
A New Cell Line from the Brain of Red Hybrid Tilapia (Oreochromis spp.) for Tilapia Lake Virus Propagation

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Preparation for Primary Cell Cultures
2.2. Subculture and Maintenance
2.3. Cryopreservation and Recovery
2.4. Cell Growth Characteristics
2.5. Identification of Origin Cell Line Using Mitochondrial Markers
2.6. Detection of Mycoplasma and Snakehead Retrovirus Contamination in RHTiB Cell Line
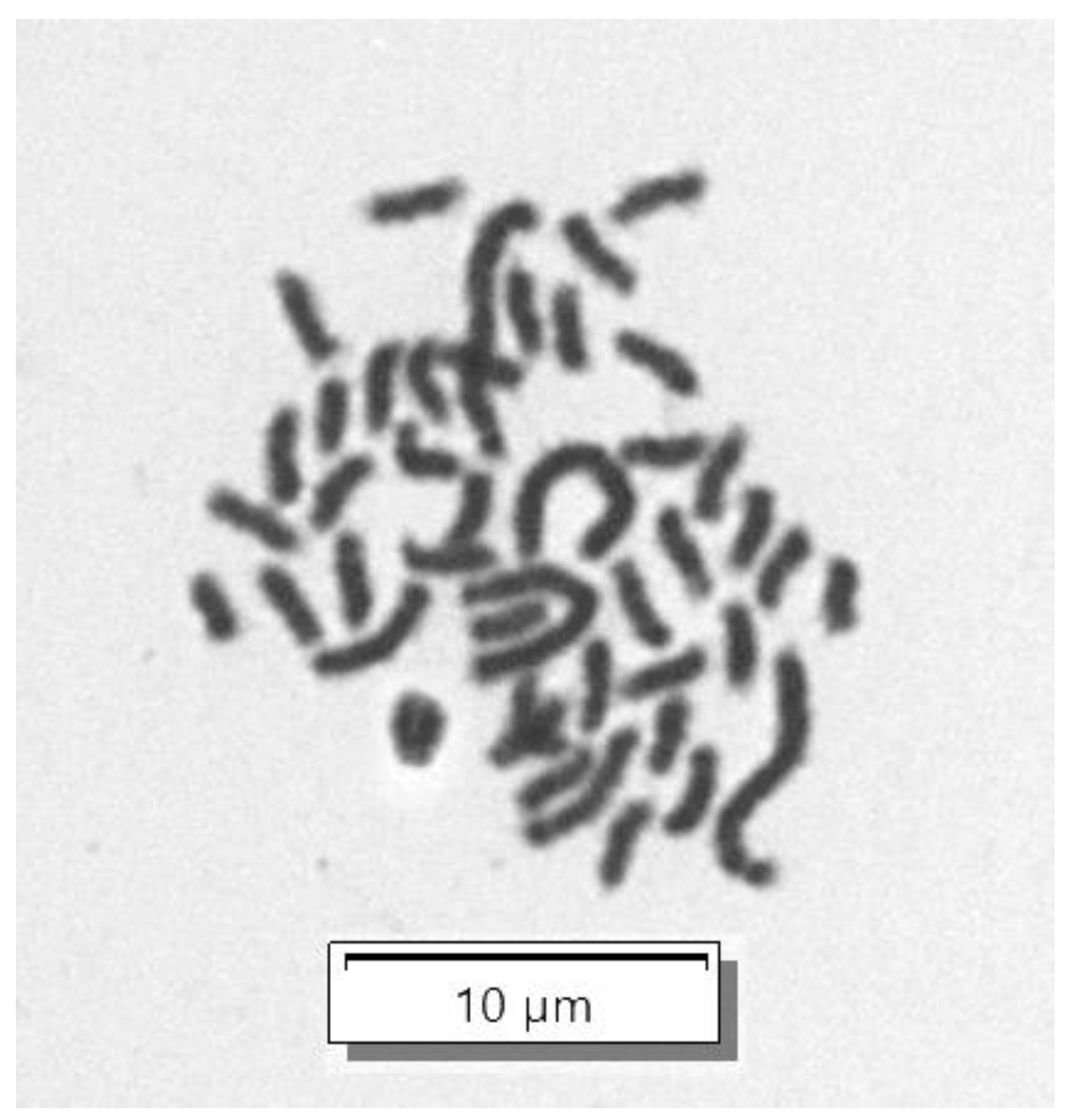
2.7. Chromosome Analysis of RHTiB Cells
2.8. Immunofluorescent Assay to Detect TiLV in RHTiB Cell Line
2.9. Propagation of TiLV in RHTiB Cell Line under Varying pH and Temperature Conditions
2.10. Statistical Analysis
3. Results
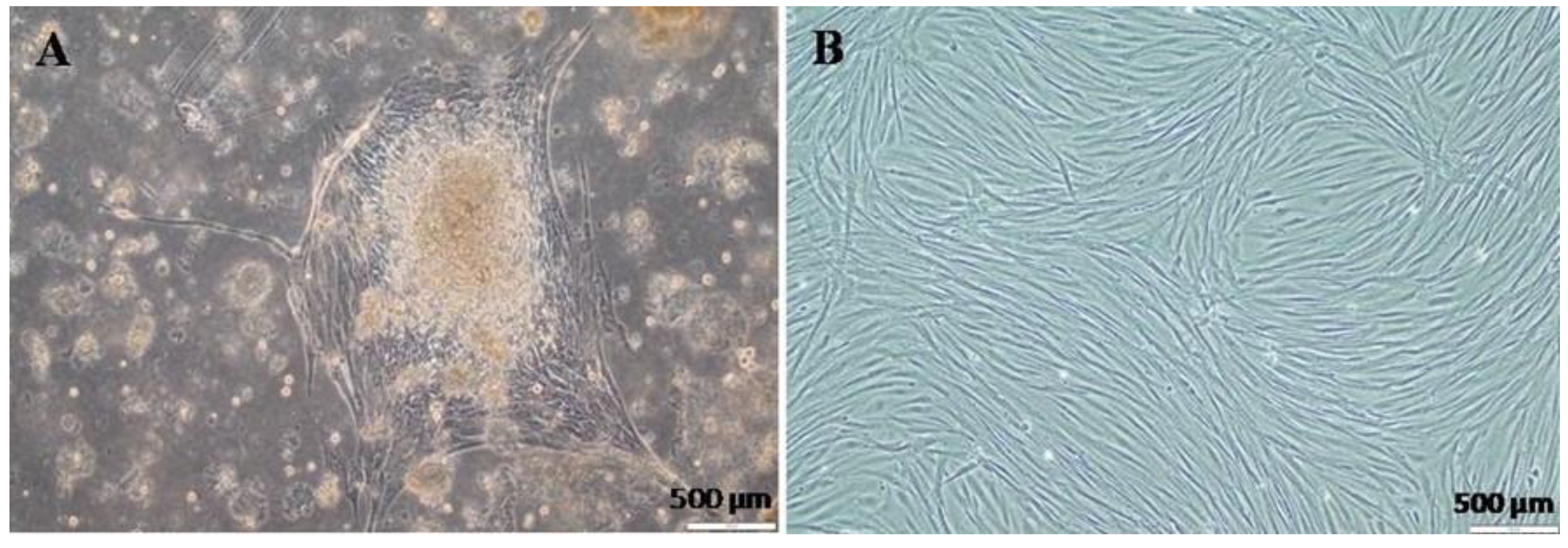
3.1. Primary Cell Isolation and Culture
3.2. Optimization of RHTiB Cell Growth Conditions
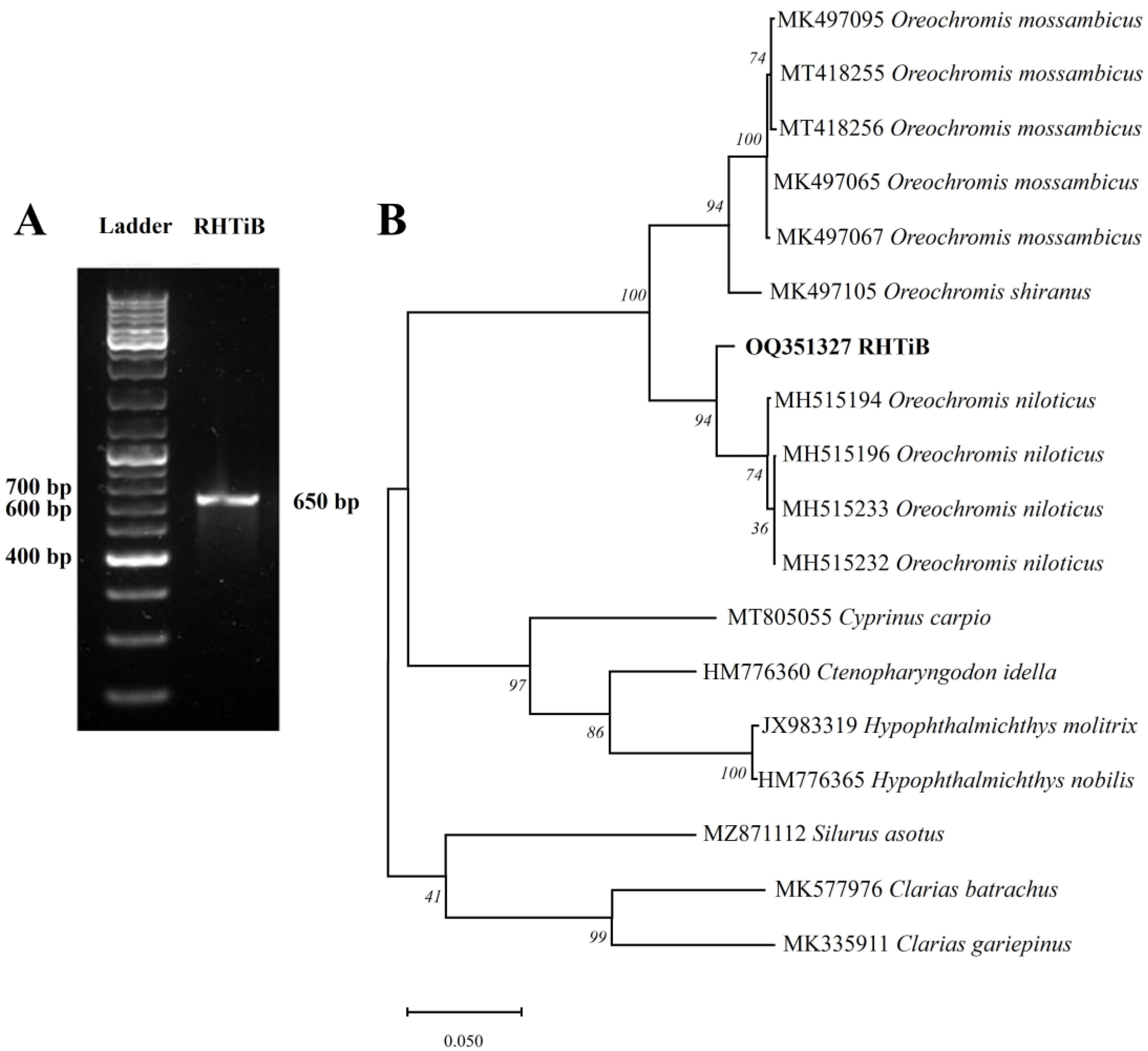
3.3. Species Identification of RHTiB Cell Line Using cox1 Gene
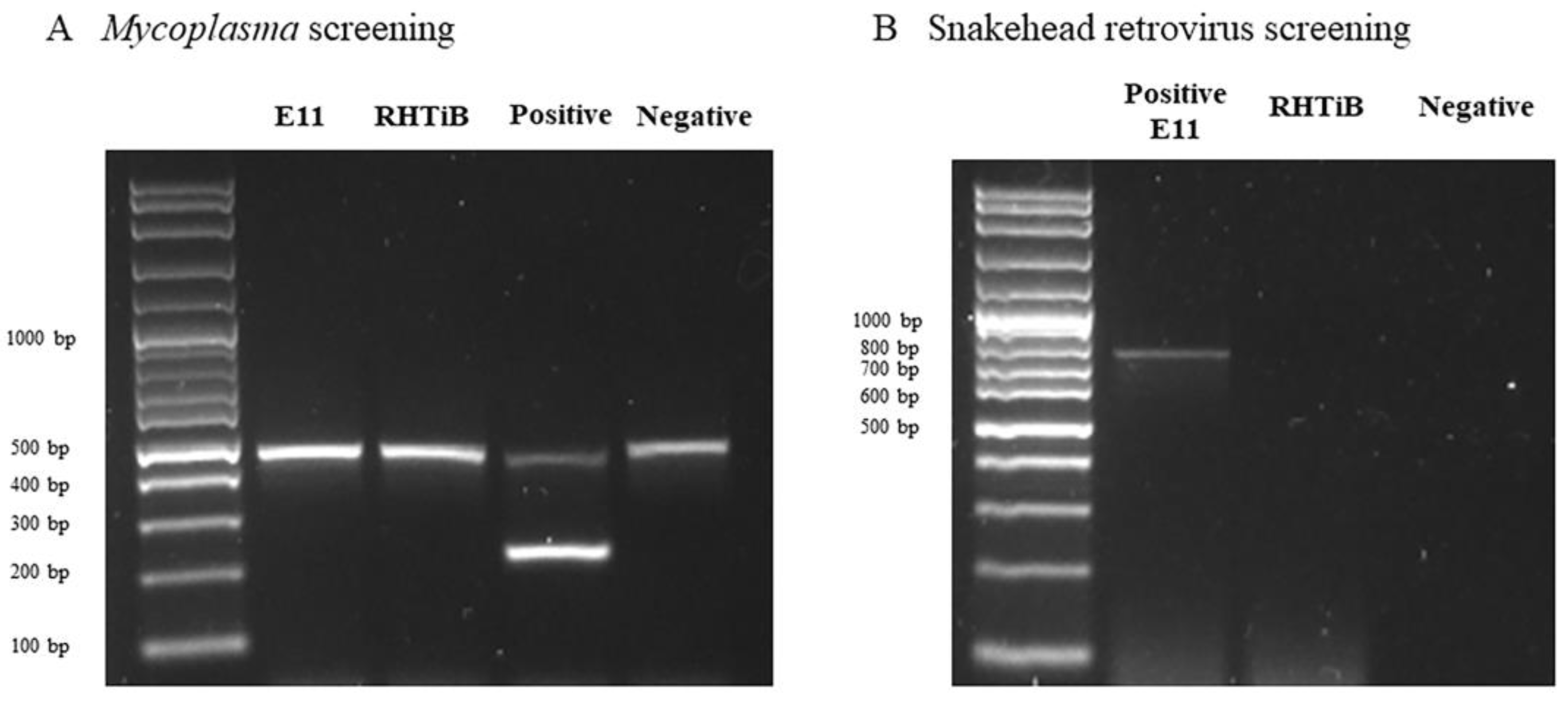
3.4. Detection of Mycoplasma and Snakehead Retrovirus Contamination
3.5. Detection of TiLV Infection in RHTiB Using Immunofluorescence
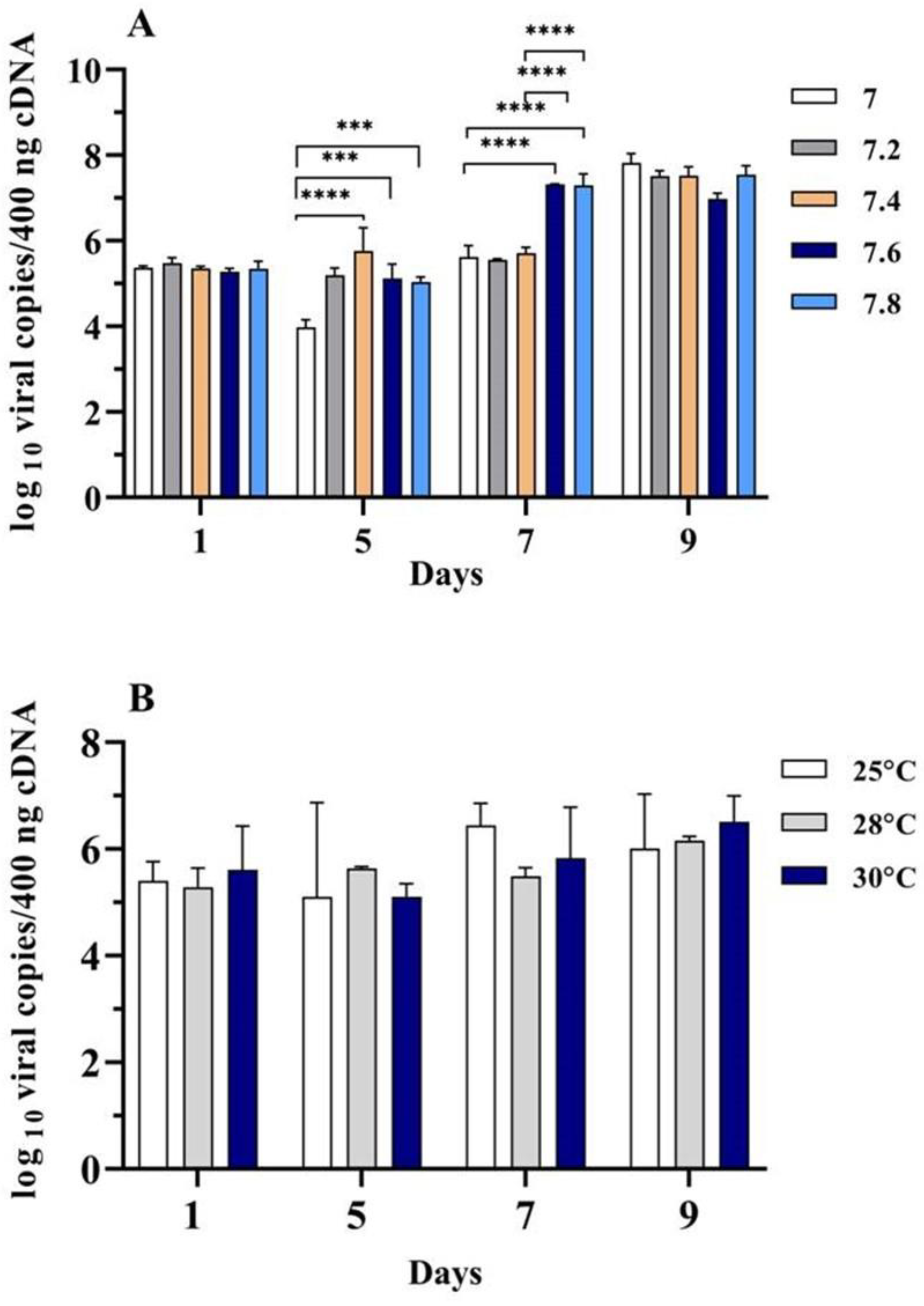
3.6. Propagation of TiLV in RHTiB Cells under Different pH and Temperature Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Del-Pozo, J.; Mishra, N.; Kabuusu, R.; Cheetham, S.; Eldar, A.; Bacharach, E.; Lipkin, W.I.; Ferguson, H.W. Syncytial Hepatitis of Tilapia (Oreochromis niloticus L.) is Associated with Orthomyxovirus-Like Virions in Hepatocytes. Vet. Pathol. 2017, 54, 164–170. [Google Scholar] [CrossRef]
- Wang, B.; Thompson, K.D.; Wangkahart, E.; Yamkasem, J.; Bondad-Reantaso, M.G.; Tattiyapong, P.; Jian, J.; Surachetpong, W. Strategies to enhance tilapia immunity to improve their health in aquaculture. Rev. Aquac. 2023, 15, 41–56. [Google Scholar] [CrossRef]
- FAO. In Brief to the State of World Fisheries and Aquaculture 2022; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- Koonin, E.V.; Krupovic, M.; Surachetpong, W.; Wolf, Y.I.; Kuhn, J.H. ICTV Virus Taxonomy Profile: Amnoonviridae 2023. J. Gen. Virol. 2023, 104, 001903. [Google Scholar] [CrossRef]
- Eyngor, M.; Zamostiano, R.; Kembou Tsofack, J.E.; Berkowitz, A.; Bercovier, H.; Tinman, S.; Lev, M.; Hurvitz, A.; Galeotti, M.; Bacharach, E.; et al. Identification of a novel RNA virus lethal to tilapia. J. Clin. Microbiol. 2014, 52, 4137–4146. [Google Scholar] [CrossRef]
- Surachetpong, W.; Janetanakit, T.; Nonthabenjawan, N.; Tattiyapong, P.; Sirikanchana, K.; Amonsin, A. Outbreaks of Tilapia Lake Virus Infection, Thailand, 2015–2016. Emerg. Infect. Dis. 2017, 23, 1031–1033. [Google Scholar] [CrossRef]
- Jansen, M.D.; Dong, H.T.; Mohan, C.V. Tilapia lake virus: A threat to the global tilapia industry? Rev. Aquac. 2019, 11, 725–739. [Google Scholar] [CrossRef]
- Prasartset, T.; Surachetpong, W. Simultaneous detection of three important viruses affecting tilapia using a multiplex PCR assay. J. Fish Dis. 2023, 46, 459–464. [Google Scholar] [CrossRef]
- Surachetpong, W.; Roy, S.R.K.; Nicholson, P. Tilapia lake virus: The story so far. J. Fish Dis. 2020, 43, 1115–1132. [Google Scholar] [CrossRef]
- Kembou-Ringert, J.E.; Steinhagen, D.; Readman, J.; Daly, J.M.; Adamek, M. Tilapia Lake Virus Vaccine Development: A Review on the Recent Advances. Vaccines 2023, 11, 251. [Google Scholar] [CrossRef]
- Tran, T.H.; Nguyen, V.T.H.; Bui, H.C.N.; Tran, Y.B.T.; Tran, H.T.T.; Le, T.T.T.; Vu, H.T.T.; Ngo, T.P.H. Tilapia Lake Virus (TiLV) from Vietnam is genetically distantly related to TiLV strains from other countries. J. Fish Dis. 2022, 45, 1389–1401. [Google Scholar] [CrossRef]
- He, T.; Zhang, Y.-Z.; Gao, L.-H.; Miao, B.; Zheng, J.-S.; Pu, D.-C.; Zhang, Q.-Q.; Zeng, W.-W.; Wang, D.-S.; Su, S.-Q.; et al. Identification and pathogenetic study of tilapia lake virus (TiLV) isolated from naturally diseased tilapia. Aquaculture 2023, 565, 739166. [Google Scholar] [CrossRef]
- Nicholson, P.; Rawiwan, P.; Surachetpong, W. Detection of Tilapia Lake Virus Using Conventional RT-PCR and SYBR Green RT-qPCR. J. Vis. Exp. 2018, 141, e58596. [Google Scholar] [CrossRef]
- Tattiyapong, P.; Dachavichitlead, W.; Surachetpong, W. Experimental infection of Tilapia Lake Virus (TiLV) in Nile tilapia (Oreochromis niloticus) and red tilapia (Oreochromis spp.). Vet. Microbiol. 2017, 207, 170–177. [Google Scholar] [CrossRef]
- Swaminathan, T.R.; Raja, A.; Dharmaratnam, A.; Nithianantham, S.R. Comparative sensitivity of three new cell lines developed from gill, liver and brain tissues of goldfish, Carassius auratus (L.) to cyprinid herpesvirus-2 (CyHV-2). J. Virol. Methods 2021, 291, 114069. [Google Scholar] [CrossRef]
- Li, B.; Zheng, S.; Wang, Y.; Wang, Q.; Li, Y.; Yin, J.; Ren, Y.; Shi, C.; Zhao, Z.; Jiang, Z.; et al. Susceptibilities of ten fish cell lines to infection with Tilapia lake virus. Microb. Pathog. 2022, 166, 105510. [Google Scholar] [CrossRef]
- Goswami, M.; Yashwanth, B.S.; Trudeau, V.; Lakra, W.S. Role and relevance of fish cell lines in advanced in vitro research. Mol. Biol. Rep. 2022, 49, 2393–2411. [Google Scholar] [CrossRef]
- Thangaraj, R.S.; Ravi, C.; Kumar, R.; Dharmaratnam, A.; Valaparambil Saidmuhammed, B.; Pradhan, P.K.; Sood, N. Derivation of two tilapia (Oreochromis niloticus) cell lines for efficient propagation of Tilapia Lake Virus (TiLV). Aquaculture 2018, 492, 206–214. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Zeng, W.; Yin, J.; Li, Y.; Ren, Y.; Shi, C.; Bergmann, S.M.; Zhu, X. Establishment and characterization of a cell line from tilapia brain for detection of tilapia lake virus. J. Fish Dis. 2018, 41, 1803–1809. [Google Scholar] [CrossRef]
- Long, Y.; Liu, R.; Song, G.; Li, Q.; Cui, Z. Establishment and characterization of a cold-sensitive neural cell line from the brain of tilapia (Oreochromis niloticus). J. Fish Biol. 2021, 98, 842–854. [Google Scholar] [CrossRef]
- Yadav, M.K.; Rastogi, A.; Criollo Joaquin, M.P.; Verma, D.K.; Rathore, G.; Swaminathan, T.R.; Paria, A.; Pradhan, P.K.; Sood, N. Establishment and characterization of a continuous cell line from heart of Nile tilapia Oreochromis niloticus and its susceptibility to tilapia lake virus. J. Virol. Methods 2021, 287, 113989. [Google Scholar] [CrossRef]
- Li, Y.; Huang, Y.; Cai, J.; Jiang, D.; Jian, J.C.; Lu, Y.S.; Wang, B. Establishment of an astrocyte-like cell line from the brain of tilapia (Oreochromis niloticus) for virus pathogenesis and a vitro model of the blood-brain barrier. J. Fish Dis. 2022, 45, 1451–1462. [Google Scholar] [CrossRef]
- Swaminathan, T.R.; Basheer, V.S.; Gopalakrishnan, A.; Rathore, G.; Chaudhary, D.K.; Kumar, R.; Jena, J.K. Establishment of caudal fin cell lines from tropical ornamental fishes Puntius fasciatus and Pristolepis fasciata endemic to the Western Ghats of India. Acta Trop. 2013, 128, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Lertwanakarn, T.; Trongwongsa, P.; Yingsakmongkol, S.; Khemthong, M.; Tattiyapong, P.; Surachetpong, W. Antiviral Activity of Ribavirin against Tilapia tilapinevirus in Fish Cells. Pathogens 2021, 10, 1616. [Google Scholar] [CrossRef]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D. DNA barcoding Australia’s fish species. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 1847–1857. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nishizawa, T.; Kokawa, Y.; Wakayama, T.; Kinoshita, S.; Yoshimizu, M. Enhanced propagation of fish nodaviruses in BF-2 cells persistently infected with snakehead retrovirus (SnRV). Dis. Aquat. Organ. 2008, 79, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Piewbang, C.; Tattiyapong, P.; Techangamsuwan, S.; Surachetpong, W. Tilapia lake virus immunoglobulin G (TiLV IgG) antibody: Immunohistochemistry application reveals cellular tropism of TiLV infection. Fish Shellfish Immunol. 2021, 116, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Yamkasem, J.; Tattiyapong, P.; Surachetpong, W. Comparison of reverse transcription polymerase chain reaction methods for the detection of tilapia lake virus. Bull. Eur. Assoc. Fish Pathol. 2020, 40, 229–239. [Google Scholar]
- Bacharach, E.; Mishra, N.; Briese, T.; Zody, M.C.; Kembou Tsofack, J.E.; Zamostiano, R.; Berkowitz, A.; Ng, J.; Nitido, A.; Corvelo, A.; et al. Characterization of a Novel Orthomyxo-like Virus Causing Mass Die-Offs of Tilapia. mBio 2016, 7, e00431-16. [Google Scholar] [CrossRef]
- Waiyamitra, P.; Piewbang, C.; Techangamsuwan, S.; Liew, W.C.; Surachetpong, W. Infection of Tilapia tilapinevirus in Mozambique Tilapia (Oreochromis mossambicus), a Globally Vulnerable Fish Species. Viruses 2021, 13, 1104. [Google Scholar] [CrossRef]
- Gardell, A.M.; Qin, Q.; Rice, R.H.; Li, J.; Kültz, D. Derivation and osmotolerance characterization of three immortalized tilapia (Oreochromis mossambicus) cell lines. PLoS ONE 2014, 9, e95919. [Google Scholar] [CrossRef]
- Tsuji, K.; Ojima, M.; Otabe, K.; Horie, M.; Koga, H.; Sekiya, I.; Muneta, T. Effects of Different Cell-Detaching Methods on the Viability and Cell Surface Antigen Expression of Synovial Mesenchymal Stem Cells. Cell Transplant. 2017, 26, 1089–1102. [Google Scholar] [CrossRef]
- Weiskirchen, S.; Schröder, S.K.; Buhl, E.M.; Weiskirchen, R. A Beginner’s Guide to Cell Culture: Practical Advice for Preventing Needless Problems. Cells 2023, 12, 682. [Google Scholar] [CrossRef]
- Suryakodi, S.; Majeed, S.A.; Taju, G.; Vimal, S.; Sivakumar, S.; Ahmed, A.N.; Shah, F.A.; Bhat, S.A.; Sarma, D.; Begum, A.; et al. Development and characterization of novel cell lines from kidney and eye of rainbow trout, Oncorhynchus mykiss for virological studies. Aquaculture 2021, 532, 736027. [Google Scholar] [CrossRef]
- Zeng, W.; Dong, H.; Chen, X.; Bergmann, S.M.; Yang, Y.; Wei, X.; Tong, G.; Li, H.; Yu, H.; Chen, Y. Establishment and characterization of a permanent heart cell line from largemouth bass Micropterus salmoides and its application to fish virology and immunology. Aquaculture 2022, 547, 737427. [Google Scholar] [CrossRef]
- Soni, P.; Pradhan, P.K.; Swaminathan, T.R.; Sood, N. Development, characterization and application of a new epithelial cell line from caudal fin of Pangasianodon hypophthalmus (Sauvage 1878). Acta Trop. 2018, 182, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, T.R.; Dharmaratnam, A.; Raja, S.A.; Raj, N.S.; Lal, K.K. Establishment and cryopreservation of a cell line derived from caudal fin of endangered catfish Clarias dussumieri Valenciennes, 1840. J. Fish Biol. 2020, 96, 722–730. [Google Scholar] [CrossRef] [PubMed]
- Subbiahanadar Chelladurai, K.; Selvan Christyraj, J.D.; Rajagopalan, K.; Yesudhason, B.V.; Venkatachalam, S.; Mohan, M.; Chellathurai Vasantha, N.; Selvan Christyraj, J.R.S. Alternative to FBS in animal cell culture—An overview and future perspective. Heliyon 2021, 7, e07686. [Google Scholar] [CrossRef] [PubMed]
- Pilgrim, C.R.; McCahill, K.A.; Rops, J.G.; Dufour, J.M.; Russell, K.A.; Koch, T.G. A Review of Fetal Bovine Serum in the Culture of Mesenchymal Stromal Cells and Potential Alternatives for Veterinary Medicine. Front. Vet. Sci. 2022, 9, 859025. [Google Scholar] [CrossRef]
- Pedrazzani, A.S.; Quintiliano, M.H.; Bolfe, F.; Sans, E.C.O.; Molento, C.F.M. Tilapia On-Farm Welfare Assessment Protocol for Semi-intensive Production Systems. Front. Vet. Sci. 2020, 7, 606388. [Google Scholar] [CrossRef]
- Zhou, Y.; Raphael, R.M. Solution pH alters mechanical and electrical properties of phosphatidylcholine membranes: Relation between interfacial electrostatics, intramembrane potential, and bending elasticity. Biophys. J. 2007, 92, 2451–2462. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, A.; Yadav, M.K.; Joaquin, M.P.C.; Verma, D.K.; Swaminathan, T.R.; Kushwaha, B.; Paria, A.; Pradhan, P.K.; Sood, N. Development of cell lines from brain, spleen and heart of ornamental blood parrot cichlid and their susceptibility to Tilapia tilapinevirus. Aquaculture 2022, 561, 738711. [Google Scholar] [CrossRef]
- Abu Rass, R.; Kembou-Ringert, J.E.; Zamostiano, R.; Eldar, A.; Ehrlich, M.; Bacharach, E. Mapping of Tilapia Lake Virus entry pathways with inhibitors reveals dependence on dynamin activity and cholesterol but not endosomal acidification. Front. Cell Dev. Biol. 2022, 10, 1075364. [Google Scholar] [CrossRef] [PubMed]
- Kembou Tsofack, J.E.; Zamostiano, R.; Watted, S.; Berkowitz, A.; Rosenbluth, E.; Mishra, N.; Briese, T.; Lipkin, W.I.; Kabuusu, R.M.; Ferguson, H.; et al. Detection of Tilapia Lake Virus in Clinical Samples by Culturing and Nested Reverse Transcription-PCR. J. Clin. Microbiol. 2017, 55, 759–767. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamad, A.; Khemthong, M.; Trongwongsa, P.; Lertwanakarn, T.; Setthawong, P.; Surachetpong, W. A New Cell Line from the Brain of Red Hybrid Tilapia (Oreochromis spp.) for Tilapia Lake Virus Propagation. Animals 2024, 14, 1522. https://doi.org/10.3390/ani14111522
Mohamad A, Khemthong M, Trongwongsa P, Lertwanakarn T, Setthawong P, Surachetpong W. A New Cell Line from the Brain of Red Hybrid Tilapia (Oreochromis spp.) for Tilapia Lake Virus Propagation. Animals. 2024; 14(11):1522. https://doi.org/10.3390/ani14111522
Chicago/Turabian StyleMohamad, Aslah, Matepiya Khemthong, Pirada Trongwongsa, Tuchakorn Lertwanakarn, Piyathip Setthawong, and Win Surachetpong. 2024. "A New Cell Line from the Brain of Red Hybrid Tilapia (Oreochromis spp.) for Tilapia Lake Virus Propagation" Animals 14, no. 11: 1522. https://doi.org/10.3390/ani14111522
APA StyleMohamad, A., Khemthong, M., Trongwongsa, P., Lertwanakarn, T., Setthawong, P., & Surachetpong, W. (2024). A New Cell Line from the Brain of Red Hybrid Tilapia (Oreochromis spp.) for Tilapia Lake Virus Propagation. Animals, 14(11), 1522. https://doi.org/10.3390/ani14111522