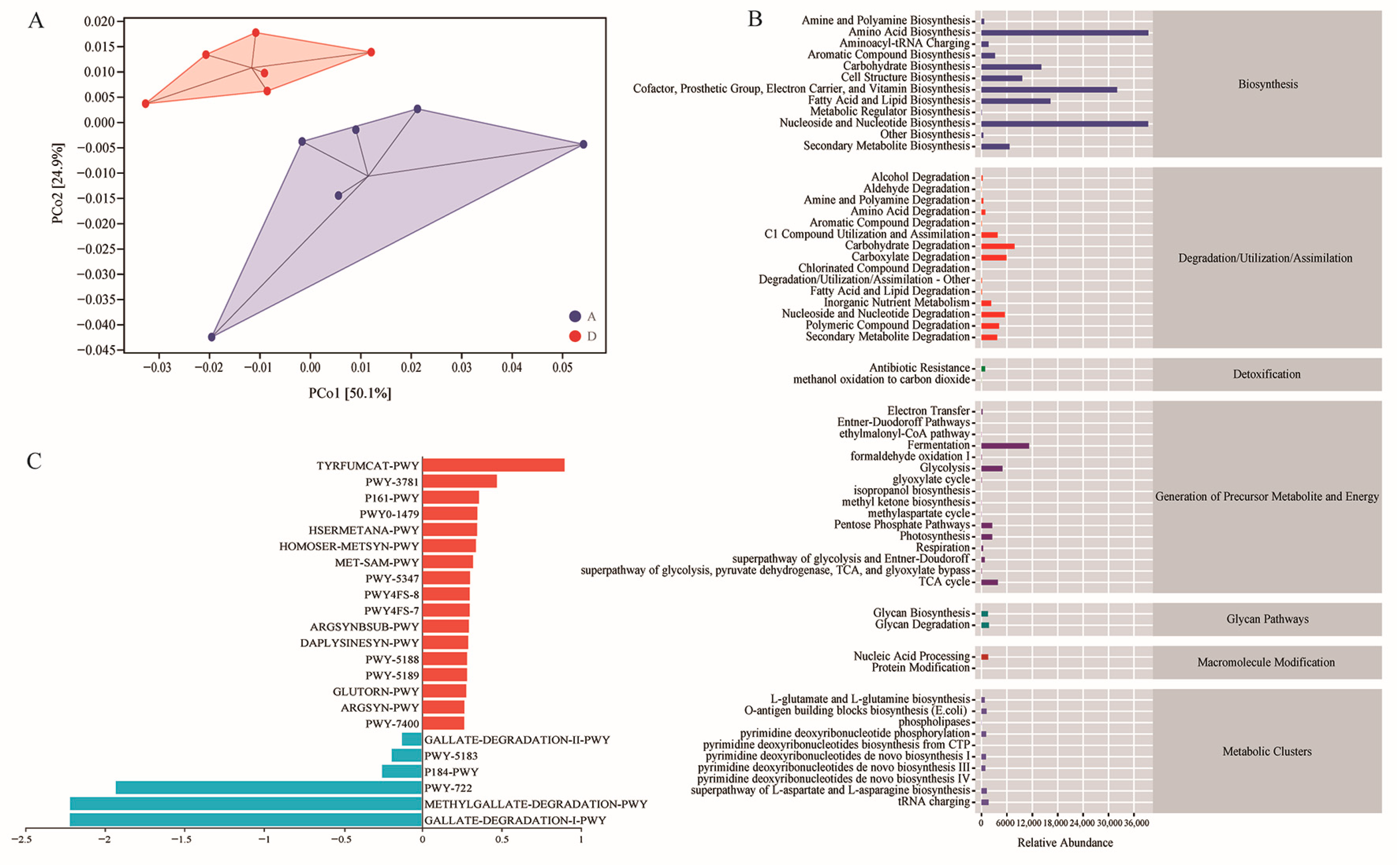
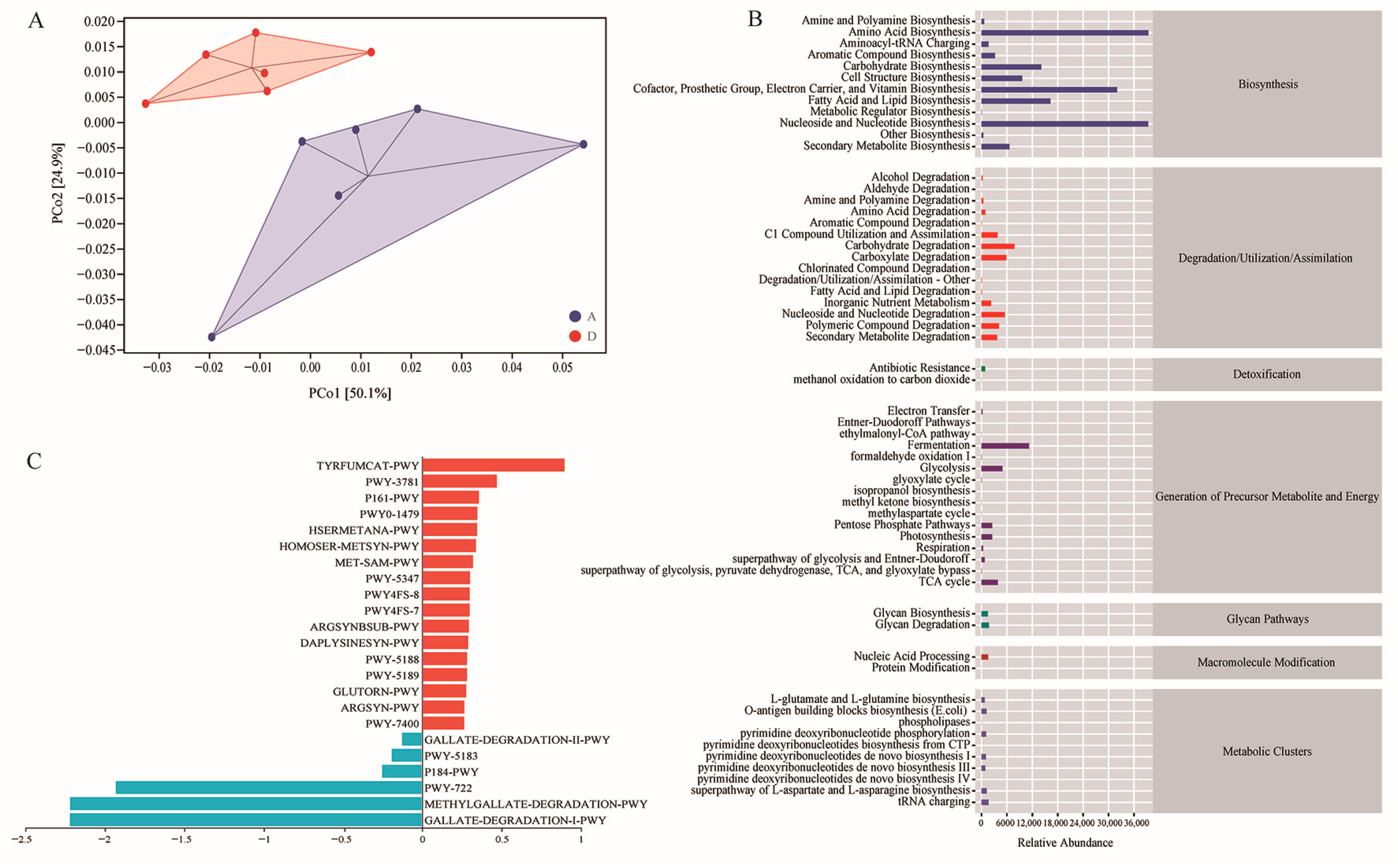
3.6. Functional Potential Prediction of the Intestinal Microbiota
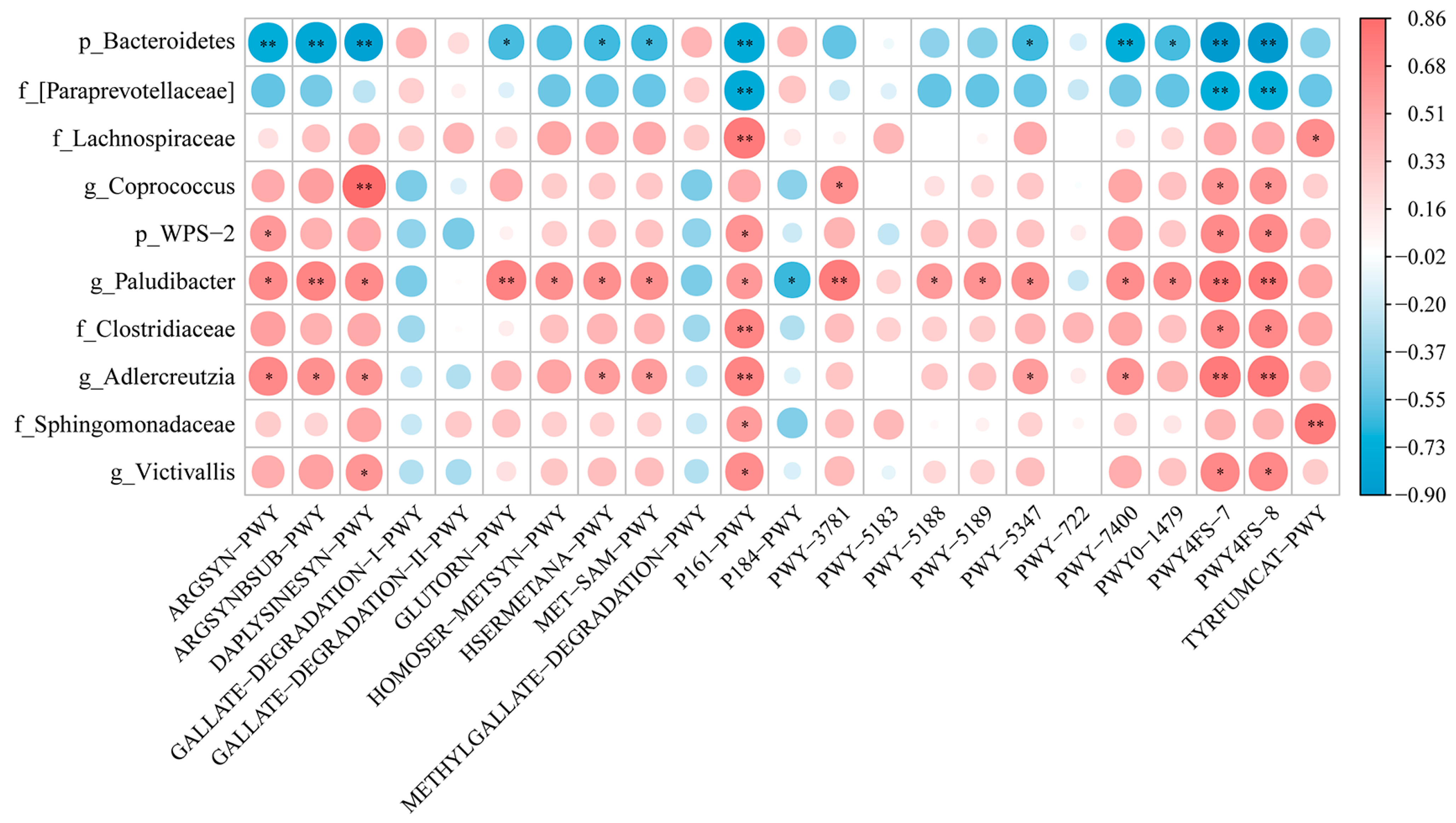
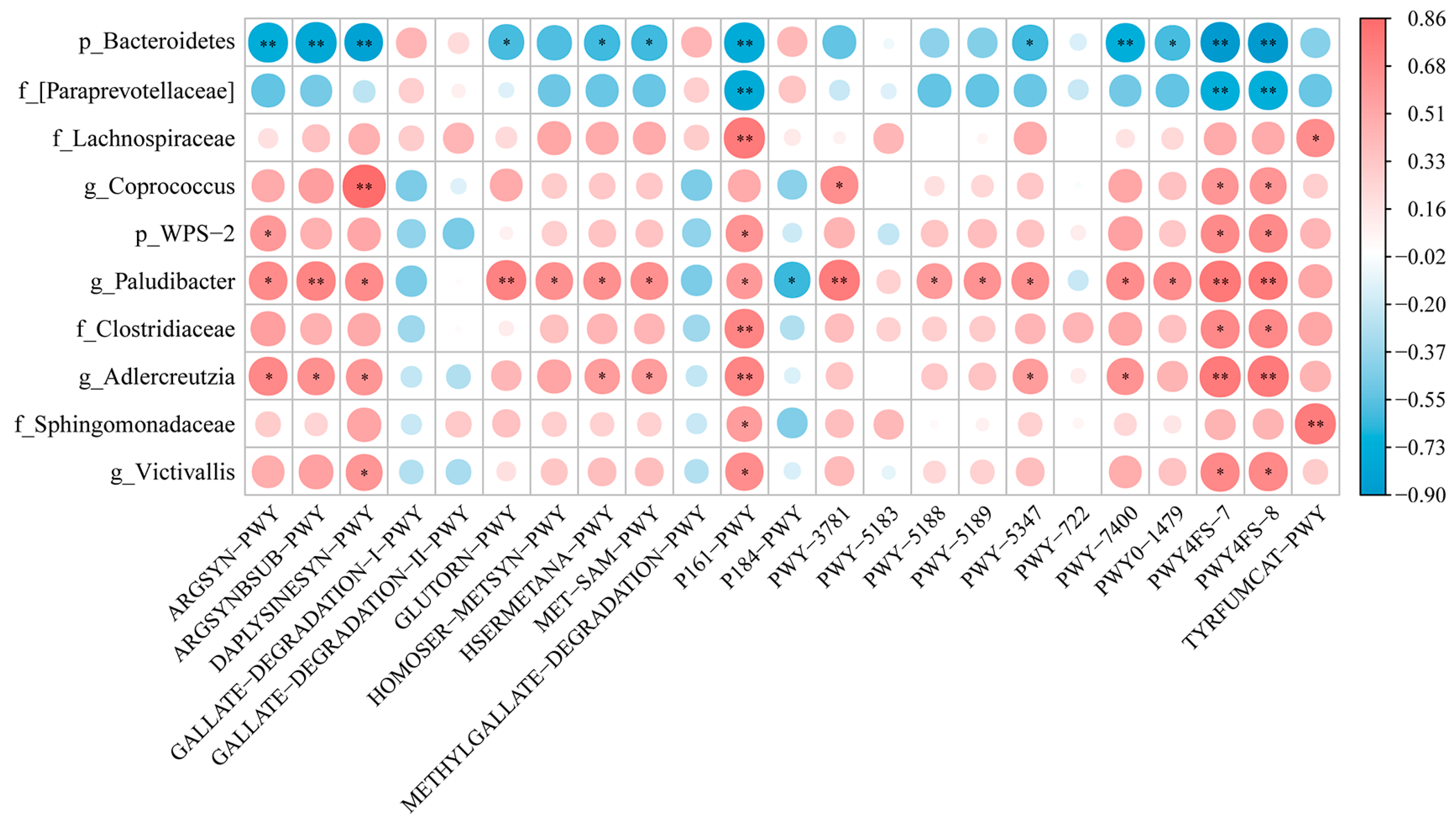
3.7. Differential Bacteria of Gut Microbiota and Differential Metabolic Pathway Enrichment Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aggrey, S.E.; Karnuah, A.B.; Sebastian, B.; Anthony, N.B. Genetic Properties of Feed Efficiency Parameters in Meat-Type Chickens. Genet. Sel. Evol. 2010, 42, 25. [Google Scholar] [CrossRef]
- Carvalho, V.B.; Leite, R.F.; Almeida, M.T.; Paschoaloto, J.R.; Carvalho, E.B.; Lanna, D.P.; Perez, H.L.; Van Cleef, E.H.; Homem Junior, A.C.; Ezequiel, J.M. Carcass Characteristics and Meat Quality of Lambs Fed High Concentrations of Crude Glycerin in Low-Starch Diets. Meat Sci. 2015, 110, 285–292. [Google Scholar] [CrossRef]
- Chen, S.; Li, X.; Liu, X.; Wang, N.; An, Q.; Ye, X.M.; Zhao, Z.T.; Zhao, M.; Han, Y.; Ouyang, K.H.; et al. Investigation of Chemical Composition, Antioxidant Activity, and the Effects of Alfalfa Flavonoids on Growth Performance. Oxid. Med. Cell Longev. 2020, 2020, 8569237. [Google Scholar] [CrossRef]
- Li, M.; Zi, X.; Tang, J.; Xu, T.; Gu, L.; Zhou, H. Effects of Cassava Foliage on Feed Digestion, Meat Quality, and Antioxidative Status of Geese. Poult. Sci. 2020, 99, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Ni, J.J.; Zhao, J.B.; Zhang, G.; Huang, C.F. Regression Equations of Energy Values of Corn, Soybean Meal, and Wheat Bran Developed by Chemical Composition for Growing Pigs. Animals 2020, 10, 1490. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Guo, W.; Gao, F.; Yang, R. Characterization and Pb(Ii) Removal Potential of Corn Straw- and Municipal Sludge-Derived Biochars. R. Soc. Open Sci. 2017, 4, 170402. [Google Scholar] [CrossRef]
- Lu, Z.; Zeng, N.; Jiang, S.; Wang, X.; Yan, H.; Gao, C. Dietary Replacement of Soybean Meal by Fermented Feedstuffs for Aged Laying Hens: Effects on Laying Performance, Egg Quality, Nutrient Digestibility, Intestinal Health, Follicle Development, and Biological Parameters in a Long-Term Feeding Period. Poult. Sci. 2023, 102, 102478. [Google Scholar] [CrossRef] [PubMed]
- Aslan, R.; Öztürk, E. Effects of Maize Silage Feeding on Growth Performance, Carcass Characteristics, Digestive System Length, Chemical Composition, and Meat Quality of Domestic Geese. Trop. Anim. Health Prod. 2022, 54, 325. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.Q.; Cheng, H.Z.; Zhao, D.H.; Yang, J.; Wang, S.B.; Wu, H.Z.; Lu, M.Y.; Xu, L.; Liu, G.J. Effects of Dietary Enteromorpha Powder Supplementation on Productive Performance, Egg Quality, and Antioxidant Performance During the Late Laying Period in Zi Geese. Poult. Sci. 2020, 99, 1062–1068. [Google Scholar] [CrossRef] [PubMed]
- Weimer, P.J. Why Don’t Ruminal Bacteria Digest Cellulose Faster? J. Dairy Sci. 1996, 79, 1496–1502. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Bayer, E.A. Plant Cell Wall Breakdown by Anaerobic Microorganisms from the Mammalian Digestive Tract. Ann. N. Y. Acad. Sci. 2008, 1125, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Yu, Z. Intestinal Microbiome of Poultry and Its Interaction with Host and Diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Mao, P.; Tian, X.; Guo, Q.; Meng, L. Effects of Dietary Supplementation of Alfalfa Meal on Growth Performance, Carcass Characteristics, Meat and Egg Quality, and Intestinal Microbiota in Beijing-You Chicken. Poult. Sci. 2019, 98, 2250–2259. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Luo, X.; Zhao, X.; Zhang, A.; Jiang, N.; Yang, L.; Huang, M.; Xu, L.; Ding, L.; Li, M.; et al. Gut Microbiota Correlates with Fiber and Apparent Nutrients Digestion in Goose. Poult. Sci. 2018, 97, 3899–3909. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.A.; Zhang, D.; Lisle, A.T.; Mikkelsen, D.; McSweeney, C.S.; Kang, S.; Bryden, W.L.; Gidley, M.J. Soluble Arabinoxylan Enhances Large Intestinal Microbial Health Biomarkers in Pigs Fed a Red Meat-Containing Diet. Nutrition 2016, 32, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Lin, X.; Gou, Z.; Fan, Q.; Jiang, S. Effects of Clostridium butyricum, Sodium Butyrate, and Butyric Acid Glycerides on the Reproductive Performance, Egg Quality, Intestinal Health, and Offspring Performance of Yellow-Feathered Breeder Hens. Front. Microbiol. 2021, 12, 657542. [Google Scholar] [CrossRef]
- Li, Z.; Fu, R.; Huang, X.; Wen, X.; Zhang, L. Oral Microbiota May Affect Osteoradionecrosis Following Radiotherapy for Head and Neck Cancer. J. Transl. Med. 2023, 21, 391. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Zhang, H.S.; Feng, Q.D.; Zhang, D.Y.; Zhu, G.L.; Yang, L. Bacterial Community Structure in Geothermal Springs on the Northern Edge of Qinghai-Tibet Plateau. Front. Microbiol. 2022, 13, 994179. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Deng, S.; Hu, S.; Xue, J.; Yang, K.; Zhuo, R.; Xiao, Y.; Fang, R. Productive Performance, Serum Antioxidant Status, Tissue Selenium Deposition, and Gut Health Analysis of Broiler Chickens Supplemented with Selenium and Probiotics—A Pilot Study. Animals 2022, 12, 1086. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic Biomarker Discovery and Explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, W.; Yang, J.; Zhao, H.; Xin, H.; Zhang, Y. Effect of Different Levels of Corn Steep Liquor Addition on Fermentation Characteristics and Aerobic Stability of Fresh Rice Straw Silage. Anim. Nutr. 2016, 2, 345–350. [Google Scholar] [CrossRef] [PubMed]
- He, L.W.; Meng, Q.X.; Li, D.Y.; Zhang, Y.W.; Ren, L.P. Meat Quality, Oxidative Stability and Blood Parameters from Graylag Geese Offered Alternative Fiber Sources in Growing Period. Poult. Sci. 2015, 94, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Ma, Y.; Ding, S.; Fang, J.; Liu, G. Regulation of Dietary Fiber on Intestinal Microorganisms and Its Effects on Animal Health. Anim. Nutr. 2023, 14, 356–369. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zhou, B.; Xi, Y.; Huan, H.; Li, M.; Yu, J.; Zhu, H.; Dai, Z.; Ying, S.; Zhou, W.; et al. Fermented Feed Regulates Growth Performance and the Cecal Microbiota Community in Geese. Poult. Sci. 2019, 98, 4673–4684. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Ma, C.; Fan, X.; Shah, A.M.; Mao, J. Use of Condensed Molasses Fermentation Solubles as an Alternative Source of Concentrates in Dairy Cows. Anim. Biosci. 2021, 34, 205–212. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, H.F.; Gao, L.X.; Zhao, F.; Lu, Q.P.; Sa, R.N. Effect of Graded Levels of Fiber from Alfalfa Meal on Intestinal Nutrient and Energy Flow, and Hindgut Fermentation in Growing Pigs. J. Anim. Sci. 2013, 91, 4757–4764. [Google Scholar] [CrossRef]
- Rodriguez-Sanchez, R.; Tres, A.; Sala, R.; Guardiola, F.; Barroeta, A.C. Evolution of Lipid Classes and Fatty Acid Digestibility Along the Gastrointestinal Tract of Broiler Chickens Fed Different Fat Sources at Different Ages. Poult. Sci. 2019, 98, 1341–1353. [Google Scholar] [CrossRef]
- Jazi, V.; Boldaji, F.; Dastar, B.; Hashemi, S.R.; Ashayerizadeh, A. Effects of Fermented Cottonseed Meal on the Growth Performance, Gastrointestinal Microflora Population and Small Intestinal Morphology in Broiler Chickens. Br. Poult. Sci. 2017, 58, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, W.F.; Liu, S.Z.; Wang, H.H. Probiotic Fermented Feed Improved the Production, Health and Nutrient Utilisation of Yellow-Feathered Broilers Reared in High Altitude in Tibet. Br. Poult. Sci. 2020, 61, 746–753. [Google Scholar] [CrossRef]
- Jin, L.; Zheng, L.Y.; Yang, L.; Li, A.; Gao, Y.Y. Effect of Dietary Fibre and Grit on Performance, Gastrointestinal Tract Development, and Grit Pattern of Goose. Br. Poult. Sci. 2020, 61, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Liu, Y. A Reasonable Diet Promotes Balance of Intestinal Microbiota: Prevention of Precolorectal Cancer. BioMed Res. Int. 2019, 2019, 3405278. [Google Scholar] [CrossRef] [PubMed]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel Disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef] [PubMed]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; de Los Reyes-Gavilán, C.G.; Salazar, N. Intestinal Short Chain Fatty Acids and Their Link with Diet and Human Health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef]
- Duncan, S.H.; Louis, P.; Flint, H.J. Cultivable Bacterial Diversity from the Human Colon. Lett. Appl. Microbiol. 2007, 44, 343–350. [Google Scholar] [CrossRef]
- Nilholm, C.; Manoharan, L.; Roth, B.; D’Amato, M.; Ohlsson, B. A Starch- and Sucrose-Reduced Dietary Intervention in Irritable Bowel Syndrome Patients Produced a Shift in Gut Microbiota Composition Along with Changes in Phylum, Genus, and Amplicon Sequence Variant Abundances, without Affecting the Micro-RNA Levels. United Eur. Gastroenterol. J. 2022, 10, 363–375. [Google Scholar] [CrossRef]
- Ueki, A.; Akasaka, H.; Suzuki, D.; Ueki, K. Paludibacter Propionicigenes Gen. Nov., Sp. Nov., a Novel Strictly Anaerobic, Gram-Negative, Propionate-Producing Bacterium Isolated from Plant Residue in Irrigated Rice-Field Soil in Japan. Int. J. Syst. Evol. Microbiol. 2006, 56, 39–44. [Google Scholar] [CrossRef]
- Malinowska, A.M.; Kok, D.E.; Steegenga, W.T.; Hooiveld, G.; Chmurzynska, A. Human Gut Microbiota Composition and Its Predicted Functional Properties in People with Western and Healthy Dietary Patterns. Eur. J. Nutr. 2022, 61, 3887–3903. [Google Scholar] [CrossRef]
- Hou, Y.; Wu, G. L-Glutamate Nutrition and Metabolism in Swine. Amino Acids 2018, 50, 1497–1510. [Google Scholar] [CrossRef]
- Dong, Z.; Sinha, R.; Richie, J.P., Jr. Disease Prevention and Delayed Aging by Dietary Sulfur Amino Acid Restriction: Translational Implications. Ann. N. Y. Acad. Sci. 2018, 1418, 44–55. [Google Scholar] [CrossRef]
- Costa, K.A.; Soares, A.D.; Wanner, S.P.; Santos, R.; Fernandes, S.O.; Martins Fdos, S.; Nicoli, J.R.; Coimbra, C.C.; Cardoso, V.N. L-Arginine Supplementation Prevents Increases in Intestinal Permeability and Bacterial Translocation in Male Swiss Mice Subjected to Physical Exercise under Environmental Heat Stress. J. Nutr. 2014, 144, 218–223. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, Z.Y.; Yang, H.M.; Zhao, F.Z.; Kong, L.L. Response of Growing Goslings to Dietary Supplementation with Methionine and Betaine. Br. Poult. Sci. 2016, 57, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Zhang, H.; Li, G.; Wu, Z. Nickel-Cobalt Bimetallic Anode Catalysts for Direct Urea Fuel Cell. Sci. Rep. 2014, 4, 5863. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Song, A.A.; Loh, T.C.; Abdul Rahim, R. Effects of Lysine and Methionine in a Low Crude Protein Diet on the Growth Performance and Gene Expression of Immunity Genes in Broilers. Poult. Sci. 2020, 99, 2916–2925. [Google Scholar] [CrossRef] [PubMed]
- Castaner, O.; Goday, A.; Park, Y.M.; Lee, S.H.; Magkos, F.; Shiow, S.T.E.; Schröder, H. The Gut Microbiome Profile in Obesity: A Systematic Review. Int. J. Endocrinol. 2018, 2018, 4095789. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, A.; Sandhu, A.K.; Edirisinghe, I.; Burton-Freeman, B.M. Functional Deficits in Gut Microbiome of Young and Middle-Aged Adults with Prediabetes Apparent in Metabolizing Bioactive (Poly)Phenols. Nutrients 2020, 12, 3595. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Basal Fattening Diet |
|---|---|
| Ingredient, % | |
| Corn grain | 50.10 |
| Sorghum grain | 21.80 |
| Soybean meal | 13.00 |
| Cottonseed meal | 10.50 |
| Limestone | 0.90 |
| Sodium bicarbonate | 1.20 |
| NaCl | 0.40 |
| DL-Methionine | 0.10 |
| 1 Pre-mix | 2.00 |
| Total | 100.00 |
| Nutritional levels | |
| 2 Metabolizable energy/(MJ/kg) | 11.74 |
| Crude protein (%) | 15.25 |
| Crude fiber (%) | 3.69 |
| Calcium (%) | 0.85 |
| Total phosphorus (%) | 0.64 |
| Available phosphorus (%) | 0.36 |
| Lysine (%) | 0.72 |
| Methionine (%) | 0.36 |
| Fermented maize stover | |
| Ingredient, % | |
| Dry matter | 35.97 |
| Organic substance | 93.30 |
| Crude protein | 6.17 |
| Crude fiber | 40.65 |
| Crude ash | 6.70 |
| Neutral detergent fiber | 76.06 |
| Acid detergent fiber | 47.06 |
| Lignin | 7.74 |
| 3 Gross energy/(MJ/kg) | 20.18 |
| Digestible energy/(MJ/kg) | 9.83 |
| Metabolizable energy/(MJ/kg) | 7.94 |
| Fermentation quality parameters | |
| pH | 3.96 |
| Lactic acid (% DM) | 4.41 |
| Acetic acid (% DM) | 1.23 |
| Propionic acid (% DM) | 0.021 |
| Butyric acid (% DM) | 4 ND |
| 5 NH3-N/TN (%) | 3.21 |
| Indicators | A | B | C | D | SEM | p-Value |
|---|---|---|---|---|---|---|
| Initial body weight (g) | 3012.93 ± 35.844 | 3009.06 ± 35.427 | 2998.25 ± 34.802 | 2995.5 ± 35.369 | 17.597 | 0.982 |
| Slaughter body weight (g) | 3505.33 ± 37.734 | 3411.67 ± 36.866 | 3419.13 ± 31.393 | 3472.93 ± 33.101 | 17.492 | 0.180 |
| Average daily gain (g) | 23.45 ± 0.225 A | 19.17 ± 0.308 C | 20.04 ± 0.335 B | 22.73 ± 0.345 A | 0.184 | <0.001 |
| Average daily feed intake (g) | 208.99 ± 0.396 | 208.26 ± 0.559 | 208.2 ± 0.172 | 208.57 ± 0.399 | 0.197 | 0.525 |
| Feed/gain (g/g) | 8.97 ± 0.078 C | 11.08 ± 0.188 A | 10.58 ± 0.159 B | 9.33 ± 0.141 C | 0.089 | <0.001 |
| Dressed yield (%) | 87.39 ± 0.199 ab | 87.31 ± 0.139 ab | 87.09 ± 0.162 b | 87.64 ± 0.178 a | 0.086 | 0.159 |
| Half-eviscerated yield (%) | 78.66 ± 0.220 | 77.85 ± 0.429 | 78.25 ± 0.215 | 78.52 ± 0.297 | 0.152 | 0.247 |
| Eviscerated yield (%) | 71.19 ± 0.192 a | 70.19 ± 0.420 b | 70.69 ± 0.309 ab | 71.29 ± 0.287 a | 0.157 | 0.051 |
| Breast muscle yield (%) | 17.64 ± 0.161 | 17.16 ± 0.195 | 17.53 ± 0.154 | 17.66 ± 0.159 | 0.084 | 0.121 |
| Leg muscle yield (%) | 12.61 ± 0.132 | 12.4 ± 0.138 | 12.39 ± 0.109 | 12.55 ± 0.111 | 0.062 | 0.511 |
| Lean meat yield (%) | 30.24 ± 0.273 | 29.55 ± 0.321 | 29.92 ± 0.254 | 30.21 ± 0.252 | 0.138 | 0.257 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, X.; Zhang, Y.; Ni, H.; Xiao, Q.; Yin, Y.; Ren, J.; Zhao, P.; Zhang, Z.; Li, X.; Li, Y.; et al. Optimization of Fermented Maize Stover for the Fattening Phase of Geese: Effect on Production Performance and Gut Microflora. Animals 2024, 14, 433. https://doi.org/10.3390/ani14030433
Hong X, Zhang Y, Ni H, Xiao Q, Yin Y, Ren J, Zhao P, Zhang Z, Li X, Li Y, et al. Optimization of Fermented Maize Stover for the Fattening Phase of Geese: Effect on Production Performance and Gut Microflora. Animals. 2024; 14(3):433. https://doi.org/10.3390/ani14030433
Chicago/Turabian StyleHong, Xiaoqing, Yonghong Zhang, Hongyu Ni, Qingxing Xiao, Yijing Yin, Jing Ren, Puze Zhao, Ziyi Zhang, Xiaohui Li, Yumei Li, and et al. 2024. "Optimization of Fermented Maize Stover for the Fattening Phase of Geese: Effect on Production Performance and Gut Microflora" Animals 14, no. 3: 433. https://doi.org/10.3390/ani14030433
APA StyleHong, X., Zhang, Y., Ni, H., Xiao, Q., Yin, Y., Ren, J., Zhao, P., Zhang, Z., Li, X., Li, Y., & Yang, Y. (2024). Optimization of Fermented Maize Stover for the Fattening Phase of Geese: Effect on Production Performance and Gut Microflora. Animals, 14(3), 433. https://doi.org/10.3390/ani14030433