
Comparative Analysis of the Ultrastructure, Bubble Pores, and Composition of Eggshells of Dwarf Layer-White and Guinea Fowl
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Hens and GF
2.2. Egg Quality Measurement
2.3. Measurement of Ultrastructure and Bubble Pores of Eggshell
2.4. Eggshell Chemical Components Measurement
2.5. Statistical Analysis
3. Results and Discussion
3.1. Eggshell Quality
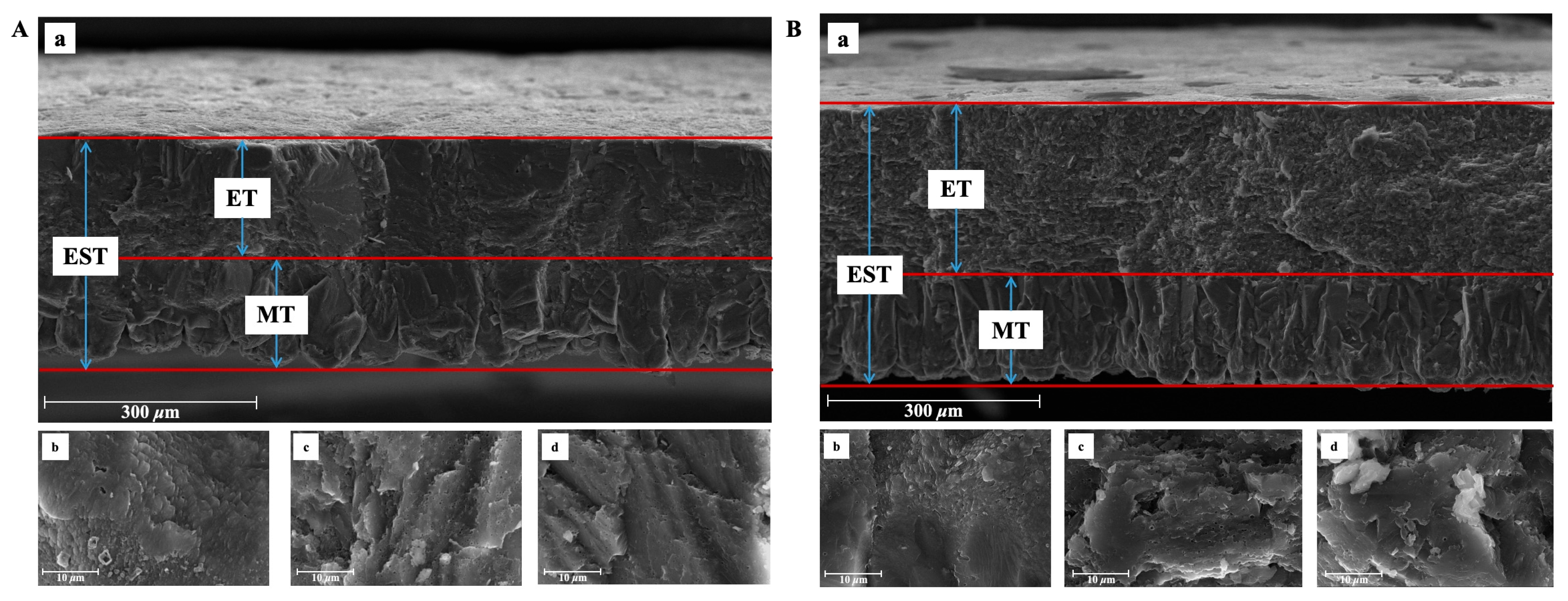
3.2. Eggshell Ultrastructure and Bubble Pores
3.3. Eggshell Bubble Pore
3.4. Eggshell Chemical Components
3.5. Correlations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dunn, I.C.; Joseph, N.T.; Bain, M.; Edmond, A.; Wilson, P.W.; Milona, P.; Nys, Y.; Gautron, J.; Schmutz, M.; Preisinger, R.; et al. Polymorphisms in eggshell organic matrix genes are associated with eggshell quality measurements in pedigree Rhode Island Red hens. Anim. Genet. 2009, 40, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Ning, Z. Research progress on bird eggshell quality defects: A review. Poult. Sci. 2023, 102, 102283. [Google Scholar] [CrossRef] [PubMed]
- Hincke, M.T.; Nys, Y.; Gautron, J.; Mann, K.; Rodriguez-Navarro, A.B.; McKee, M.D. The eggshell: Structure, composition and mineralization. Front. Biosci. Landmark 2012, 17, 1266–1280. [Google Scholar] [CrossRef]
- Siva Rama Krishna, D.; Siddharthan, A.; Seshadri, S.K.; Sampath Kumar, T.S. A novel route for synthesis of nanocrystalline hydroxyapatite from eggshell waste. J. Mater. Sci. Mater. Med. 2007, 18, 1735–1743. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G. The Reserach on Relationship between Eggshell Color, Composition, Structure and Eggshell Quality. Ph.D. Thesis, Yangzhou University, Yangzhou, China, 2010. [Google Scholar] [CrossRef]
- Masuda, Y.; Hiramatsu, H. Bioavailability and Physiological Function of Eggshells and Eggshell Membranes. In Egg Bioscience and Biotechnology; Mine, Y., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2007; pp. 129–140. [Google Scholar] [CrossRef]
- Le Roy, N.; Stapane, L.; Gautron, J.; Hincke, M.T. Evolution of the Avian Eggshell Biomineralization Protein Toolkit—New Insights from Multi-Omics. Front. Genet. 2021, 12, 672433. [Google Scholar] [CrossRef] [PubMed]
- Le Roy, N.; Combes-Soia, L.; Brionne, A.; Labas, V.; Rodriguez-Navarro, A.B.; Hincke, M.T.; Nys, Y.; Gautron, J. Guinea fowl eggshell quantitative proteomics yield new findings related to its unique structural characteristics and superior mechanical properties. J. Proteom. 2019, 209, 103511. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Hernández, A.; Gomez-Morales, J.; Rodriguez-Navarro, A.B.; Gautron, J.; Nys, Y.; Garcia-Ruiz, J.M. Identification of Some Active Proteins in the Process of Hen Eggshell Formation. Cryst. Growth Des. 2008, 8, 4330–4339. [Google Scholar] [CrossRef]
- Fernandez, M.S.; Moya, A.; Lopez, L.; Arias, J.L. Secretion pattern, ultrastructural localization and function of extracellular matrix molecules involved in eggshell formation. Matrix Biol. 2001, 19, 793–803. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.M.H.; Rodriguez-Navarro, A.B.; Vidal, M.L.; Gautron, J.; Garcia-Ruiz, J.M.; Nys, Y. Changes in eggshell mechanical properties, crystallographic texture and in matrix proteins induced by moult in hens. Br. Poult. Sci. 2005, 46, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Ma, M.; Qu, L.; Shen, M.; Dou, T.; Wang, K. Estimation of genetic parameters related to eggshell strength using random regression models. Br. Poult. Sci. 2015, 56, 645–650. [Google Scholar] [CrossRef]
- Rodriguez-Navarro, A.; Kalin, O.; Nys, Y.; Garcia-Ruiz, J.M. Influence of the microstructure on the shell strength of eggs laid by hens of different ages. Br. Poult. Sci. 2002, 43, 395–403. [Google Scholar] [CrossRef]
- Radwan, L.M. Eggshell quality: A comparison between Fayoumi, Gimieizah and Brown Hy-Line strains for mechanical properties and ultrastructure of their eggshells. Anim. Prod. Sci. 2016, 56, 908–912. [Google Scholar] [CrossRef]
- Sun, C.J.; Chen, S.R.; Xu, G.Y.; Liu, X.M.; Yang, N. Global variation and uniformity of eggshell thickness for chicken eggs. Poult. Sci. 2012, 91, 2718–2721. [Google Scholar] [CrossRef] [PubMed]
- Benavides-Reyes, C.; Folegatti, E.; Dominguez-Gasca, N.; Litta, G.; Sanchez-Rodriguez, E.; Rodriguez-Navarro, A.B.; Faruk, M.U. Research Note: Changes in eggshell quality and microstructure related to hen age during a production cycle. Poult. Sci. 2021, 100, 101287. [Google Scholar] [CrossRef] [PubMed]
- Carnarius, K.M.; Conrad, K.M.; Mast, M.G.; Macneil, J.H. Relationship of eggshell ultrastructure and shell strength to the soundness of shell eggs. Poult. Sci. 1996, 75, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.; Tyler, C. The strength of guinea fowl (Numida meleagris) egg shells. Br. Poult. Sci. 1967, 7, 291–296. [Google Scholar] [CrossRef]
- Ar, A.; Rahn, H.; Paganelli, C.V. The Avian Egg: Mass and Strength. Condor 1979, 81, 331–337. [Google Scholar] [CrossRef]
- Panheleux, M.; Kalin, O.; Gautron, J.; Nys, Y. Features of eggshell formation in guinea fowl: Kinetics of shell deposition, uterine protein secretion and uterine histology. Br. Poult. Sci. 1999, 40, 632–643. [Google Scholar] [CrossRef]
- Pérez-Huerta, A.; Dauphin, Y. Comparison of the structure, crystallography and composition of eggshells of the guinea fowl and graylag goose. Zoology 2016, 119, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Dunn, I.C.; Rodriguez-Navarro, A.B.; Mcdade, K.; Schmutz, M.; Preisinger, R.; Waddington, D.; Wilson, P.W.; Bain, M.M. Genetic variation in eggshell crystal size and orientation is large and thesetraits are correlated with shell thickness and are associated with eggshell matrix protein markers. Anim. Genet. 2012, 43, 410–418. [Google Scholar] [CrossRef]
- Drabik, K.; Karwowska, M.; Wengerska, K.; Próchniak, T.; Adamczuk, A.; Batkowska, J. The Variability of Quality Traits of Table Eggs and Eggshell Mineral Composition Depending on Hens’ Breed and Eggshell Color. Animals 2021, 11, 1204. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.M.; Wang, Z.Y.; Lu, J. Study on the relationship between eggshell colors and egg quality as well as shell ultrastructure in Yangzhou chicken. Afr. J. Biotechnol. 2009, 8, 2898–2902. [Google Scholar]
- Arzate-Vázquez, I.; Mendez-Mendez, J.V.; Flores-Johnson, E.A.; Nicolas-Bermudez, J.; Chanona-Perez, J.J.; Santiago-Cortes, E. Study of the porosity of calcified chicken eggshell using atomic force microscopy and image processing. Micron 2019, 118, 50–57. [Google Scholar] [CrossRef]
- Tyler, C. Studies on egg shells. VI. —The distribution of pores in egg shells. J. Sci. Food Agric. 1955, 6, 170–176. [Google Scholar] [CrossRef]
- Board, R.G.; Scott, V.D. Porosity of the Avian Eggshell. Am. Zool. 1980, 20, 339–349. [Google Scholar] [CrossRef]
- Tullett, S.G. The porosity of avian eggshells. Comp. Biochem. Physiol. A Physiol. 1984, 78, 5–13. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, S.T.; Nie, F.Q.; Feng, L.; Zhu, G.S.; Jiang, L. Elaborate Architecture of the Hierarchical Hen’s Eggshell. Nano Res. 2011, 4, 171–179. [Google Scholar] [CrossRef]
- Tullett, S.G. Regulation of avian eggshell porosity. J. Zool. 1975, 177, 339–348. [Google Scholar] [CrossRef]
- Rosenberger, J.; Pytlak, K. Comparison of the eggshell mammillary microstructure of Galliformes and Anseriformes. J. Zool. 2024, 322, 340–350. [Google Scholar] [CrossRef]
- Fathi, M.B.; RahmatAbadi, Z.T. Eggshell microstructure, shell quality indices, mineralogy, and UV-Vis absorbance of domestic eggs of Iran. J. Photochem. Photobiol. 2024, 21, 100235. [Google Scholar] [CrossRef]
- Tian, Q.P.; Song, Z.H. Study on Improving Egg Fertilization Rate of Distant Hybridization Chicken. China Guizhou J. Anim. Husb. Vet. Med. 2012, 36, 7–8. [Google Scholar]
- Ning, Z.H. Breeding of New Supporting Lines for Grain-Saving Small Laying Hens and Research on Supporting Breeding Technology. Ph.D. Thesis, China Agricultural University, Beijing, China, 2004. [Google Scholar] [CrossRef]
- Feeding Standard for Chickens (NY/T33-2004). Available online: https://www.chinesestandard.net/PDF/English.aspx/NYT33-2004 (accessed on 9 May 2024).
- Idowu, O.M.O.; Ajuwon, R.O.; Oso, A.O.; Akinloye, O.A. Effects of Zinc Supplementation on Laying Performance, Serum Chemistry and Zn Residue in Tibia Bone, Liver, Excreta and Egg Shell of Laying Hens. Int. J. Poult. Sci. 2011, 10, 225–230. [Google Scholar] [CrossRef]
- Ren, H.-L.; Zhao, X.-Y.; Di, K.-Q.; Li, L.-H.; Hao, E.-Y.; Chen, H.; Zhou, R.-Y.; Nie, C.-S.; Wang, D.-H. Eggshell translucency in late-phase laying hens and its effect on egg quality and physiological indicators. Front. Vet. Sci. 2023, 10, 1133752. [Google Scholar] [CrossRef]
- Wang, D.H.; Li, Y.J.; Liu, L.; Liu, J.S.; Bao, M.; Yang, N.; Zhuo-Cheng, H.; Ning, Z.H. Traits of eggshells and shell membranes of translucent eggs. Poult. Sci. 2017, 96, 1514. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.E.; Legako, J.F. Traditional Methods for Mineral Analysis. In Food Analysis; Nielsen, S.S., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 371–386. [Google Scholar] [CrossRef]
- Liu, G.Y.; Shi, L.; Chen, Y.F.; Chen, H.; Zhang, C.; Wang, Y.T.; Ning, Z.H.; Wang, D.H. Estimation of genetic parameters of eggshell translucency and production traits in different genotypes of laying hens. Poult. Sci. 2023, 102, 102616. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.C.; Ning, Z.H.; Xu, G.Y.; Hou, Z.C.; Yang, N. Heritabilities and genetic and phenotypic correlations of egg quality traits in brown-egg dwarf layers. Poult. Sci. 2005, 84, 1209–1213. [Google Scholar] [CrossRef] [PubMed]
- Mohamadzamani, D.; Amini, S.; Seyed; Javidan, M.; Dehghan, S. Investigation of the Relationship Between Egg Shell Strength and Thickness Using Non-Destructive Ultrasound Method. J. Biosyst. Eng. 2022, 47, 263–269. [Google Scholar] [CrossRef]
- Ketta, M.; Tůmová, E. Relationship between eggshell thickness and other eggshell measurements in eggs from litter and cages. Ital. J. Anim. Sci. 2018, 17, 234–239. [Google Scholar] [CrossRef]
- Robinson, D.; King, N. The structure of the organic mammillary cores in some weak egg shells. Br. Poult. Sci. 1970, 11, 39–44. [Google Scholar] [CrossRef]
- Bunk, M.J.; Balloun, S. Ultrastructure of the mammillary region of low puncture strength avian eggshells. Poult. Sci. 1978, 57, 639–647. [Google Scholar] [CrossRef]
- Van Toledo, B.; Parsons, A.; Combs, G., Jr. Role of ultrastructure in determining eggshell strength. Poult. Sci. 1982, 61, 569–572. [Google Scholar] [CrossRef]
- Nys, Y.; Gautron, J.; Garcia-Ruiz, J.M.; Hincke, M.T. Avian eggshell mineralization: Biochemical and functional characterization of matrix proteins. Cr. Palevol. 2004, 3, 549–562. [Google Scholar] [CrossRef]
- Gautron, J.; Stapane, L.; Le Roy, N.; Nys, Y.; Rodriguez-Navarro, A.B.; Hincke, M.T. Avian eggshell biomineralization: An update on its structure, mineralogy and protein tool kit. BMC Mol. Cell Biol. 2021, 22, 11. [Google Scholar] [CrossRef] [PubMed]
- Tang, X. Study on the Characteristics and Variations of Goose Eggshell Structure and Components. Ph.D. Thesis, Yangzhou University, Yangzhou, China, 2022. [Google Scholar] [CrossRef]
- Rodríguez-Navarro, A.B.; Domínguez-Gasca, N.; Athanasiadou, D.; Le Roy, N.; González-Segura, A.; Reznikov, N.; Hincke, M.T.; McKee, M.D.; Checa, A.G.; Nys, Y.; et al. Guinea fowl eggshell structural analysis at different scales reveals how organic matrix induces microstructural shifts that enhance its mechanical properties. Acta Biomater. 2024, 178, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Navarro, A.B.; Marie, P.; Nys, Y.; Hincke, M.T.; Gautron, J. Amorphous calcium carbonate controls avian eggshell mineralization: A new paradigm for understanding rapid eggshell calcification. J. Struct. Biol. 2015, 190, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Damaziak, K.; Marzec, A.; Riedel, J.; Wojcik, W.; Pstrokonski, P.; Szudrowicz, H.; Gozdowski, D. Effect of pearl guinea fowl eggshell ultrastructure and microstructure on keets hatchability. Poult. Sci. 2023, 102, 102733. [Google Scholar] [CrossRef]
- Jonchère, V.; Brionne, A.; Gautron, J.; Nys, Y. Identification of uterine ion transporters for mineralisation precursors of the avian eggshell. BMC Physiol. 2012, 12, 10. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.E.; Böhm, C.F.; Harris, J.; Demmert, B.; Jacob, D.E.; Mondeshki, M.; Ruiz-Agudo, E.; Rodríguez-Navarro, C. Nonclassical crystallization in vivo et in vitro (I): Process-structure-property relationships of nanogranular biominerals. J. Struct. Biol. 2016, 196, 244–259. [Google Scholar] [CrossRef]
- Ahmad, S.; Hashim, J.; Ghazali, M. The effects of porosity on mechanical properties of cast discontinuous reinforced metal–matrix composite. J. Compos. Mater. 2005, 39, 451–466. [Google Scholar] [CrossRef]
- Sun, C.; Xu, G.; Yang, N. Differential label-free quantitative proteomic analysis of avian eggshell matrix and uterine fluid proteins associated with eggshell mechanical property. Proteomics 2013, 13, 3523–3536. [Google Scholar] [CrossRef]
- Marie, P.; Labas, V.; Brionne, A.; Harichaux, G.; Hennequet-Antier, C.; Nys, Y.; Gautron, J. Quantitative proteomics and bioinformatic analysis provide new insight into protein function during avian eggshell biomineralization. J. Proteom. 2015, 113, 178–193. [Google Scholar] [CrossRef] [PubMed]
- Mann, K.; Macek, B.; Olsen, J.V. Proteomic analysis of the acid-soluble organic matrix of the chicken calcified eggshell layer. Proteomics 2006, 6, 3801–3810. [Google Scholar] [CrossRef] [PubMed]
- Marie, P.; Labas, V.; Brionne, A.; Harichaux, G.; Hennequet-Antier, C.; Rodriguez-Navarro, A.B.; Nys, Y.; Gautron, J. Quantitative proteomics provides new insights into chicken eggshell matrix protein functions during the primary events of mineralisation and the active calcification phase. J. Proteom. 2015, 126, 140–154. [Google Scholar] [CrossRef] [PubMed]
- Dennis, J.E.; Xiao, S.Q.; Agarwal, M.; Fink, D.J.; Heuer, A.H.; Caplan, A.I. Microstructure of matrix and mineral components of eggshells from white leghorn chickens (Gallus gallus). J. Morphol. 1996, 228, 287–306. [Google Scholar] [CrossRef]
- Cusack, M.; Fraser, A.C.; Stachel, T. Magnesium and phosphorus distribution in the avian eggshell. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2003, 134, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.; Sun, C.; Shen, M.; Wang, K.; Yang, N.; Zheng, J.; Xu, G. Genetic architecture dissection by genome-wide association analysis reveals avian eggshell ultrastructure traits. Sci. Rep. 2016, 6, 28836. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Traits | Dwarf Layer-White | Guinea Fowl |
|---|---|---|
| ELL (cm) | 5.48 ± 0.19 a | 5.30 ± 0.21 b |
| ESL (cm) | 4.19 ± 0.12 a | 4.05 ± 0.13 b |
| ESI | 1.31 ± 0.47 | 1.31 ± 0.40 |
| ESA (cm2) | 65.80 ± 3.08 a | 62.82 ± 3.02 b |
| EW (g) | 52.76 ± 3.71 a | 49.22 ± 3.57 b |
| ESS (kg/cm2) | 3.38 ± 0.78 b | 9.70 ± 3.28 a |
| SUW | 0.06 ± 0.01 b | 0.20 ± 0.07 a |
| ESMT (μm) | 11.66 ± 7.44 b | 16.94 ± 10.66 a |
| EST (μm) | 302.09 ± 54.92 b | 458.78 ± 55.97 a |
| ESR (%) | 12.73 ± 1.66 b | 17.93 ± 1.63 a |
| Traits | Dwarf Layer-White | Guinea Fowl |
|---|---|---|
| MAN | 787.03 ± 93.32 | 801.83 ± 115.54 |
| MTR (%) | 41.99 ± 3.81 a | 32.72 ± 3.36 b |
| ETR (%) | 58.01 ± 3.81 b | 67.28 ± 3.36 a |
| QBPs | 687.51 ± 79.42 a | 20.79 ± 11.51 b |
| AABPs (μm2) | 0.05 ± 0.01 b | 0.15 ± 0.09 a |
| ASBPs (μm2) | 37.15 ± 5.92 a | 1.54 ± 0.42 b |
| ARBE (%) | 3.23 ± 0.47 a | 0.14 ± 0.04 b |
| APBPs (μm) | 0.84 ± 0.04 b | 1.09 ± 0.51 a |
| Traits | Mean Value of 3 Eggshell Layers | Mean Value of 3 Eggshell Ends | |||||
|---|---|---|---|---|---|---|---|
| ML | PL | VCL | Blunt End | Middle End | Sharp End | ||
| Dwarf Layer-White | QBPs | 532.06 ± 151.20 c | 1236.55 ± 233.77 a | 819.35 ± 122.37 b | 840.29 ± 311.40 | 905.08 ± 410.53 | 798.03 ± 320.55 |
| AABPs (μm2) | 0.058 ± 0.011 b | 0.067 ± 0.011 a | 0.064 ± 0.104 ab | 0.068 ± 0.019 | 0.060 ± 0.017 | 0.062 ± 0.021 | |
| ASBPs (μm2) | 31.93 ± 14.24 c | 82.31 ± 14.00 a | 53.00 ± 18.73 b | 61.51 ± 27.54 | 51.36 ± 24.53 | 50.53 ± 25.33 | |
| ARBE (%) | 2.89 ± 1.39 c | 7.20 ± 1.23 a | 4.64 ± 1.64 b | 5.38 ± 2.41 | 4.50 ± 2.15 | 4.42 ± 2.22 | |
| APBPs (μm) | 0.90 ± 0.09 | 0.96 ± 0.13 | 0.95 ± 0.12 | 0.99 ± 0.18 | 0.91 ± 0.17 | 0.90 ± 0.28 | |
| Guinea fowl | QBPs | 5.96 ± 3.05 b | 19.59 ± 10.48 a | 14.73 ± 18.93 a | 15.32 ± 9.49 | 12.64 ± 6.38 | 11.1 ± 6.87 |
| AABPs (μm2) | 0.086 ± 0.052 b | 0.058 ± 0.046 b | 0.122 ± 0.064 a | 0.096 ± 0.076 | 0.079 ± 0.055 | 0.106 ± 0.070 | |
| ASBPs (μm2) | 0.49 ± 0.30 b | 1.25 ± 0.69 a | 1.24 ± 0.55 a | 1.13 ± 0.64 | 0.92 ± 0.75 | 0.94 ± 0.65 | |
| ARBE (%) | 0.06 ± 0.04 b | 0.11 ± 0.06 a | 0.12 ± 0.06 a | 0.12 ± 0.09 | 0.08 ± 0.64 | 0.09 ± 0.08 | |
| APBPs (μm) | 0.98 ± 0.31 | 0.83 ± 0.40 | 0.98 ± 0.33 | 0.91 ± 0.53 | 0.84 ± 0.39 | 1.09 ± 0.48 | |
| Traits | Dwarf Layer-White | Guinea Fowl |
|---|---|---|
| Organic matter | 3.15 ± 0.20 B | 4.10 ± 0.51 A |
| Inorganic matter | 96.85 ± 0.20 A | 95.90 ± 0.51 B |
| Ca | 32.45 ± 1.83 b | 34.52 ± 0.58 a |
| P | 0.12 ± 0.02 A | 0.07 ± 0.02 B |
| Mg | 0.75 ± 0.21 | 0.63 ± 0.12 |
| Traits 2 | ESS | EST | ESMT | MT | ET | MAN | QBPs | AABPs | ASBPs | ARBE |
|---|---|---|---|---|---|---|---|---|---|---|
| ESS | 1 | 0.783 ** | 0.124 | 0.173 | 0.548 ** | 0.111 | 0.131 | 0.258 | 0.188 | 0.246 |
| EST | 0.652 ** | 1 | 0.140 | 0.769 ** | 0.911 ** | −0.053 | 0.216 | 0.237 | 0.206 | 0.199 |
| ESMT | 0.161 | 0.090 | 1 | −0.135 | −0.104 | 0.063 | 0.240 | −0.299 | −0.131 | 0.022 |
| MT | 0.286 | 0.611 ** | −0.008 | 1 | 0.257 | −0.423 ** | 0.041 | 0.325 | 0.313 | 0.345 |
| ET | 0.534 * | 0.829 ** | −0.015 | 0.347 | 1 | −0.030 | 0.042 | 0.113 | 0.040 | 0.024 |
| MAN | 0.175 | −0.171 | 0.094 | −0.551 ** | 0.019 | 1 | 0.356 * | −0.463 * | 0.592 * | 0.465 * |
| QBPs | −0.455 * | −0.210 | −0.111 | −0.074 | −0.204 | −0.300 | 1 | −0.348 * | 0.655 ** | 0.529 ** |
| AABPs | 0.241 | 0.225 | −0.195 | 0.049 | 0.260 | 0.363 | −0.629 ** | 1 | 0.478 * | 0.284 |
| ASBPs | −0.084 | 0.146 | −0.078 | 0.103 | 0.329 | −0.357 | 0.501 * | 0.439 * | 1 | 0.712 ** |
| ARBE | −0.256 | −0.359 | −0.269 | −0.203 | −0.143 | 0.196 | 0.417 * | 0.308 | 0.398 * | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-T.; Chen, Y.-F.; Zhang, J.-J.; Zhang, Q.; Zhao, X.-Y.; Zhou, R.-Y.; Chen, H.; Wang, D.-H. Comparative Analysis of the Ultrastructure, Bubble Pores, and Composition of Eggshells of Dwarf Layer-White and Guinea Fowl. Animals 2024, 14, 1496. https://doi.org/10.3390/ani14101496
Wang Y-T, Chen Y-F, Zhang J-J, Zhang Q, Zhao X-Y, Zhou R-Y, Chen H, Wang D-H. Comparative Analysis of the Ultrastructure, Bubble Pores, and Composition of Eggshells of Dwarf Layer-White and Guinea Fowl. Animals. 2024; 14(10):1496. https://doi.org/10.3390/ani14101496
Chicago/Turabian StyleWang, Yi-Tong, Yi-Fan Chen, Jun-Jie Zhang, Quan Zhang, Xiao-Yu Zhao, Rong-Yan Zhou, Hui Chen, and De-He Wang. 2024. "Comparative Analysis of the Ultrastructure, Bubble Pores, and Composition of Eggshells of Dwarf Layer-White and Guinea Fowl" Animals 14, no. 10: 1496. https://doi.org/10.3390/ani14101496
APA StyleWang, Y.-T., Chen, Y.-F., Zhang, J.-J., Zhang, Q., Zhao, X.-Y., Zhou, R.-Y., Chen, H., & Wang, D.-H. (2024). Comparative Analysis of the Ultrastructure, Bubble Pores, and Composition of Eggshells of Dwarf Layer-White and Guinea Fowl. Animals, 14(10), 1496. https://doi.org/10.3390/ani14101496