1. Introduction
In modern society dogs (
Canis lupus familiaris) have to perform a wide range of roles including assistance [
1], detection [
2], herding and guarding livestock [
3] and competitive sports [
4], as well as companionship as pets [
5]. The trainability and cognitive performance of these dogs in each of their different roles is highly dependent on a number of factors that include genetics (breed), level of training, motivation, arousal, inhibitory control, persistence, human dependence, and independence in problem solving, and these factors may also interact with one another in different ways [
6,
7,
8,
9]. Breed can influence how dogs interact with humans; for example, pastoral breeds show a stronger preference for looking to their owner for help than do gundog breeds [
10,
11], and working dog breeds are better at understanding human gestures than non-working breeds [
12], but experience can overcome breed differences [
13]. This indicates that training and experience may be just as important as genetics in determining the cognitive performance of dogs [
14].
Studies on the impact of training background (discipline and training level) on problem solving ability have shown differences between dogs trained to a high level in working or sporting roles and pet dogs that have received little or no training. For example, dogs that are highly trained for work or sporting activities outperformed dogs with only basic obedience training in a box opening task [
15], as well as in both a detour and a puzzle box task [
16]. Search and rescue dogs outperformed pet dogs with basic obedience training in an A-not-B task, showing higher levels of inhibitory control [
6], and improved performance in a puzzle box task [
17]. Trained assistance dogs outperform pet dogs in a detour task [
7] and outperform pet dogs with recreational sport training (primarily obedience and agility) or no training on a puzzle task [
18], although in the latter study, dogs trained in a recreational sport also outperform those with no training.
Inhibitory control has been shown to have a significant effect on canine problem solving ability [
19]. This describes the ability to inhibit an impulsive action, immediately favourable but undesired or inappropriate, in favour of an alternative action, typically more desired and appropriate for the situation [
6,
20]. Highly trained dogs demonstrate sustained attention to stimuli [
21] and enhanced levels of inhibitory control compared to untrained dogs [
6,
7,
16]. Inhibitory control involves complex cognitive processes and may be influenced by motivation and arousal, and good inhibitory control is beneficial in tasks requiring dogs to ignore stimuli in their environment or work in high arousal environments.
Highly trained dogs exhibit more motivation and persistence in problem-solving tasks than do untrained dogs [
16,
17]. Training can also alter the level of dependence on cues from the handler, as working dogs appear to be less socially dependent on their owners [
18,
22]. These studies primarily focus on training level, rather than on the impact of specific discipline; however, training disciplines that require different cognitive skills are predicted to impact on the development of executive function [
18,
23], which may in turn affect problem solving ability.
Independence is also a critical factor in determining trainability and cognitive performance. Scent detection dogs, for example, are required to act independently when performing scent detection work [
24] and also have to exhibit good task focus, requiring some level of inhibitory control to avoid distraction from a search [
25,
26]. Agility dogs are required to follow their handler’s cues and signals throughout agility training and performance and demonstrate a higher level of handler gaze during problem solving compared to untrained pet dogs [
27,
28]. Obedience dogs show even less independent thinking while performing their specific discipline, as they are required to closely follow each command given by their handler, although these dogs must also display a good level of inhibitory control to maintain focus on their handler [
14,
29].
Additional factors that also require consideration in determining the trainability and cognitive ability of dogs include sex, age, temperament, and arousal. In a study involving 483 males and 549 females, female dogs achieved a higher success rate in the cylinder task than male dogs, indicating that females have greater inhibitory control [
30]. Age has been found to influence inhibitory control, which decreases in older dogs, consequently exerting negative effects on cognitive ability [
20] and cognitive flexibility [
31]. Temperament can also affect the reliance of dogs on the owner vs. an unfamiliar demonstrator [
9]; dogs with ‘irritable’ behaviours, as reported by owners, gazed less towards their owner during a detour task, but performed better when the task was first demonstrated by an unfamiliar person. Increased arousal can enhance performance, but if an animal is already in a higher arousal state, further arousal is likely to result in poorer cognitive performance [
32]. In pet dogs, higher levels of arousal typically have negative impacts on performance when compared with the performance of highly trained working dogs [
7].
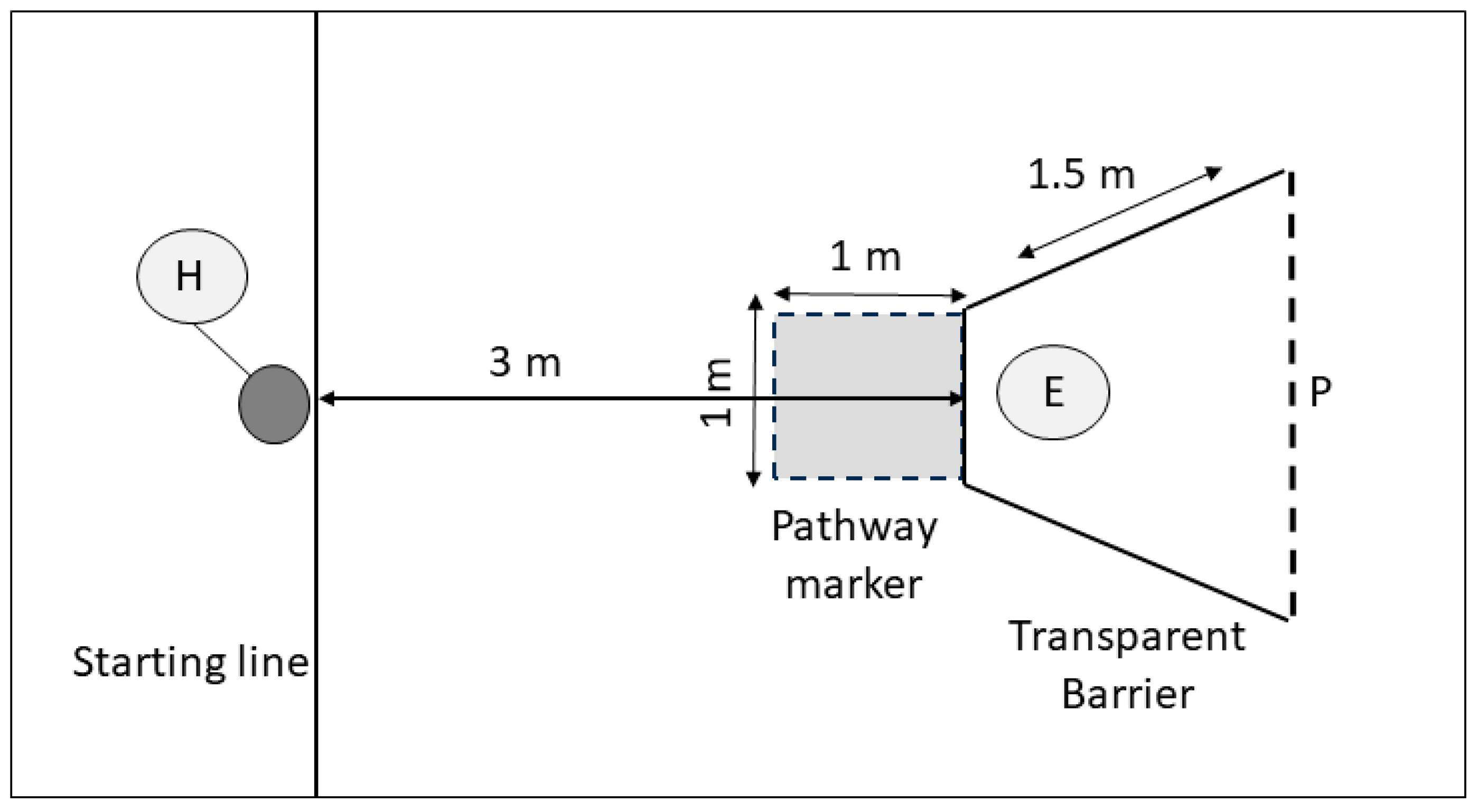
Considering these primary factors affecting trainability and cognitive performance, the aim of this study was to assess the effect of training discipline (obedience, scent work, and agility) and training level on problem solving in pet dogs, using two specific cognitive tasks (the A-not-B task and the detour task) that are heavily reliant on inhibitory control for their successful completion [
33,
34]. The A-not-B task determines whether the dog is able to inhibit search behaviour towards a previously rewarded location after watching the experimenter move the reward to a new location [
20,
35,
36]. In the detour task, the dog must move around a transparent barrier to access a visible reward, requiring the dog to inhibit its direct approach response to the reward [
7,
34]. We therefore predicted
a priori that scent work dogs, who are required to work independently and who require inhibitory control to remain task focused during searches, would outperform dogs primarily trained in the other disciplines, and that they would demonstrate enhanced performance in both the A-not-B and detour tasks. In addition, we also assessed dogs for stress indicators and eye contact with their handler as measures of anxiety, arousal level, human dependence, and attachment [
7,
37,
38]. We predicted that dogs showing a high number of stress indicators and increased eye contact with their handler would demonstrate poorer cognitive performance due to high arousal and lack of handler independence.
3. Results
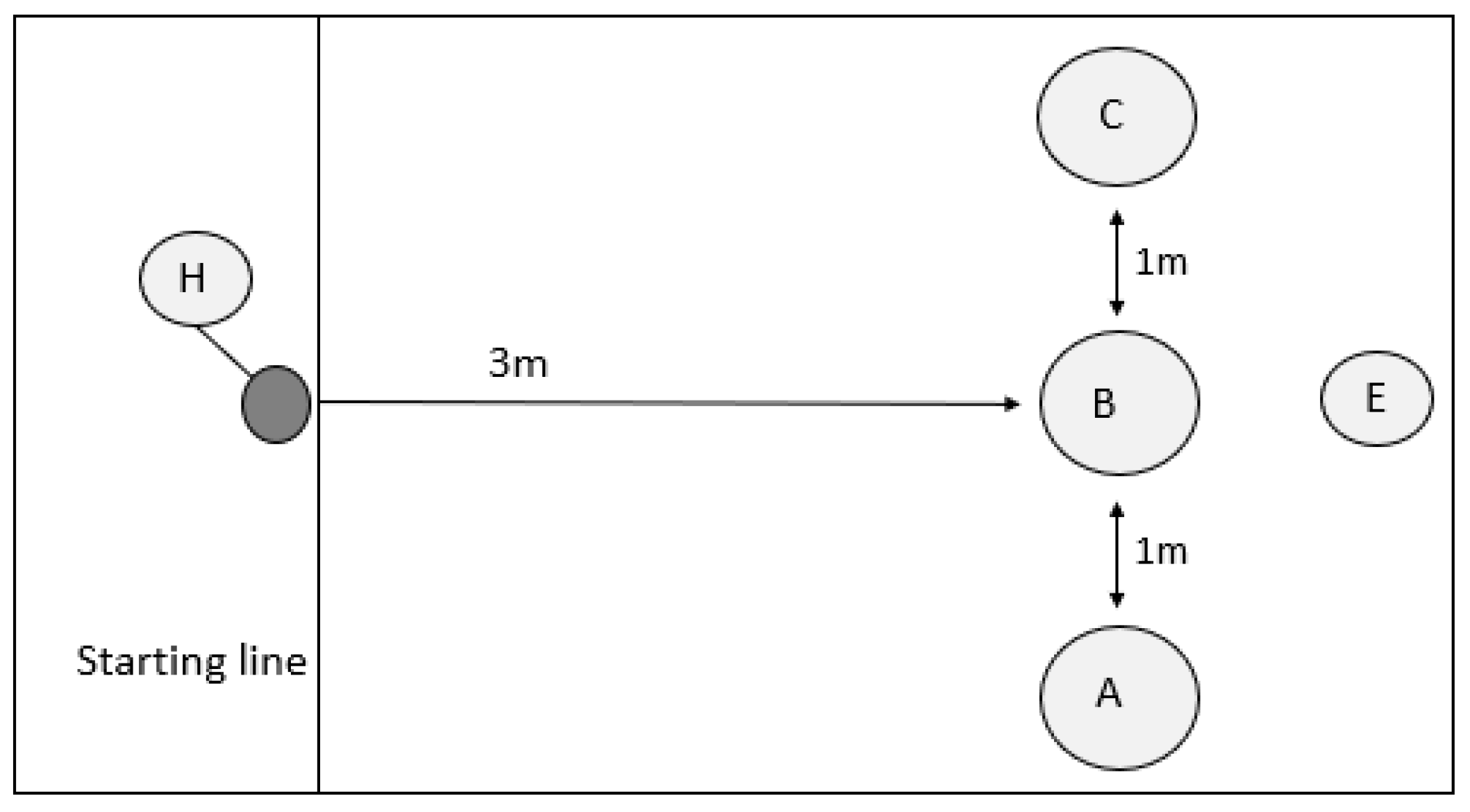
3.1. A-Not-B Task
Dogs were considered successful in the A-not-B task if they selected the ‘B’ pot at least 8/10 times during the test phase. We also separated A-not-B dogs further, splitting them into those that failed the training phase (did not select A correctly five times in a row: FAIL_TRAIN; N = 4), those that passed the training phase, but did not pass the test phase criteria (PASS_TRAIN_FAIL_TEST; N = 15), and those that passed both training and test phases (PASS_TRAIN_PASS_TEST; N = 18). See
Supplementary Tables S2 and S3 for the A-not-B data used in the analyses.
Primary Training: The primary training discipline had no significant effect on success in meeting the test criteria in the A-not-B task (χ2 = 0.960, d.f. = 3, p = 0.811), and there was also no significant effect of the primary training discipline on the number of perseverative errors in the A-not-B task (ANOVA: F3,29 = 0.497, p = 0.688). There was no effect of the primary training discipline (rmANOVA: F3,27 = 1.171, p = 0.339), trial (rmANOVA: F3.094,83.539 = 1.410, p = 0.245), or interaction between these factors (rmANOVA: F9.282,83.593 = 0.973, p = 0.470) on the time to select a pot in the A-not-B task.
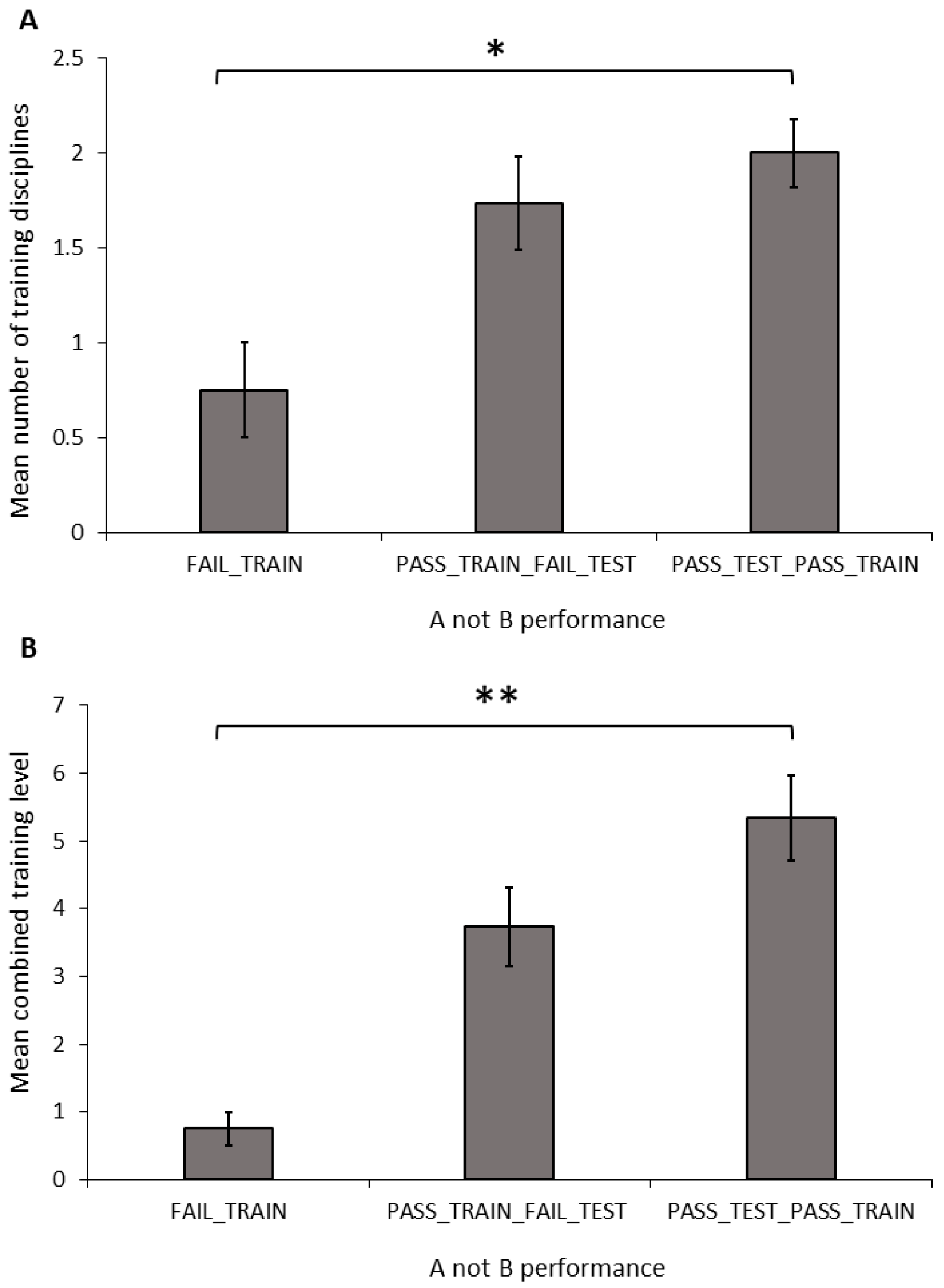
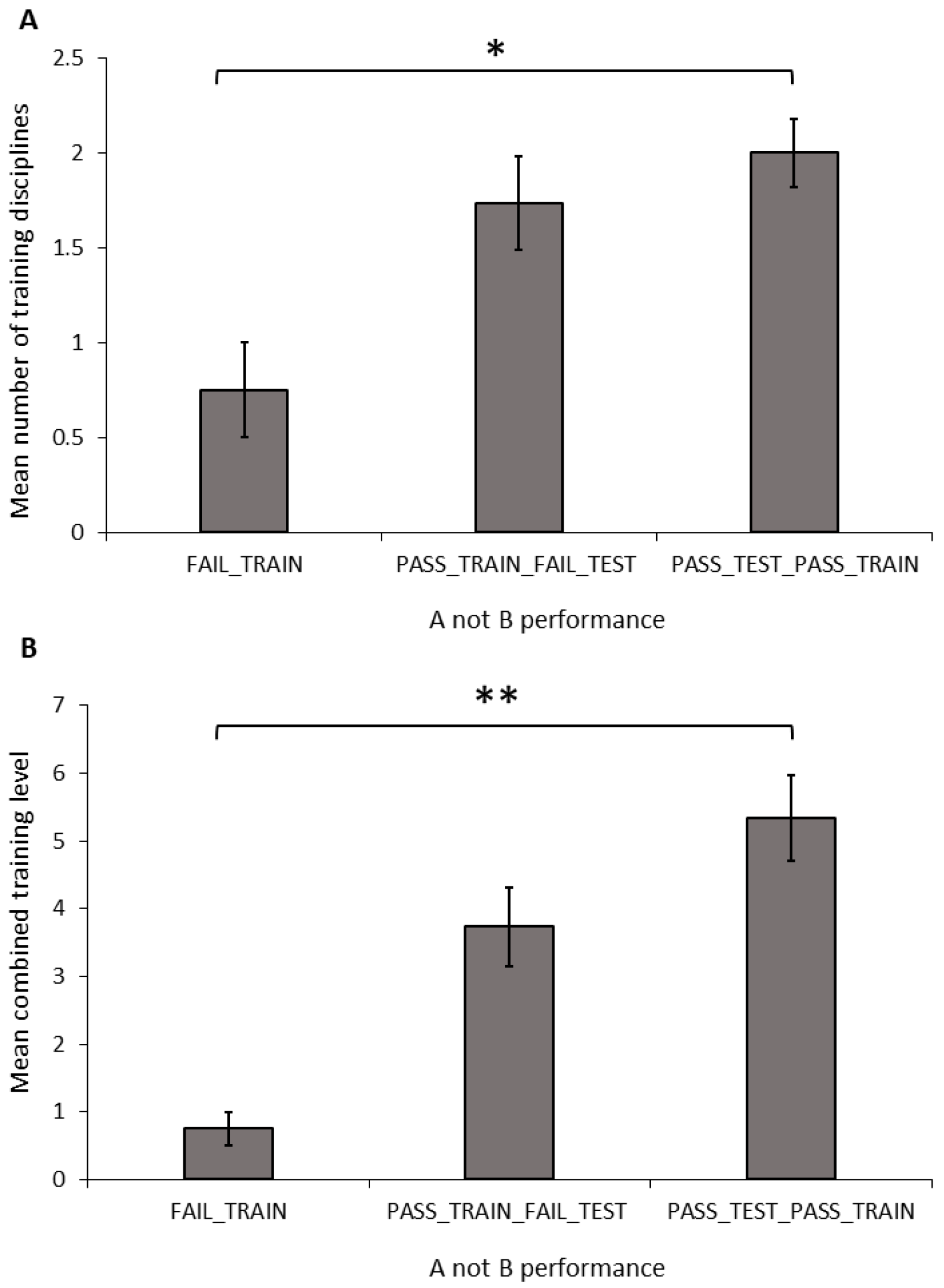
Number of disciplines: The number of training disciplines a dog participated in differed significantly between the performance groups in the A-not-B task (
Figure 3A; ANOVA: F
2,34 = 3.681,
p = 0.036). The FAIL_TRAIN group took part in significantly fewer disciplines than did the PASS_TRAIN_PASS_TEST group (Sidak:
p = 0.031); however, there was no difference between dogs in the FAIL_TRAIN and PASS_TRAIN_FAIL_TEST groups (Sidak:
p = 0.126) or the PASS_TRAIN_FAIL_TEST and PASS_TRAIN_PASS_TEST groups (Sidak:
p = 0.747). The number of disciplines a dog trained in showed no significant relationship with the number of perseverative errors during the A-not-B task (Pearson’s correlation: r = −0.035, N = 33,
p = 0.846).
Combined training experience: The combined training level differed significantly between the A-not-B groups (
Figure 3B; ANOVA: F
2,34 = 6.428,
p = 0.004). The FAIL_TRAIN group had a significantly lower combined training experience score compared to that of the PASS_TRAIN_PASS_TEST group (Sidak:
p = 0.004); whereas the FAIL_TRAIN group did not differ significantly from the PASS_TRAIN_FAIL_TEST group (Sidak:
p = 0.098); and there was also no significant difference between the PASS_TRAIN_FAIL_TEST group and the PASS_TRAIN_PASS_TEST group (Sidak:
p = 0.182). The combined training experience showed no significant relationship with the number of perseverative errors during the A-not-B task (Pearson’s correlation: r = −0.227, N = 33,
p = 0.204).
Success, speed, and error rate: Only dogs that completed all ten A-not-B test trials were included in the analyses. Dogs that passed the A-not-B task required fewer trials to select the correct ‘B’ pot (mean ± sem: 1.22 ± 0.10) than dogs that failed the A-not-B task (2.33 ± 0.46; GLM: χ2 = 8.392, d.f. = 1, p = 0.004). First trial selection predicted whether dogs would go on to meet trial criteria in the A-not-B task (χ2 = 4.591, d.f. = 1, p = 0.032), with 14/18 dogs that succeeded in reaching the test criteria (8/10) making the correct choice on the first trial, and only 5/13 dogs that failed making the correct choice on the first trial meeting the test criteria.
Dogs did not alter their speed of pot choice significantly across trials in the A-not-B task (rmANOVA: F2.973,86.217 = 1.972, p = 0.125), nor was there an interaction between success in the task and the speed of choice across the trials (rmANOVA: F2.973,86.217 = 1.729, p = 0.167); however, dogs that met the criteria (8/10 correct) in the A-not-B task were significantly faster at choosing a pot than those that failed to reach this criteria (rmANOVA: F1,29 = 13.949, p = 0.001). Dogs that were slower, on average, to choose a pot also made a greater number of perseverative errors overall (Pearson’s correlation: r = 0.528, N = 31, p = 0.002).
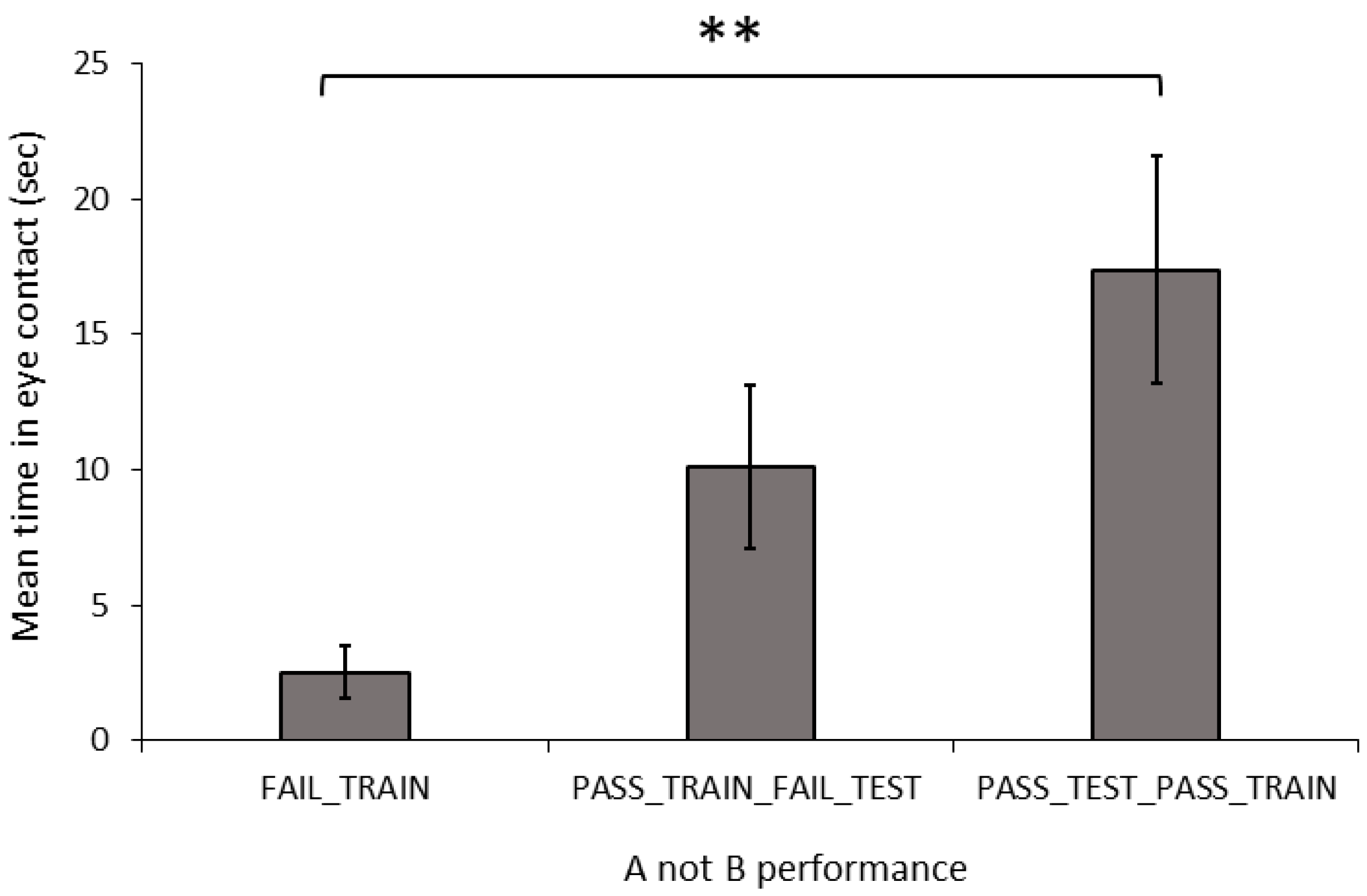
Behaviour and Dog Characteristics: In the A-not-B, task there was a significant difference in the duration of eye contact with the owner between the training-testing groups (
Figure 4; Welch: F
2,20.467 = 7.889,
p = 0.003). The FAIL_TRAIN dogs made significantly less eye contact than did the PASS_TRAIN_PASS_TEST dogs (G-H:
p = 0.009), whereas the PASS_TRAIN_FAIL_TEST dogs did not differ in the level of eye contact compared to those of the other two groups (FAIL_TRAIN: G-H:
p = 0.074; PASS_TEST_PASS_TRAIN; G-H:
p = 0.352). The number of stress indicators did not differ significantly between the three groups (ANOVA: F
2,31 = 0.067,
p = 0.936).
There was no significant relationship between the duration of eye contact with the owner (Pearson’s correlation: r = −0.287, N = 30, p = 0.124) or the number of stress indicators (Pearson’s correlation: r = 0.143, N = 30, p = 0.452) with the number of perseverative errors across 10 trials in the test for the A-not-B task.
There was no effect of age group (χ2 = 0.027, d.f. = 1, p = 0.869), sex (χ2 = 0.068, d.f. = 1, p = 0.794), neuter status (χ2 = 0.160, d.f. = 1, p = 0.689), or interaction between neuter status and sex (χ2 = 0.002, d.f. = 1, p = 0.967), on success in the A-not-B task. Breed category (gundog, pastoral, other) did not significantly affect success in the A-not-B task (χ2 = 3.282, d.f. = 2, p = 0.194).
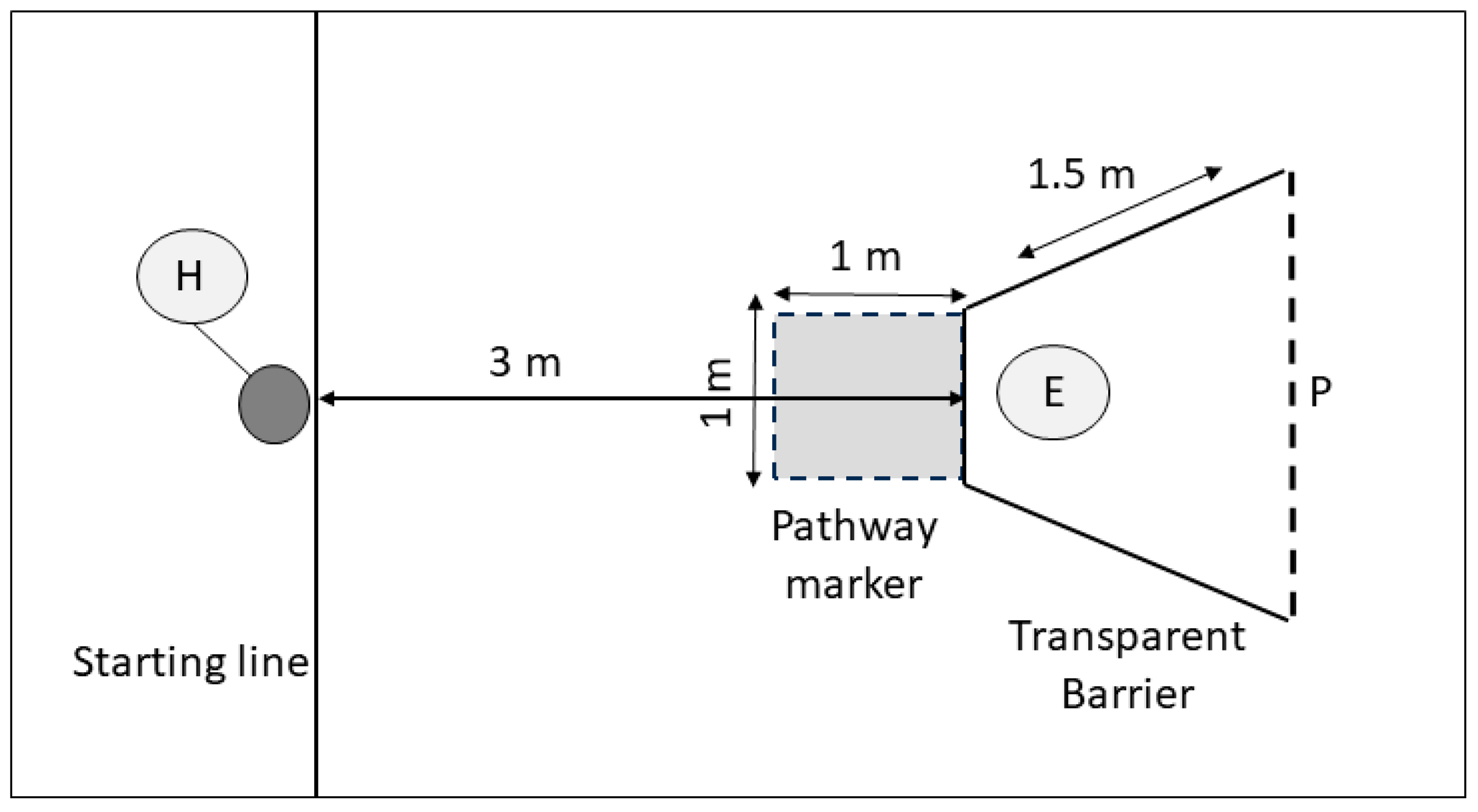
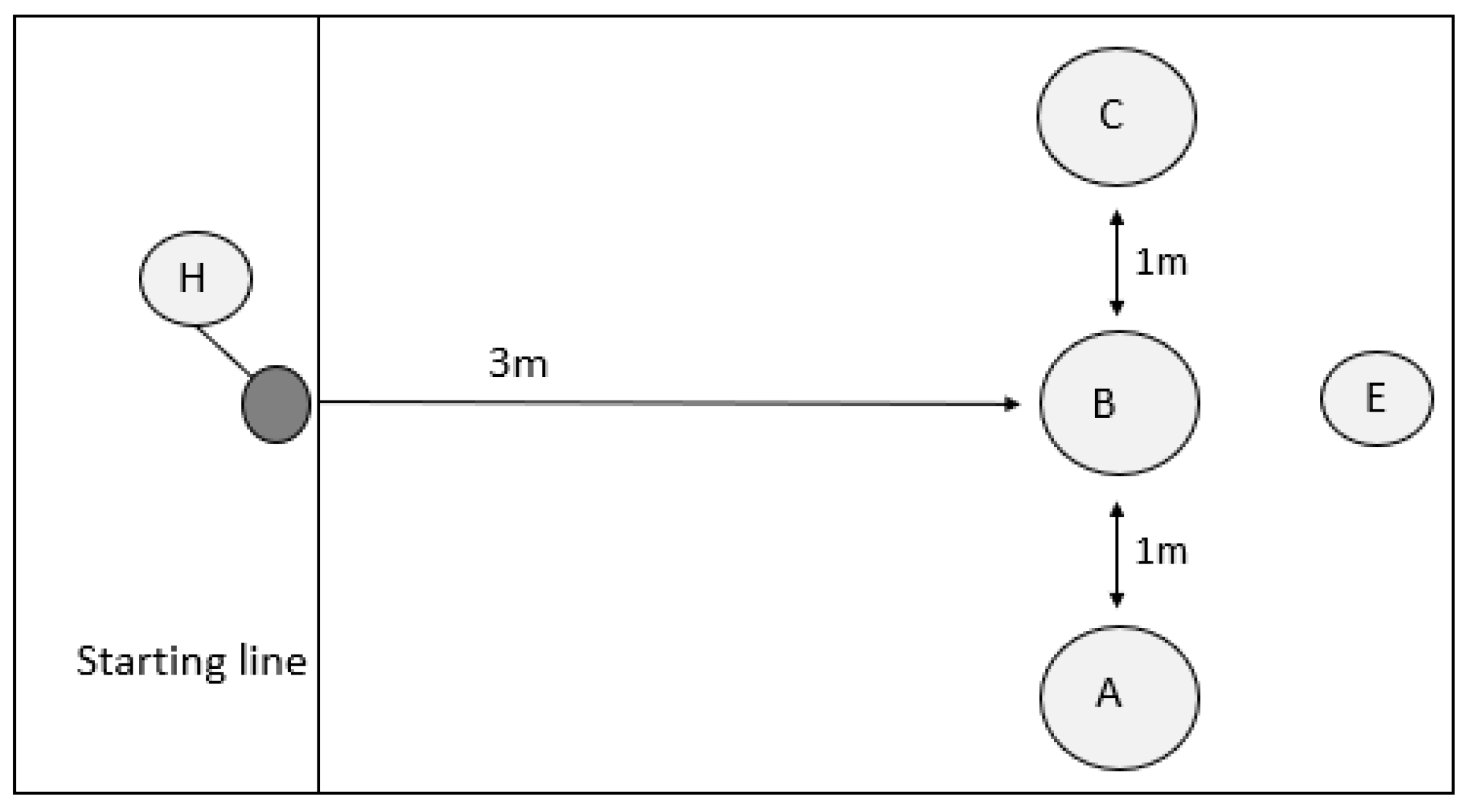
3.2. Detour Task
Dogs were considered successful in the detour task if they moved around the barrier in 8/10 trials, without contacting the barrier (contact error) or entering the direct approach zone (path error). A total of 14 dogs met the criteria for the detour task (pass), and 24 did not reach this criteria (fail). See
Supplementary Tables S4 and S5 for the detour data used in the analyses.
Primary Training Discipline: The primary training discipline did not affect the success rate in the detour task (χ2 = 2.934, d.f. = 3, p = 0.402). Moreover, the primary training discipline did not impact on the contact error rate (Welch: F3,11.856 = 1.860, p = 0.191) or the path error rate (ANOVA: F3,34 = 0.817, p = 0.493), but did significantly affect the total error rate (Welch: F3,17.247 = 3.277, p = 0.046) in the detour task. However, the results of the Games–Howell post hoc pairwise tests were not significant for any pair-wise comparisons (G-H: p > 0.05).
Number of disciplines: The number of disciplines a dog trained in did not differ between dogs that succeeded or failed in the detour task (
Figure 5A; ANOVA: F
1,36 = 0.246,
p = 0.623). There was also no relationship between the number of training disciplines and the number of contact errors (Pearson’s correlation: r = 0.166, N = 38,
p = 0.319), the number of path errors (Pearson’s correlation: r = 0.017, N = 38,
p = 0.918), or the total number of errors (Pearson’s correlation: r = 0.094, N = 38,
p = 0.575) made during the detour task.
Combined training experience: The combined training level across disciplines did not differ between dogs that failed or succeeded in the detour task (
Figure 5B; ANOVA: F
1,36 = 0.249,
p = 0.621). There was also no relationship between the combined training experience and the number of path errors (Pearson’s correlation: r = −0.083, N = 38,
p = 0.918) or the total number of errors (Pearson’s correlation: r = 0.142, N = 38,
p = 0.394) made in the detour task. However, dogs with a greater combined training experience were more likely to contact the barrier in the detour task (Pearson’s correlation: r = 0.422, N = 38,
p = 0.008).
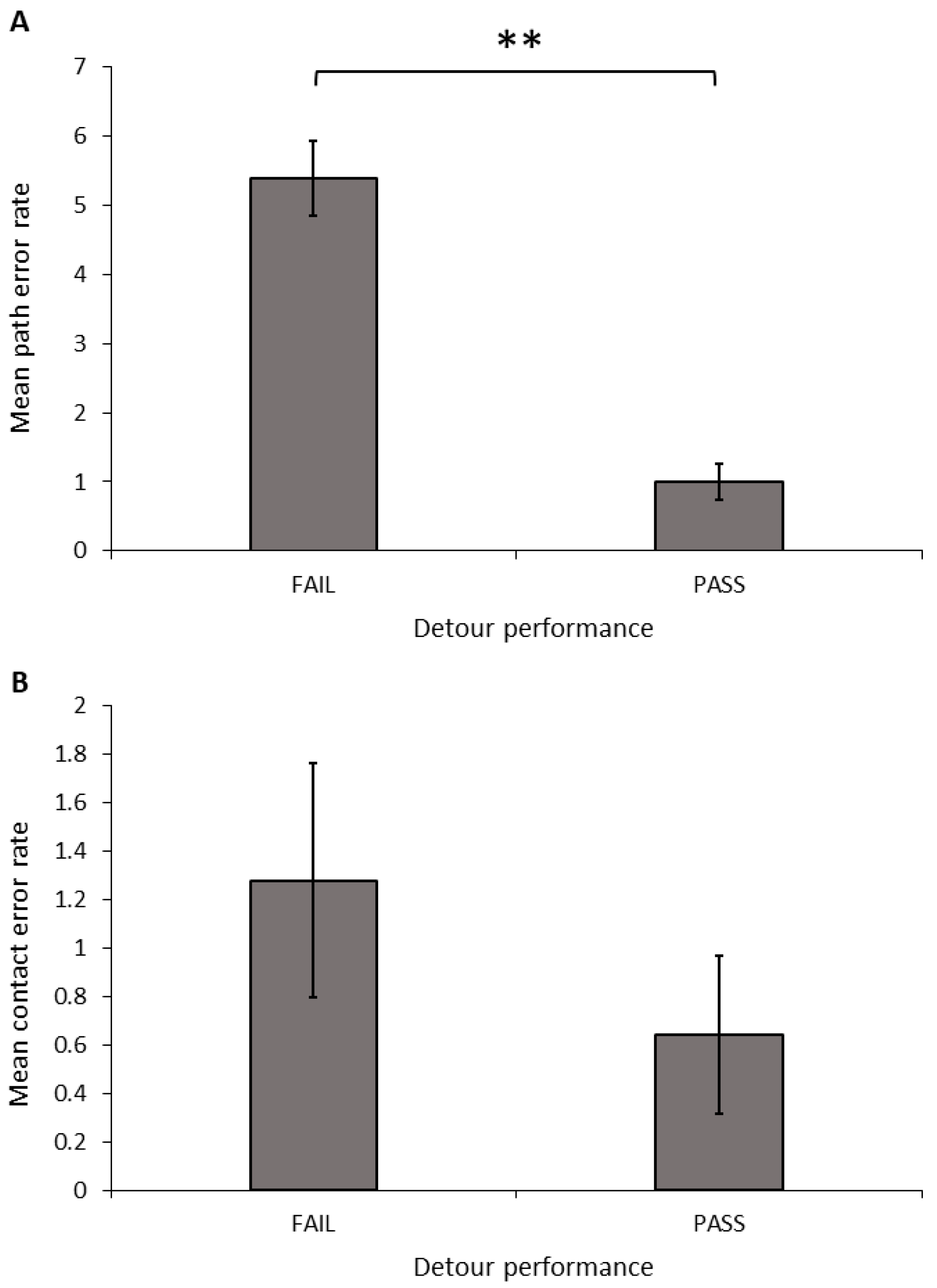
Success, speed, and error rate: Only dogs that completed all ten detour test trials were included in the analyses. Dogs that succeeded in meeting the criteria in the detour task made significantly fewer path errors (
Figure 6A;
t-test equal variances not assumed:
t = 6.670, d.f. = 23.887,
p < 0.001) and total errors (
t-test equal variances not assumed:
t = 5.568, d.f. = 24.447,
p < 0.001) than dogs that failed to reach these criteria; however, there was no significant difference in the number of contact errors (
Figure 6B;
t-test:
t = 1.024, d.f. = 30,
p = 0.314). We also found a significant relationship between the successful detour rate and both the path error rate (Spearman’s correlation: r = −0.997, N = 32,
p < 0.001) and the total error rate (Spearman’s correlation: r = −0.943, N = 32,
p < 0.001). Dogs with a higher detour success rate had a lower number of path errors and total errors, but there was no relationship between the success rate and the contact error rate (Spearman’s correlation: r = 0.204, N = 32,
p = 0.263).
The dogs did not alter their detour speed significantly across the trials (rmANOVA: F3.609,108.283 = 1.545, p = 0.132), nor was there an interaction between success in the task and the speed of detour (rmANOVA: F3.609,108.283 = 0.959, p = 0.474); however, dogs that met the criteria (8/10 correct) in the detour task were significantly faster (3.143 s, CI: 1.477, 4.810) in moving around the barrier than those that failed to reach the criteria (7.442 s, CI 5.973, 8.912; rmANOVA: F1,30 = 15.615, p < 0.001).
The dogs that took more time on average to complete the detour task, on average, made a greater number of path errors (Pearson’s correlation: r = 0.518, N = 32, p = 0.002) and total errors (Pearson’s correlation: r = 0.370, N = 32, p = 0.037), but there was no relationship between the average detour time and the contact errors (Pearson’s correlation: r = −0.062, N = 32, p = 0.738).
Behaviour and dog characteristics: There was no significant difference in the number of stress indicators (ANOVA: F1,33 = 1.204, p = 0.281) or time spent watching the owner (ANOVA: F1,33 = 1.810, p = 0.188) between the dogs that passed or failed the detour task.
We found no significant relationship between error rates and the duration of eye contact with the owner (Pearson’s correlation: contact error rate: r = 0.091, N = 35, p = 0.603; path error rate: r = −0.232, N = 35, p = 0.181; combined error rate: r = −0.164, N = 35, p = 0.346). There was a significant relationship between the number of stress indicators and the number of path errors that dogs made during the detour task (Pearson’s correlation: r = −0.336, N = 35, p = 0.048); dogs that exhibited more stress indicator signals made fewer path errors across the 10 detour trials. However, the contact error rate (Pearson’s correlation: r = −0.044, N = 35, p = 0.603) and the total error rate (Pearson’s correlation: r = −0.164, N = 35, p = 0.346) showed no relationship with the number of stress indicators.
The age group did have a significant effect on success in the detour task (χ2 = 4.284, d.f. = 1, p = 0.038), with younger dogs performing better than older dogs, but there was no effect of sex (χ2 = 0.803, d.f. = 1, p = 0.370), neuter status (χ2 = 1.672, d.f. = 1, p = 0.196), or interaction between neuter status and sex (χ2 = 0.827, d.f. = 1, p = 0.363) on success in the detour task. Breed category had no effect on the success rates in the detour task (χ2 = 1.433, d.f. = 2, p = 0.488).
3.3. Comparison of Performance in A-Not-B and Detour Tasks
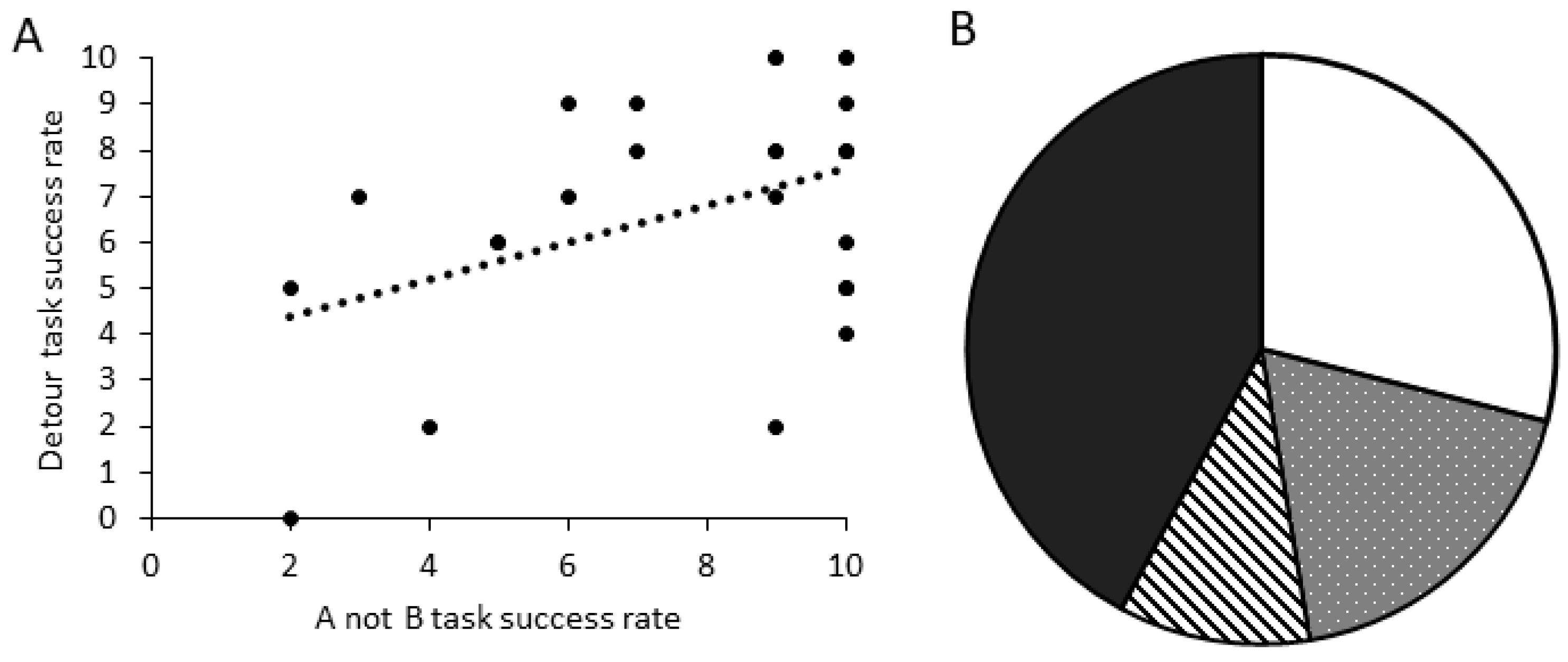
There was a significant positive relationship between the number of correct trials in the A-not-B task and the detour task for dogs that completed all 10 trials in both tasks (
Figure 7A; GLM score comparison: χ
2 = 5.194, d.f. = 1,
p = 0.023). When comparing criteria performance, dogs that were successful in the A-not-B task (PASS_TRAIN_PASS_TEST) were significantly more likely to also be successful in the detour task (
Figure 7B; GLM binomial pass/fail: χ
2 = 5.355, d.f. = 1,
p = 0.021).
There was no significant relationship between the number of perseverative errors dogs made in the A-not-B task and errors made during the detour task in regards to the contact error rate (Pearson’s correlation: r = −0.147, N = 33, p = 0.414), the total error rate (Pearson’s correlation: r = 0.155, N = 33, p = 0.388), or the path error rate (Pearson’s correlation: r = 0.325, N = 33, p = 0.065).
3.4. Principal Components Analysis
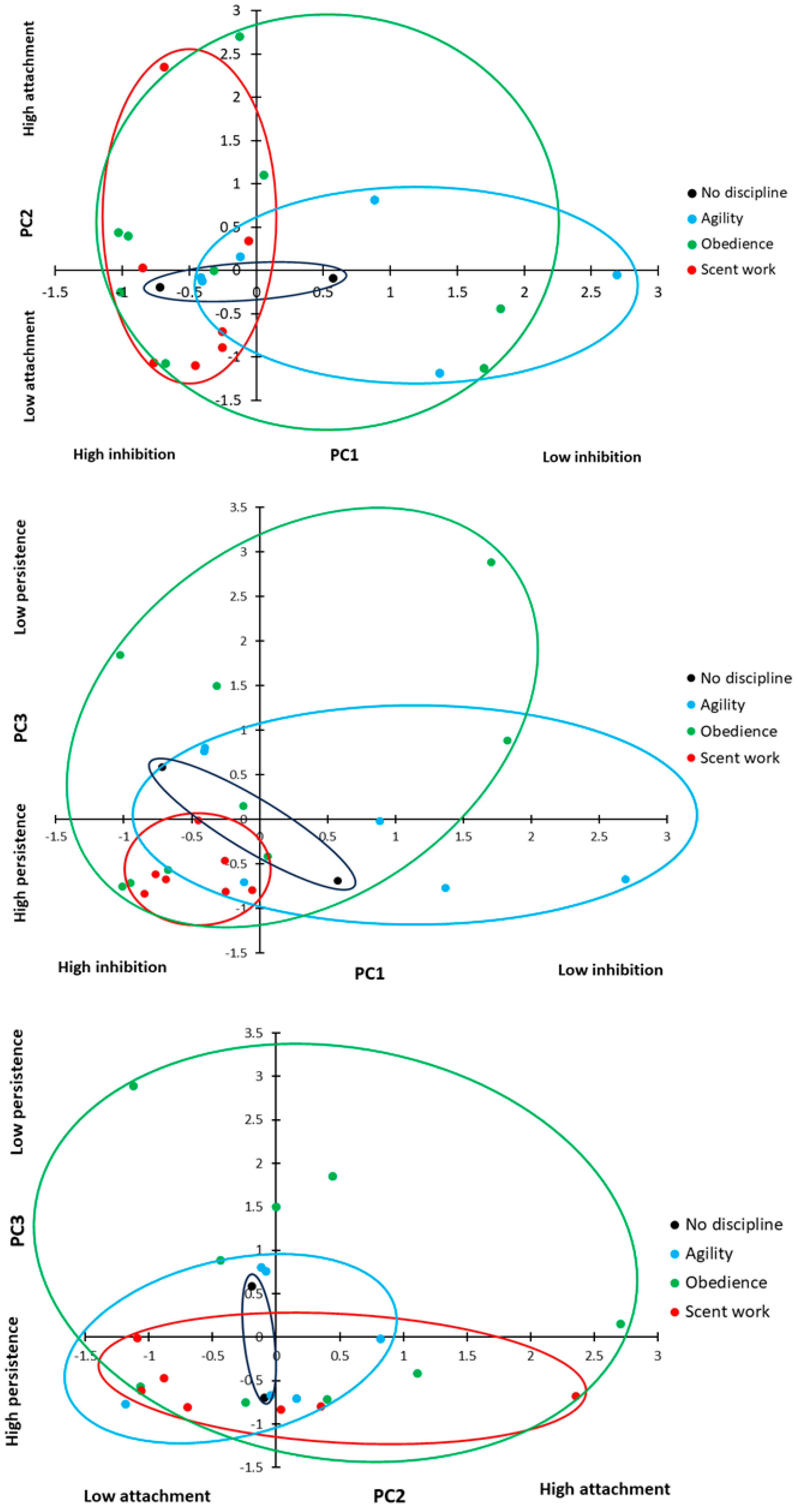
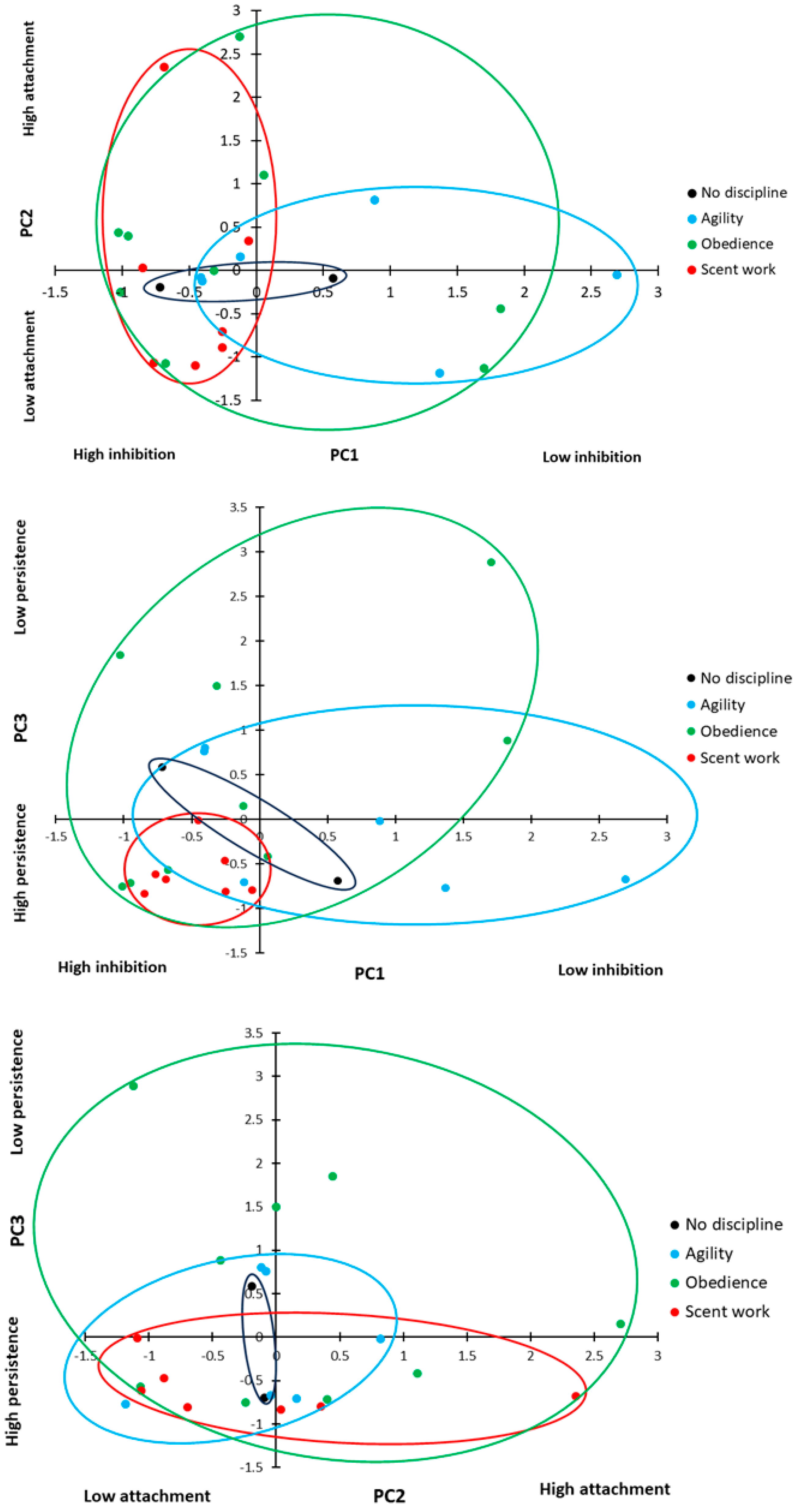
Overall, the PCA revealed three underlying behavioural components, which explained 72.3% of the cumulative variation in behaviour (
Table 3). PC1 included speed and path errors in the detour task, as well as time to select a pot and perseverative errors in the A-not-B task. A high PC1 score represents dogs that took longer to perform both tasks and committed a greater number of errors in both tasks. We considered this to be a good measure of inhibitory control, those dogs with a high score in PC1 showed low inhibitory control. PC2 included stress indicators score and time spent in eye contact with the owner. A high PC2 score covaried with an increased number of stress indicators and increased duration of eye contact, suggesting that this component was related to attachment and arousal behaviour, with anxious dogs seeking owner reassurance in uncertainty. PC3 included contact errors only; a high PC3 score represented dogs that made a high number of contact errors. Contact errors were not related to task success or other errors (see above), but from observation, appeared to represent the dogs attempting an alternative solution to the task; therefore, we consider this to be a measure of persistence. Dogs with a low PC3 score are more likely to persist in repeating the previously rewarded behaviour, whereas dogs with a high PC3 score are more likely to try alternative ways to access the reward.
There was no difference in inhibitory control (PC1: inhibition) scores between dogs based on the discipline number (ANOVA: F
2,17 = 0.514,
p = 0.607) or a relationship between combined training experience and inhibitory control (ANOVA: F
1,17 = 2.075,
p = 0.136). However, there was a significant difference in inhibitory control based on the primary training discipline (
Figure 8; ANOVA: F
2,17 = 3.467,
p = 0.035). Dogs trained primarily in scent work showed significantly higher levels of inhibitory control compared to dogs trained primarily in agility (
Figure 8; Sidak:
p = 0.012), but there was no difference between the other primary disciplines (Sidak:
p > 0.05 for all comparisons). There was no significant effect of sex, neuter status, breed group, or age group on inhibitory control (ANOVA: sex: F
1,18 = 0.603,
p = 0.447; neuter status: F
1,18 = 0.487,
p = 0.494; breed group: F
2,18 = 0.732,
p = 0.495; age group: F
1,18 = 2.001,
p = 0.174).
Attachment and arousal behaviour (PC2: attachment) scores showed no difference between the primary training disciplines (
Figure 8; ANOVA: F
2,17 = 0.754,
p = 0.485) or the number of disciplines a dog trained in (ANOVA: F
2,17 = 0.220,
p = 0.805), and there was no relationship with overall training experience (ANOVA: F
1,17 = 2.447,
p = 0.136). There was no significant effect of sex, neuter status, breed group, or age group on attachment (ANOVA: sex: F
1,18 = 0.054,
p = 0.819; neuter status: F
1,18 = 0.940,
p = 0.345; breed group: F
2,18 = 0.275,
p = 0.763; age group: F
1,18 = 0.787,
p = 0.387).
There was a significant difference in persistence (PC3: persistence) scores between dogs with different primary disciplines (
Figure 8; GLM: χ
2 = 7.029, d.f. = 2,
p = 0.030); dogs trained in scent work showed higher persistence (a lower contact rate) than dogs trained in obedience (
t = 2.519, d.f. = 8.984,
p = 0.033); however, there was no significant difference between the other training groups (agility vs. scent work:
t = 1.559, d.f. = 6.318,
p = 0.167; agility vs. obedience: t = 1.200, d.f. = 12.780,
p = 0.252). There was no significant effect of sex, neuter status, breed group, or age group on persistence (ANOVA: sex: F
1,18 = 0.447,
p = 0.512; neuter status: F
1,18 = 0.628,
p = 0.438; breed group: F
2,18 = 0.350,
p = 0.710; age group: F
1,18 = 0.057,
p = 0.815).
4. Discussion
Previous studies on the impact of training on dog cognition and inhibitory control have largely partitioned dogs into ‘highly trained’ (work or sports) vs. untrained pet dogs, without considering whether the type of training a dog undertakes influences its cognitive ability [
18,
23]. The main goal of our study was to determine whether training in a specific discipline enhances the cognitive ability of pet dogs in tasks that require inhibitory control (A-not-B and detour task), comparing scent work, agility, and obedience. We predicted
a priori that dogs trained primarily in scent work would outperform dogs trained primarily in agility or obedience due to the requirements for dogs trained in scent work to develop both handler independence [
24] and inhibitory control [
25,
26]. We found no evidence to support this hypothesis in the individual tasks, with primary training discipline failing to predict success or error rate in either the A-not-B task or the detour task. However, the primary training discipline did affect dog behaviour across the tasks. Scent dogs exhibited higher overall inhibitory control compared to agility dogs, as well as greater persistence in repeating the rewarded behaviour in the detour task compared to the behaviour of the obedience trained dogs. These results are similar to those of Carballo et al. [
18], who compared the performance of assistance/support dogs to trained family dogs (primarily obedience and agility trained) or untrained family dogs. Scent work requires dogs to perform one task across multiple contexts (find the scent and indicate or track), whereas in agility and obedience training, dogs must perform multiple different behaviours, thus relying on human direction to know which behaviour is expected of them. Assistance dogs, like scent work dogs, require a high level of independence and persistence, alongside inhibitory control, to avoid distractions and perform successfully [
44]. While the cognitive task was different, involving the use of a puzzle box rather than the A-not-B or detour tasks, the assistance dogs outperformed the trained and untrained family dogs, demonstrating less social dependence [
18]. Scent work training has also been shown to create a positive bias in dogs [
45]; however, we found no difference between scent dogs and dogs trained in other disciplines to support an effect of the training discipline on attachment and arousal levels, indicating that differences in arousal levels between disciplines in our cohort was not the main factor impacting on overall performance.
In line with previous studies [
6], we found that combined training experience affected success in the A-not-B task; dogs that succeeded in the A-not-B (at least 8/10 correct) task had been trained in more disciplines and to a higher combined training level compared to dogs that failed to pass the initial training phase. The lower performance may be due to lack of attention to experimenter; dogs that failed the training stage also made less initial eye contact with their owner, indicating they may have paid less attention to the human-directed indication of the correct pot to select to obtain food during the ‘A’ training trials. Despite previous training experience impacting on the initial training phase, we found no significant effect of training level on either criteria or perseverative search errors in dogs that had passed the training phase and no relationship between training experience and the number of successful trials. Attention to human signals is thought to play an important role in success in the A-not-B task, but previous studies have primarily shown that ostensive human communication (e.g., physical gestures such as pointing or gaze shift) after placing the ‘B’ reward leads to errors rather than a positive effect on performance [
35]. There was no significant relationship between performance in the test element for dogs that passed the training stage and prior training experience, contrary to the previous study, which found that highly trained water rescue dogs made fewer perseverative errors during the ‘B’ test phase compared to untrained dogs [
6]. Our sample size (N = 37) was lower than that used in this previous study (N = 48), perhaps reducing our ability to detect an effect of training. In addition, in our study, combined training experience was considered on a linear scale, rather than partitioning dogs between highly trained and untrained, covering a wider range of experience.
Irrespective of training experience, dogs that took longer to make a pot selection in the A-not-B task during the test phase were also more likely to make perseverative errors and fail to meet our criteria. The task may have been more challenging for dogs that have greater reliance on human direction; those taking longer to select a pot may have been waiting for their handler to provide the correct indication [
27]. However, our data showed that dogs who make more eye contact with their handler performed better on the A-not-B task, indicating that this is unlikely. The relationship between latency and performance may instead have been due to motivation for the reward [
45]. Very high-value rewards can decrease inhibitory control through increased arousal, negatively impacting on cognitive performance that requires inhibition [
46]. A longer latency between release and pot selection may suggest that these dogs were unfocused and unable to inhibit their distracted behaviour, therefore taking longer to initially select a pot. Alternatively, perceived high-value rewards can also increase the speed for obtaining the reward [
47] and the attention to human communication in placement of the reward [
48]. Therefore, relative perceived reward value could either hinder or improve performance. We did not determine how valuable the reward was to the dogs, only that they would perform the action of pot selection to obtain it during the familiarisation trials. Perceived reward value and how this impacts on performance has not been tested in the A-not-B task, but it does impact on performance in the cylinder task [
46] and inhibitory control in social interaction tasks [
20]; therefore, it should be considered in future work.
Previous studies have also found enhanced performance on the detour task in highly trained dogs [
7,
16]; however, we found no evidence to support that increased training experience improves performance in pet dogs. This may have been due to a lack of variation in the arousal level related to training experience among the dogs that took part in the test; arousal level has been found to strongly influence detour task performance [
7]. However, dogs with higher combined training experience were more likely to contact the barrier. Barrier contact rate was not associated with the overall level of performance, either in terms of detour speed or ability to meet our detour task criteria (8/10 direct), nor was it related to the frequency of path errors, which was higher in dogs that failed the detour task. This contrasts with previous research in which barrier contact was a good indicator of detour performance [
7]. Given that highly trained dogs outperform those with less training on detour tasks and also exhibit increased inhibitory control to directly approach the reward [
7,
8], the positive relationship between contact errors and training experience seems to be counterintuitive. From our observations, it appeared that contact errors were more strongly associated with dogs attempting an alternative solution to the detour behaviour to gain access to the reward, rather than simply a lack of inhibition resulting in a direct approach to the food. Highly trained dogs have been shown to demonstrate increased puzzle solving abilities [
15,
17,
18]; therefore, it could be that those dogs with a high barrier contact rate were trying to ‘solve’ the puzzle of how to gain the reward without performing the detour behaviour. A direct comparison of puzzle solving ability and detour behaviour would be required to determine if this might be the case.
While contact error rate did not predict detour success, path error rate, in which the dog initially directly approaches the reward, rather than uses the shortest route to detour the barrier, did relate to success. A higher path error rate was strongly associated with low direct detour success across the ten trials, as found in previous work [
7]. Dogs with lower path error rates also detoured the apparatus faster [
7,
49], demonstrating that inhibitory control of the direct approach to the reward improved speed of performance as well as accuracy. Whilst previous training experience did not alter path error rate or overall success, dogs with a higher number of stress indicators were less likely to make path errors during the detour task. In this task the experimenter holding the food bowl is directly facing the dog, a reduction in path errors may be due to a reluctance in dogs that have higher stress levels to directly approach a stranger [
50]. Alternatively, stress can alter the way rewards are perceived [
51], and higher stress or arousal levels may devalue the reward increasing inhibition of direct approach behaviour in the detour task.
Irrespective of training experience, dogs that succeeded in the A-not-B task were more likely to also succeed in the detour task, potentially as both tasks require inhibitory control [
7,
35]. This result is contrary to previous studies that have found no relationship in performance between the A-not-B task and the cylinder task or detour task [
20,
43,
52,
53] and lack of general support in the literature for correlations in performance across tasks requiring impulse control is confirmed in a recent review [
54]. Whilst the detour task is considered comparable to the cylinder task [
7], the difference in motor control, reaching vs. whole body movement around a barrier, and requirement in the detour task to initially move away from the food may lead to important differences in the executive functions required. In a comparison between wolves and dogs on a detour vs. cylinder task, wolves outperformed dogs in the detour task, whereas dogs were more efficient in the cylinder task [
46] indicating differences in the cognitive function tested using these two tasks. In the only other study to date using both A-not-B and a detour task, Vernouillet et al. [
52] tested the number of trials it took for the dog to successfully choose location ‘B’ in the A-not-B task, but did not continue to test the dog’s performance beyond the first successful choice. We found 8/18 dogs that chose the ‘B’ location correctly the first time during the test trials went on to make perseverative errors in subsequent trials, and 14/18 dogs that selected the ‘B’ pot correctly in the first test trial passed our criteria across the 10 trials. Therefore, while dogs that were successful on the first trial were more likely to reach our test criteria, the use of the first trial only did not always predict success, and this result may fail to capture some of the variation in behaviour among individuals.
An issue raised in the comparison of performance across multiple tasks is the ‘ceiling response’, which limits the range of cognitive performance between dogs and thus greatly restricts the ability to find a significant relationship between performance in different cognitive tasks [
20,
23,
54]. In this respect, our success rate, 14/38 (37%) for the detour task, was comparable to that of 45% obtained in a similar study [
16]. However, for the A-not-B task, our success rate 18/37 (48%) was considerably lower than the rate obtained previously (e.g., >80%) in other studies [
20,
52]. The latter, potentially reflects our higher criteria level to pass the training phase for this task, requiring five successive correct choices measured across ten trials for all dogs, irrespective of performance. The dogs in our study therefore had greater fidelity to the ‘A’ pot due to additional training trials. Running multiple test trials in each task would have enhanced the accuracy of the measurements of dog cognitive performance, and therefore not only increased variation within task, but also increased the ability to identify a relationship in performance between tasks. This was confirmed by the PCA analysis where several measurements of inhibitory control across both tasks (error rate, task completion time), loaded onto a single principal component that explained 37% of the behavioural variation among dogs. Furthermore, the sample size in our study (N = 37) was higher than the sample sizes in other studies that compared performance in the A-not-B task with the cylinder or detour task (N = 11 to 34:54). Therefore, the lower sample sizes across these studies, along with variation in the way in which successes and errors in tasks were determined, may have led to a mismatch between our results and those of previous researchers [
23,
52,
54].
Despite a relationship between success in the A-not-B and the detour tasks, showing a common component of cognitive ability, we found no relationship between the error rates across the two tasks. During the detour task, in addition to inhibiting a direct approach to the food, perseveration in the direct path is rewarded, whereas in the A-not-B task, perseveration in choosing the A pot is not rewarded, so the A-not-B task requires a greater degree of cognitive flexibility, in addition to inhibitory control [
52]. Errors in the A-not-B task represent a lack of cognitive flexibility to inhibit the previously rewarded response, whereas errors in the detour task are due to a lack of inhibition to approach the food directly, but may also include aspects of spatial perception and navigation to successfully detour the barrier [
23,
54]. Therefore, a comparison of errors rather than of successes may be inappropriate when comparing the performance between these tasks.
Stress indicators upon first entering the test environment did not provide a good predictor of cognitive performance, despite predictions that levels of arousal would significantly impact on cognitive ability [
7,
32]. There was a positive relationship in the PCA analysis between eye contact and stress indicators, although PC2 did not predict cognitive performance. Increased eye contact has been found to indicate a stronger attachment between the dog and owner [
38], and the positive relationship between stress indicators and eye contact duration suggests that dogs experiencing higher stress levels were also seeking owner reassurance [
55]. Owner presence enhances problem solving ability in dogs, irrespective of the owner’s behaviour [
56]. The lack of a relationship between stress indicators and general cognitive performance may therefore have been due to owners remaining present throughout the tasks, reducing the impact of initial stress levels on cognitive performance.
Previous studies have found effects of both breed type [
16] and sex [
30] on cognitive performance, but we found no support for this in either task. Breed tends to influence the level of attention paid to human signals [
10,
11,
12,
13]. This is considered important in the A-not-B task, where success is related to attention to the experimenter positioning the reward [
57], but not in the detour task, when the behaviour is not demonstrated. Experience has also been found to reduce breed differences in cognitive traits [
13,
14], and so a variable training background across our participants may have reduced the likelihood of finding breed-related differences in behaviour. Neuter status had no effect on performance in either task, confirming previous work indicating that it has no effect on cognitive tasks [
30]. We found no effect of age group on performance in the A-not-B task, but age affected performance in the detour task. Younger dogs were more likely to meet the detour criteria than were older dogs, potentially due to a general decline in cognitive function with age [
58]; however, age appeared not to impact on performance in other detour studies [
7,
11,
16]. A lack of effect of age in the A-not-B task indicates that this response is less likely to be due to cognitive decline in general affecting inhibitory control, but may potentially be due to differences in navigation and spatial perception. For example, allocentric navigation (use of landmarks to navigate a route) shows a significant decline with age in dogs [
59], and exploratory behaviours are higher in younger dogs [
60]. This may lead to a greater tendency in younger dogs to navigate successfully around the barrier and remember their route on subsequent trials.
While this study clearly points towards an effect of prior training discipline on inhibitory control in pet dogs, the results do not show strong support for training enhancing cognitive performance. This may be due to limitations in the current study. For example, a relatively small sample size in comparison to previous studies may have reduced the ability to detect the effects of sex or breed type [
16,
30]. Additionally, the study relied on owner identification of the breed of participants; however, breed identification by visual characteristics can be unreliable [
61]. The incorrect allocation of dogs to specific breed groups may have confounded the results, and the verification of breed, either by access to registration details or DNA analysis, may be beneficial in future studies. Testing dog performance on only two tasks may also have reduced our ability to detect the impact of training experience. Previous studies comparing highly trained dogs to untrained dogs have used a greater range of cognitive tasks, but these only detected an impact of training on some of the tasks used [
23,
54]. Therefore, further work is needed to determine whether pet dog training experience can impact on performance across a wider range of cognitive tasks.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}