
Evidence of the Existence of Site-Specific Female Contact Pheromones Involved in the Sexual Interaction Behavior of the Pacific Whiteleg Shrimp Penaeus vannamei
 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Extraction of Liposoluble Cuticular Compounds
2.2. Bioassays
2.2.1. Bioassay I: Effect of Immature Female Extracts on the Mating Behavior of a Male Shrimp
2.2.2. Bioassay II: Effect of Mature Female Extracts on the Mating Behavior of a Male Shrimp
2.3. Data Collection and Statistical Analysis
3. Results
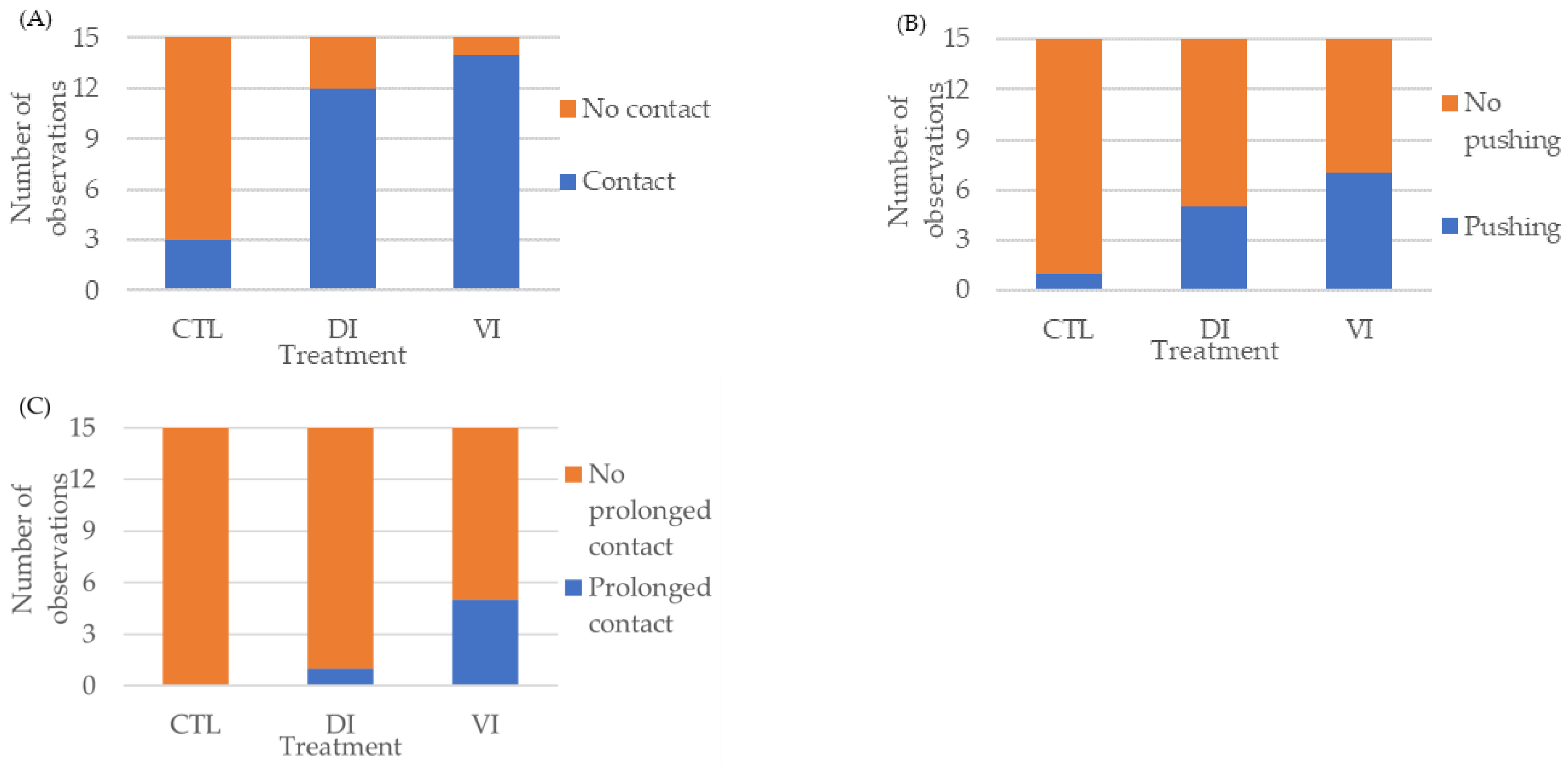
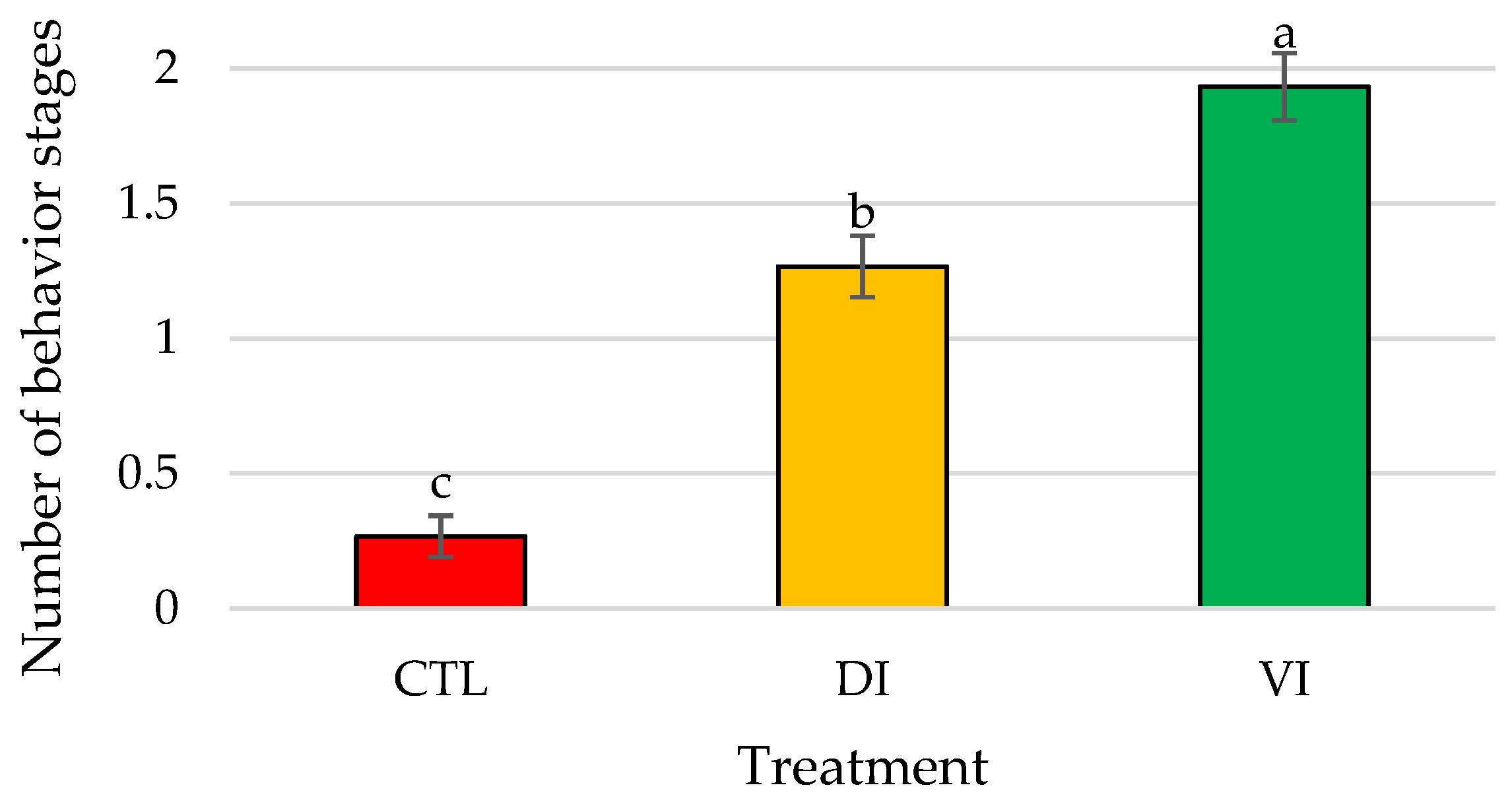
3.1. Bioassay I: Effect of Immature Female Extracts on the Mating Behavior of Shrimp Males
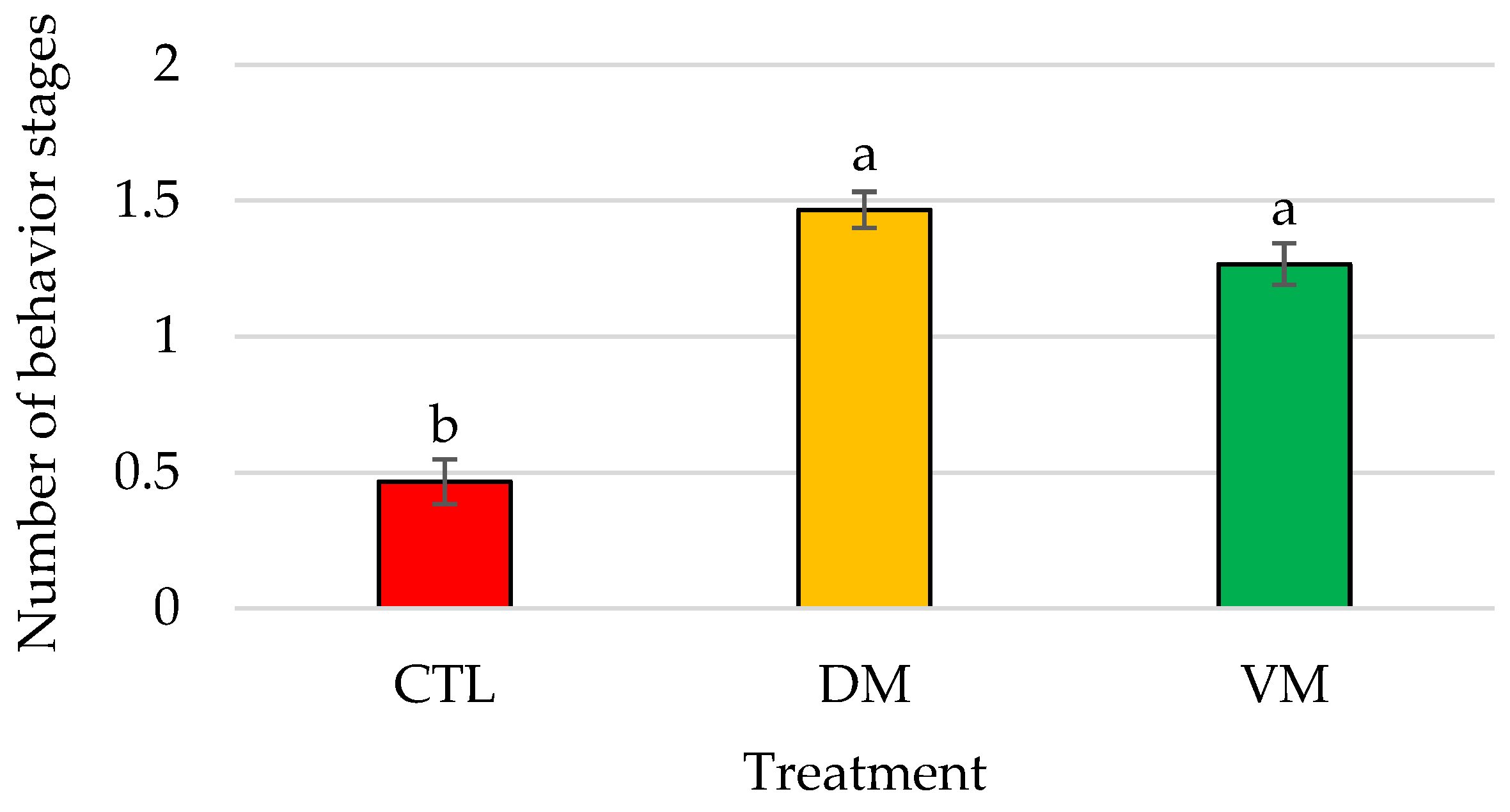
3.2. Bioassay II: Effect of Mature Female Extracts on the Mating Behavior of Shrimp Males
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2022. In Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar]
- Alfaro-Montoya, J.; Braga, A.; Umaña-Castro, R. Research frontiers in penaeid shrimp reproduction: Future trends to improve commercial production. Aquaculture 2019, 503, 70–87. [Google Scholar] [CrossRef]
- Law, J.H.; Regnier, F.E. Pheromones. Perception 1971, 40, 533–548. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Terschak, J.A.; Harley, M.A.; Lin, J.; Hardege, J.D. Simultaneously hermaphroditic shrimp use lipophilic cuticular hydrocarbons as contact sex pheromones. PLoS ONE 2011, 6, e17720. [Google Scholar] [CrossRef] [PubMed]
- Caskey, J.L.; Watson, G.M.; Bauer, R.T. Studies on contact sex pheromones of the caridean shrimp Palaemonetes pugio: II. The role of glucosamine in mate recognition. Invertebr. Reprod. Dev. 2009, 53, 105–116. [Google Scholar] [CrossRef]
- Barki, A.; Ilan, K. Mating behavior and behavioral assay for female receptivity in the red-claw crayfish Cherax quadricarinatus. J. Crustac. Biol. 1999, 19, 493–497. [Google Scholar] [CrossRef]
- Ryan, E. Pheromone: Evidence in a decapod crustacean. Science 1966, 151, 340–341. [Google Scholar] [CrossRef] [PubMed]
- Barkari, A.; Jones, C.; Karplus, I. Chemical Communication and Aquaculture of Decapod Crustaceans: Needs, Problems, and Possible Solutions. In Chemical Communication in Crustaceans; Breithaupt, T., Thiel, M., Eds.; Springer Science+Business: New York, NY, USA, 2011; pp. 485–510. [Google Scholar] [CrossRef]
- Montemayor, J. Uso de Feromonas y Aminas Biogénicas como Atractantes en Alimento para Langostinos Macrobrachium rosenbergii. Master’s Thesis, Universidad Autónoma de Nuevo León, Nuevo León, México, 1995. [Google Scholar]
- Kruangkum, T.; Saetan, J.; Chotwiwatthanakun, C.; Vanichviriyakit, R.; Thongroda, S.; Thintharua, P.; Tulyanandae, T.; Sobhon, P. Co-culture of males with late premolt to early postmolt female giant freshwater prawns, Macrobrachium rosenbergii resulted in greater abundances of insulin-like androgenic gland hormone and gonad maturation in male prawns as a result of olfactory receptors. Anim. Reprod. Sci. 2019, 210, 106198. [Google Scholar] [CrossRef] [PubMed]
- Kamio, M.; Yambe, H.; Fusetani, N. Chemical cues for intraspecific chemical communication and interspecific interactions in aquatic environments: Applications for fisheries and aquaculture. Fish. Sci. 2022, 88, 203–239. [Google Scholar] [CrossRef]
- Kelly, T.R.; Fitzgibbon, Q.P.; Smith, G.G.; Banks, T.M.; Ventura, T. Tropical rock lobster (Panulirus ornatus) uses chemoreception via the antennular lateral flagellum to identify conspecific ecdysis. Sci. Rep. 2023, 13, 12409. [Google Scholar] [CrossRef]
- Dunham, P.J. Pheromones and behaviour in Crustacea. In Endocrinology of Selected Invertebrate Types; Laufer, H., Downer, R., Eds.; Liss: New York, NY, USA, 1988; p. 500. [Google Scholar] [CrossRef]
- Bamber, S.D.; Naylor, E. Chemical communication and behavioural interaction between sexually mature male and female shore crabs (Carcinus maenas). J. Mar. Biol. Assoc. UK 1996, 76, 691–699. [Google Scholar] [CrossRef]
- Misamore, M.; Browdy, C. Mating behavior in the white shrimps Penaeus setiferus and P. vannamei: A generalized model for mating in Penaeus. J. Crustac. Biol. 1996, 16, 61–70. [Google Scholar] [CrossRef]
- Yano, I.; Kanna, R.A.; Oyama, R.N.; Wyban, J.A. Mating behaviour in the penaeid shrimp Penaeus vannamei. Mar. Biol. 1988, 97, 171–175. [Google Scholar] [CrossRef]
- Ayub, Z.; Ahmed, M. A description of the ovarian development stages of penaeid shrimps form the coast of Pakistan. Aquac. Res. 2002, 33, 767–776. [Google Scholar] [CrossRef]
- Bauer, R.T.; Abdalla, J.H. Male mating tactics in the shrimp Palaemonetes pugio (Decapoda, Caridea): Precopulatory mate guarding vs. pure searching. Ethology 2001, 107, 185–199. [Google Scholar] [CrossRef]
- Kelly, L.S.; Snell, T.W. Role of surface glycoproteins in mate-guarding of the marine harpacticoid Tigriopus japonicus. Mar. Biol. 1998, 130, 605–612. [Google Scholar] [CrossRef]
- Raviv, S.; Parnes, S.; Sagi, A. Coordination of reproduction and molt in decapods. In Reproductive Biology of Crustaceans; Mente, E., Ed.; Science Publishers: Enfeld, NH, USA, 2008; pp. 365–390. [Google Scholar] [CrossRef]
- Wall, D.; Paterson, B.; Mohan, R. Behaviour of juvenile mud crabs Scylla serrata in aquaculture: Response to odours of moulting or injured crabs. Appl. Anim. Behav. Sci. 2009, 121, 63–73. [Google Scholar] [CrossRef]
- Adams, J.A.; Moore, P.A. Discrimination of conspecific male molt odor signals by male crayfish, Orconectes rusticus. J. Crustac. Biol. 2003, 23, 7–14. [Google Scholar] [CrossRef]
- Breithaupt, T.; Thiel, M. Chemical communication in crustaceans: Research challenges for the twenty-first century. In Chemical Communication in Crustaceans; Breithaupt, T., Thiel, M., Eds.; Springer Science+Business: New York, NY, USA, 2011; pp. 3–22. [Google Scholar] [CrossRef]
- Hardege, J.D.; Jennings, A.; Hayden, D.; Müller, C.T.; Pascoe, D.; Bentley, M.G.; Clare, A.S. Novel behavioral assay and partial purification of a female-derived sex pheromone in Carcinus maenas. Mar. Ecol. Prog. Ser. 2002, 244, 179–189. [Google Scholar] [CrossRef]
- Asai, N.; Fusetani, N.; Matsunaga, S.; Sasaki, J. Sex pheromones of the hair crab Erimacrus isenbeckii. Part 1. Isolation and structures of novel ceramides. Tetrahedron 2000, 56, 9895–9899. [Google Scholar] [CrossRef]
- Kamio, M.; Araki, M.; Nagayama, T.; Matsunaga, S.; Fusetani, N. Behavioral and electrophysiological experiments suggest that the antennular outer flagellum is the site of pheromone reception in the male helmet crab Telmessus cheiragonus. Biol. Bull. 2005, 208, 12–19. [Google Scholar] [CrossRef]
- Parker, G.A. Courtship persistence and female-guarding as male time investment strategies. Behaviour 1974, 48, 157–183. [Google Scholar] [CrossRef]
- Calderón-Pérez, J.A.; Rendón-Rodríguez, S.; Solís-Ibarra, R. Daily cycle and body characteristics of mating Litopenaeus vannamei shrimps (Decapoda: Penaeidae) in the wild off southern Sinaloa, Mexico. Rev. Biol. Trop. 2007, 55, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Hardege, J.D.; Bartels-Hardege, H.D.; Fletcher, N.; Terschak, J.A.; Harley, M.; Smith, M.A.; Davidson, L.; Hayden, D.; Müller, C.T.; Lorch, M.; et al. Identification of a female sex pheromone in Carcinus maenas. Mar. Ecol. Prog. Ser. 2011, 436, 177–189. [Google Scholar] [CrossRef]
- Ginzel, M.D.; Blomquist, G.J.; Millar, J.G.; Hanks, L.M. Role of contact pheromones in mate recognition in Xylotrechus colonus. J. Chem. Ecol. 2003, 29, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Stoffolano, J.G.; Schauber, E.; Yin, C.M.; Tillman, J.A.; Blomquist, G.J. Cuticular hydrocarbons and their role in copulatory behavior in Phormia regina (Meigen). J. Insect. Physiol. 1997, 43, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Caskey, J.L.; Bauer, R.T. Behavioral tests for a possible contact sex pheromone in the caridean shrimp Palaemonetes pugio. J. Crustac. Biol. 2005, 25, 571–576. [Google Scholar] [CrossRef]
- Derby, C.D. The crustacean antennule: A complex organ adapted for lifelong function in diverse environments and lifestyles. Biol. Bull. 2021, 240, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Harzsch, S.; Krieger, J. Crustacean olfactory systems: A comparative review and a crustacean perspective on insect olfatory systems. Prog. Neurobiol. 2018, 161, 23–60. [Google Scholar] [CrossRef] [PubMed]
- Dunham, D.W.; Oh, J.W. Chemical sex discrimination in the crayfish Procambarus clarkii: Role of antennules. J. Chem. Ecol. 1992, 18, 2363–2372. [Google Scholar] [CrossRef]
- Ache, B.W. Chemoreception and thermoreception. In The Biology of Crustacea; Atwood, H.L., Sandeman, D.C., Eds.; Academic press: Cambridge, MA, USA, 1982; Volume 3, pp. 368–398. [Google Scholar]
- Eap, D.; Correa, S.; Ngo-Vu, H.; Derby, C.D. Chemosensory basis of feeding behavior in Pacific white shrimp, Litopenaeus vannamei. Biol. Bull. 2020, 239, 115–131. [Google Scholar] [CrossRef]
- Wyatt, T.D. Pheromones. Curr. Biol. 2017, 27, R739–R743. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, H. The crustacean cuticle: Structure, composition and mineralization. Front. Biosci. Elite 2012, 4, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.W.; Blomquist, G.J. Ecological, behavioral, and biochemical aspects of insect hydrocarbons. Annu. Rev. Entomol. 2005, 50, 371–393. [Google Scholar] [CrossRef] [PubMed]
- Billeter, J.C.; Atallah, J.; Krupp, J.J.; Millar, J.G.; Levine, J.D. Specialized cells tag sexual and species identity in Drosophila melanogaster. Nature 2009, 461, 987–991. [Google Scholar] [CrossRef]
- Rundle, H.D.; Chenoweth, S.F.; Doughty, P.; Blows, M.W. Divergent selection and the evolution of signal traits and mating preferences. PLoS Biol. 2005, 3, 1988–1995. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Degree | Behavior | Description |
|---|---|---|
| 0 | No response | No change in behavior was observed. |
| 1 | Contact | The male had at least one contact with the artificial female and initiated an examination with the antennules. |
| 2 | Pushing | After contact, the male took impulse to push the artificial female. |
| 3 | Prolonged contact | After contact and pushing, the antennules of male maintained constant contact with the artificial female for at least 10 s. |
| Behavior | Treatment | p-Value | ||
|---|---|---|---|---|
| CTL | DI | VI | ||
| Contact | 0.40 ± 0.24 b | 2.07 ± 0.54 a | 2.80 ± 0.50 a | <0.01 |
| Pushing | 0.07 ± 0.07 b | 0.40 ± 0.16 ab | 0.73 ± 0.25 a | <0.05 |
| Behavior | Treatment | p-Value | ||
|---|---|---|---|---|
| CTL | DM | VM | ||
| Contact | 0.87 ± 0.41 b | 2.67 ± 0.51 a | 3.60 ± 0.40 a | <0.01 |
| Pushing | 0.07 ± 0.07 | 0.53 ± 0.17 | 0.40 ± 0.16 | 0.052 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Vera, J.A.; Ponce-Rivas, E.; Braga, A.; Paniagua-Chávez, C.G.; Alfaro-Montoya, J.; Rosales-Leija, M. Evidence of the Existence of Site-Specific Female Contact Pheromones Involved in the Sexual Interaction Behavior of the Pacific Whiteleg Shrimp Penaeus vannamei. Animals 2024, 14, 1523. https://doi.org/10.3390/ani14111523
Gutiérrez-Vera JA, Ponce-Rivas E, Braga A, Paniagua-Chávez CG, Alfaro-Montoya J, Rosales-Leija M. Evidence of the Existence of Site-Specific Female Contact Pheromones Involved in the Sexual Interaction Behavior of the Pacific Whiteleg Shrimp Penaeus vannamei. Animals. 2024; 14(11):1523. https://doi.org/10.3390/ani14111523
Chicago/Turabian StyleGutiérrez-Vera, José A., Elizabeth Ponce-Rivas, André Braga, Carmen G. Paniagua-Chávez, Jorge Alfaro-Montoya, and Misael Rosales-Leija. 2024. "Evidence of the Existence of Site-Specific Female Contact Pheromones Involved in the Sexual Interaction Behavior of the Pacific Whiteleg Shrimp Penaeus vannamei" Animals 14, no. 11: 1523. https://doi.org/10.3390/ani14111523
APA StyleGutiérrez-Vera, J. A., Ponce-Rivas, E., Braga, A., Paniagua-Chávez, C. G., Alfaro-Montoya, J., & Rosales-Leija, M. (2024). Evidence of the Existence of Site-Specific Female Contact Pheromones Involved in the Sexual Interaction Behavior of the Pacific Whiteleg Shrimp Penaeus vannamei. Animals, 14(11), 1523. https://doi.org/10.3390/ani14111523