

Positive In Vitro Effect of ROCK Pathway Inhibitor Y-27632 on Qualitative Characteristics of Goat Sperm Stored at Low Temperatures
 , , , , , , and
, , , , , , and
Abstract
Simple Summary
Abstract
1. Introduction
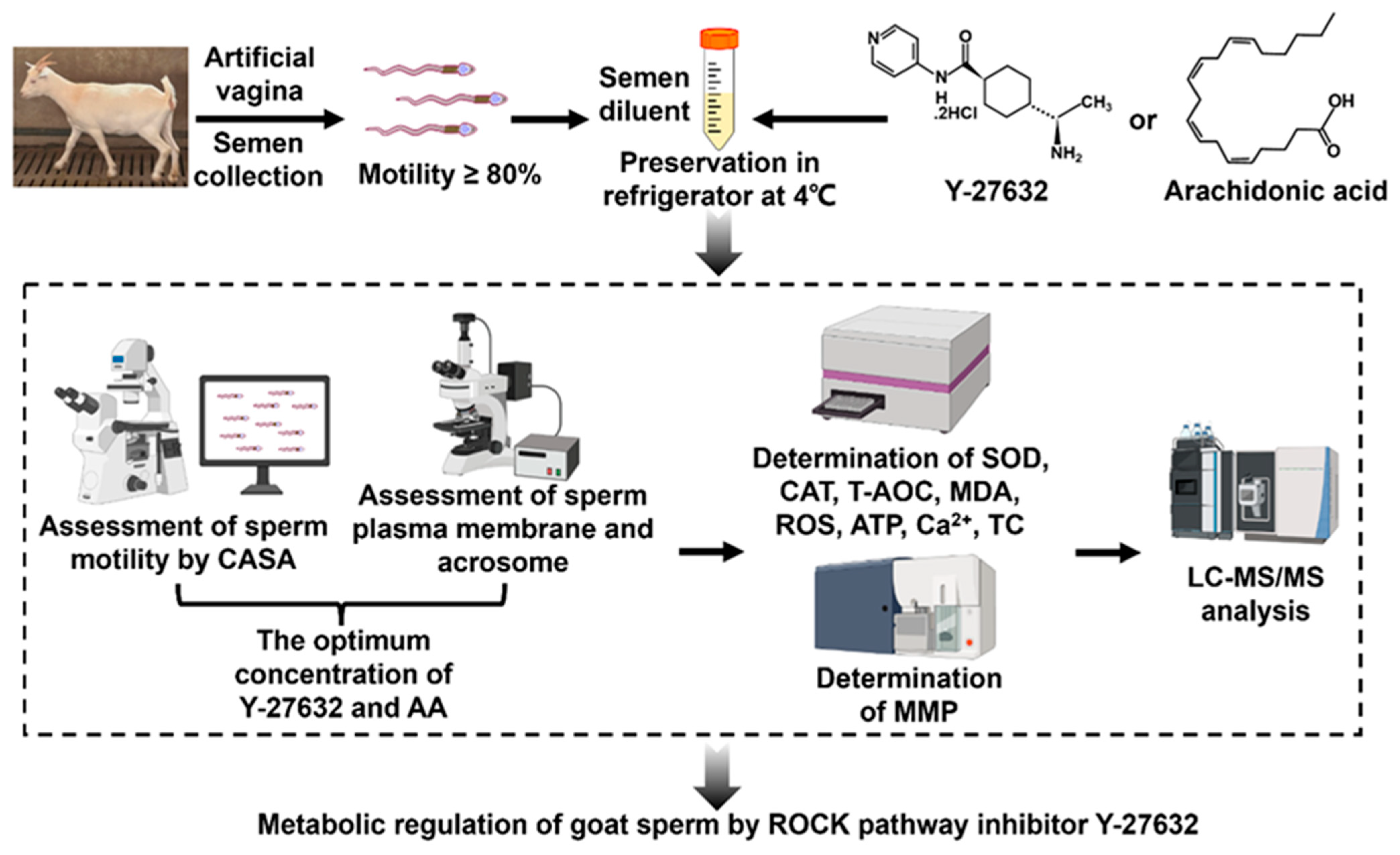
2. Material and Methods
2.1. Ethics Statement
2.2. Dilution Preparation
2.3. Semen Processing
2.4. Analysis of Sperm Motility
2.5. Evaluation of Sperm Plasma Membrane Integrity Rate
2.6. Detection of Sperm Acrosome Integrity Rate
2.7. Determination of T-AOC, SOD, CAT Activity, and MDA Content
2.8. Determination of ROS Levels
2.9. Determination of Mitochondrial Membrane Potential
2.10. Determination of TC and Ca2+ Content in Sperm
2.11. Mass Spectrometric Analysis
Metabolite Extraction
2.12. LC-MS/MS Analysis
2.13. Data Analyses Processing
2.14. Artificial Insemination
2.15. Statistical Analysis
3. Results
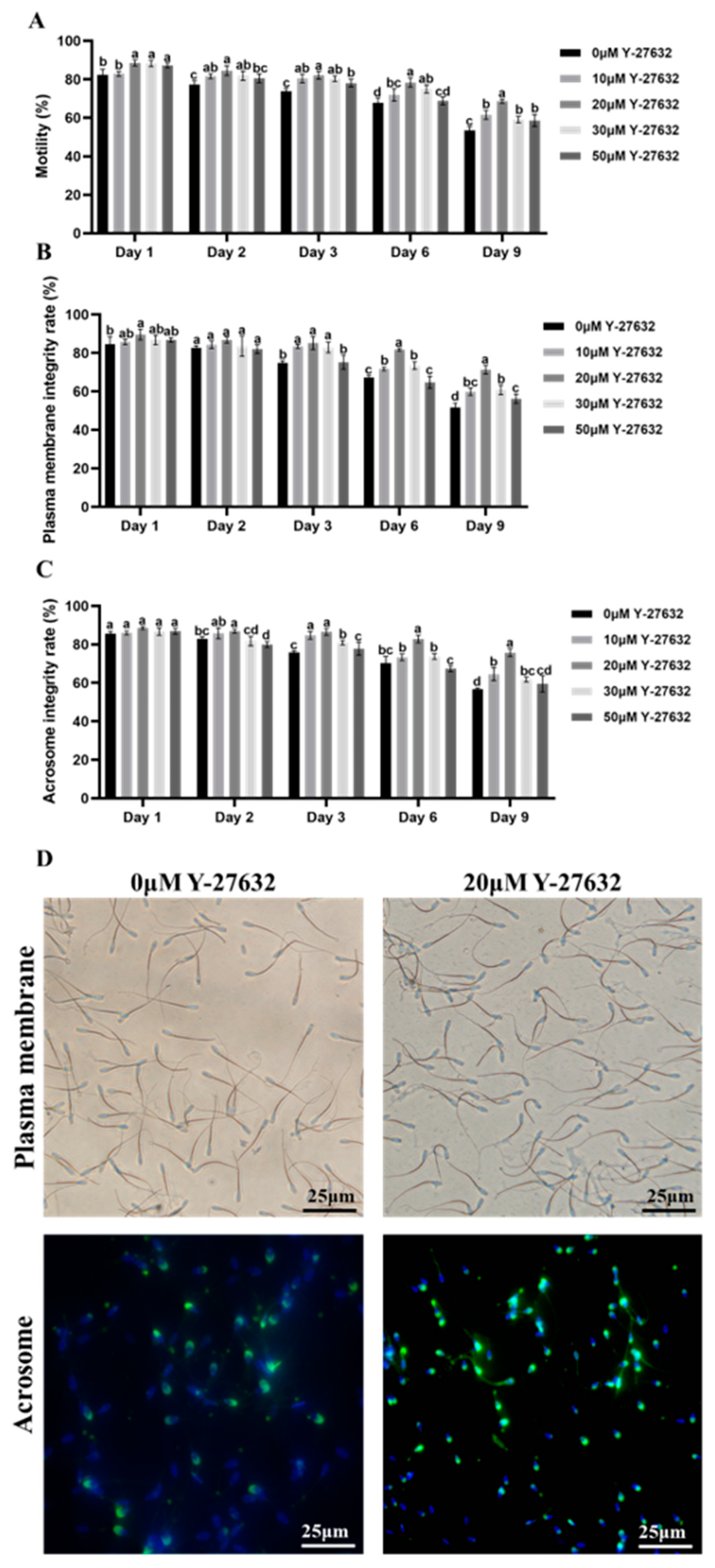
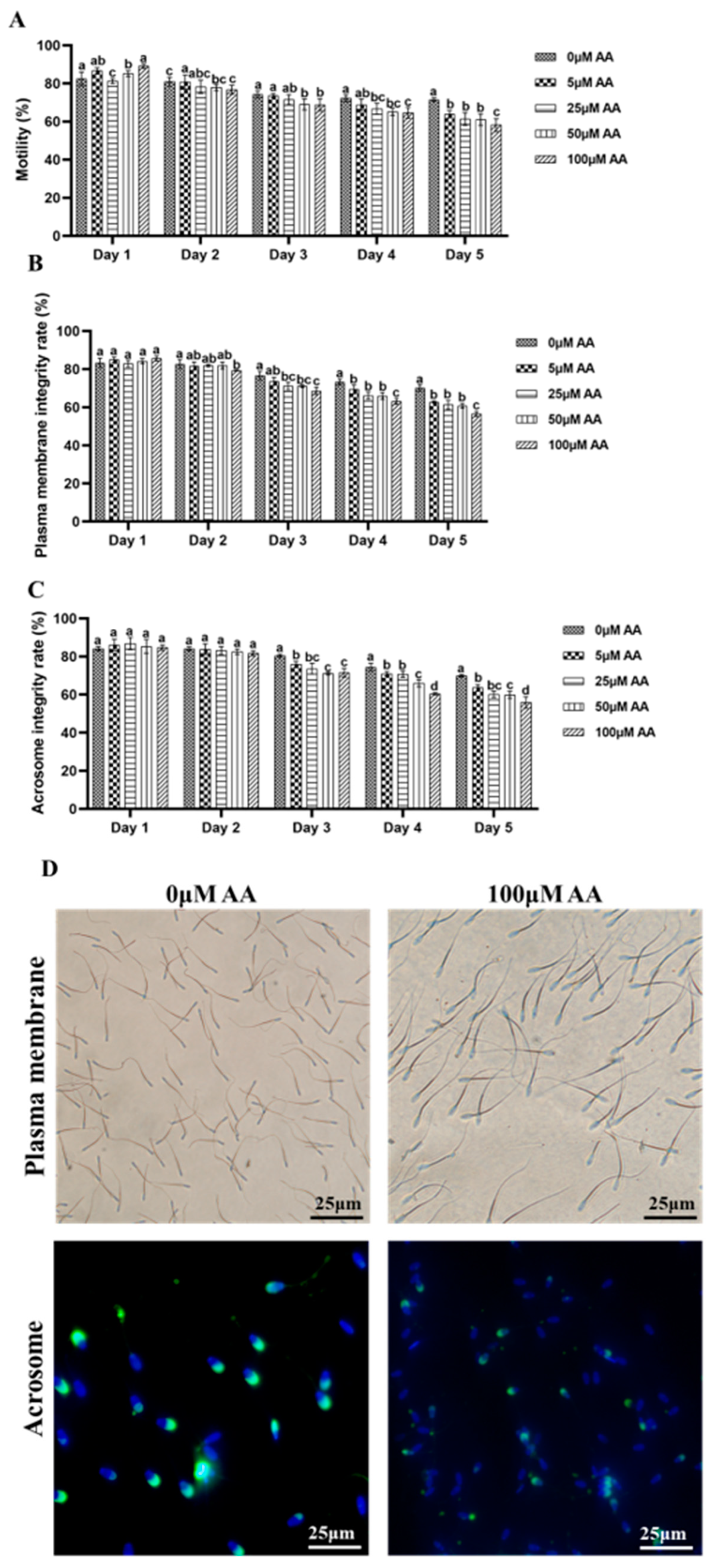
3.1. Effects of Y-27632 and AA on the Quality of Goat Sperm
3.2. Analysis of Y-27632 and AA on Antioxidant Ability of Goat Sperm
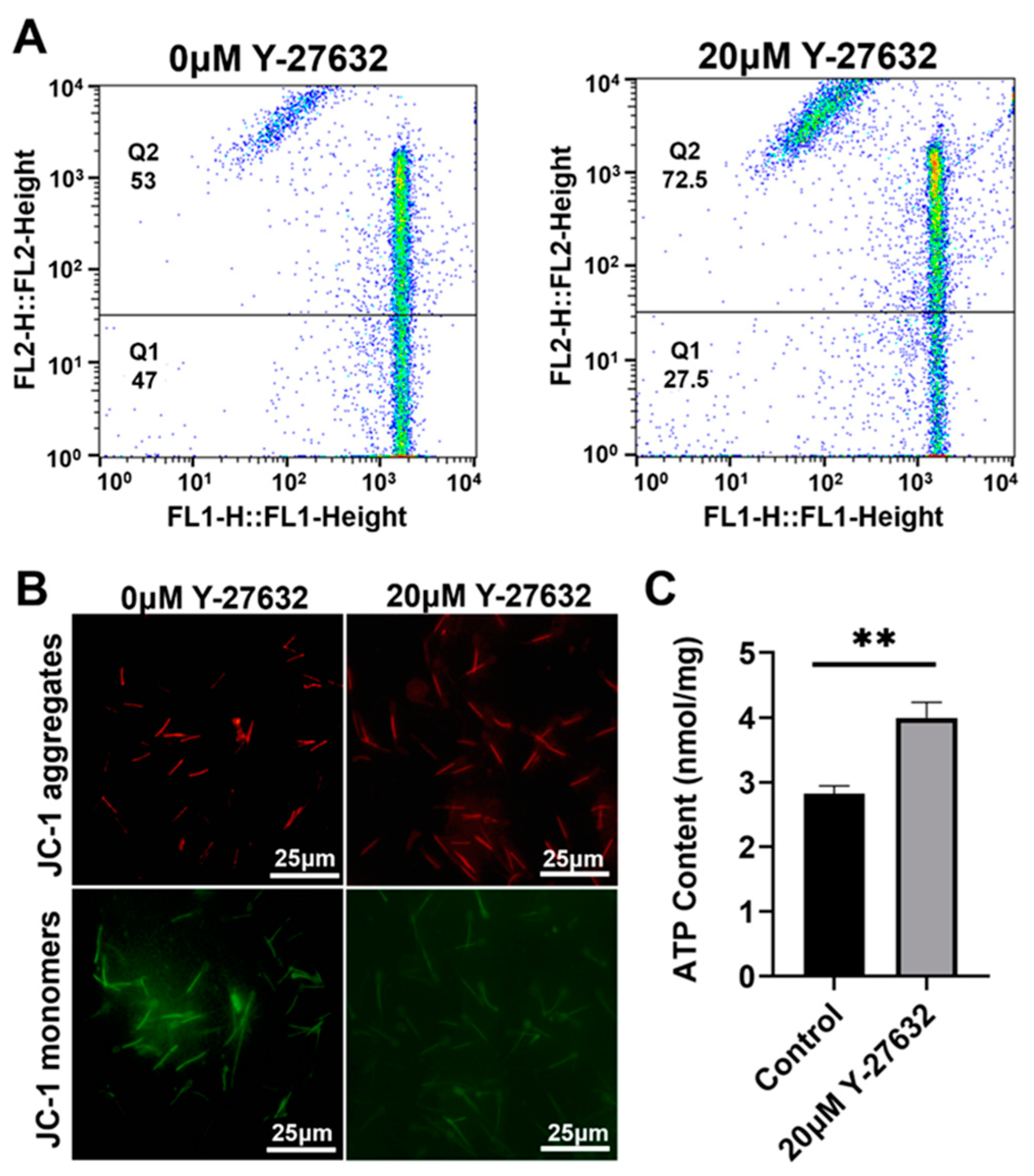
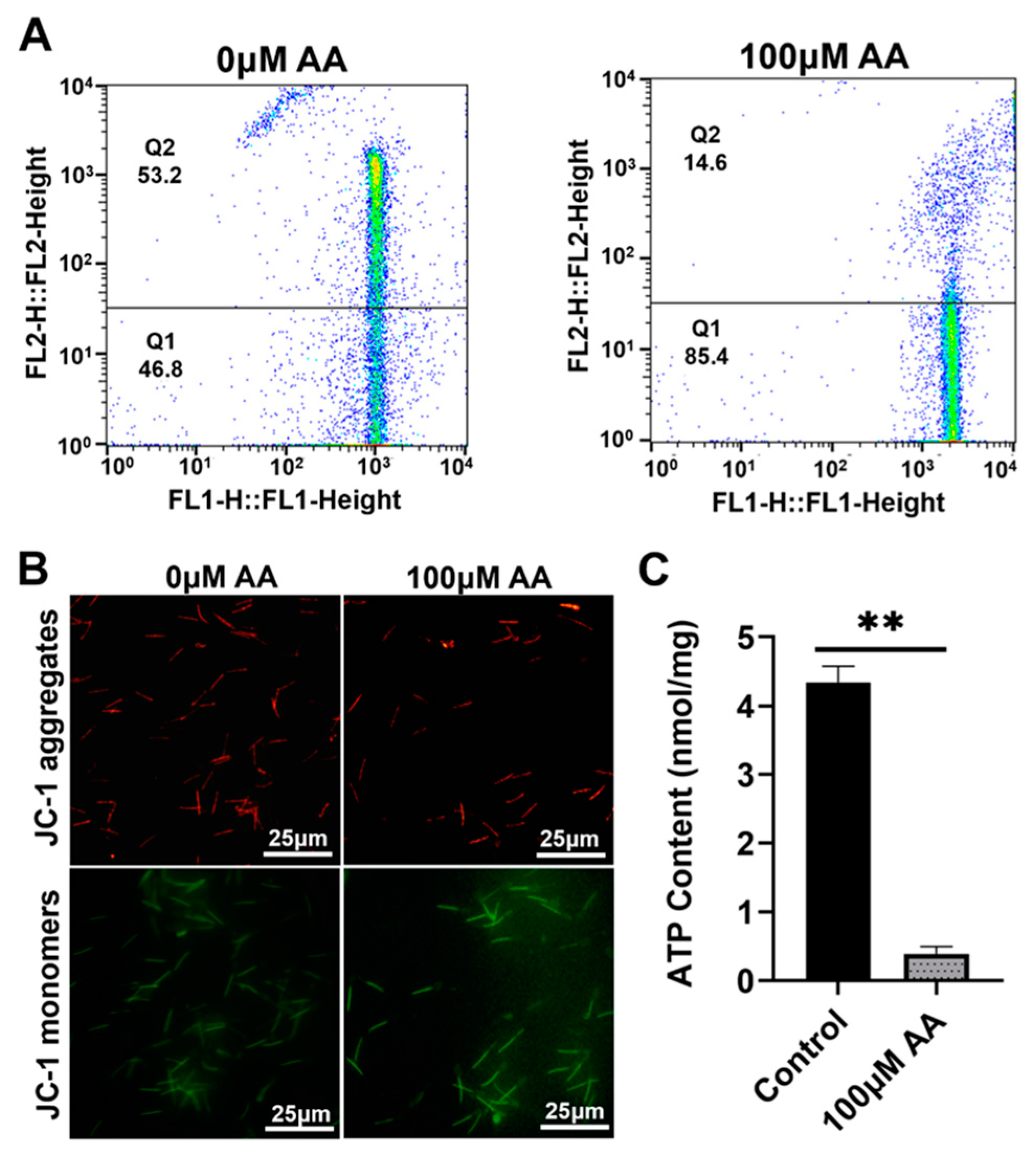
3.3. Effects of Y-27632 and AA on Mitochondrial Energy Metabolism in Goat Sperm
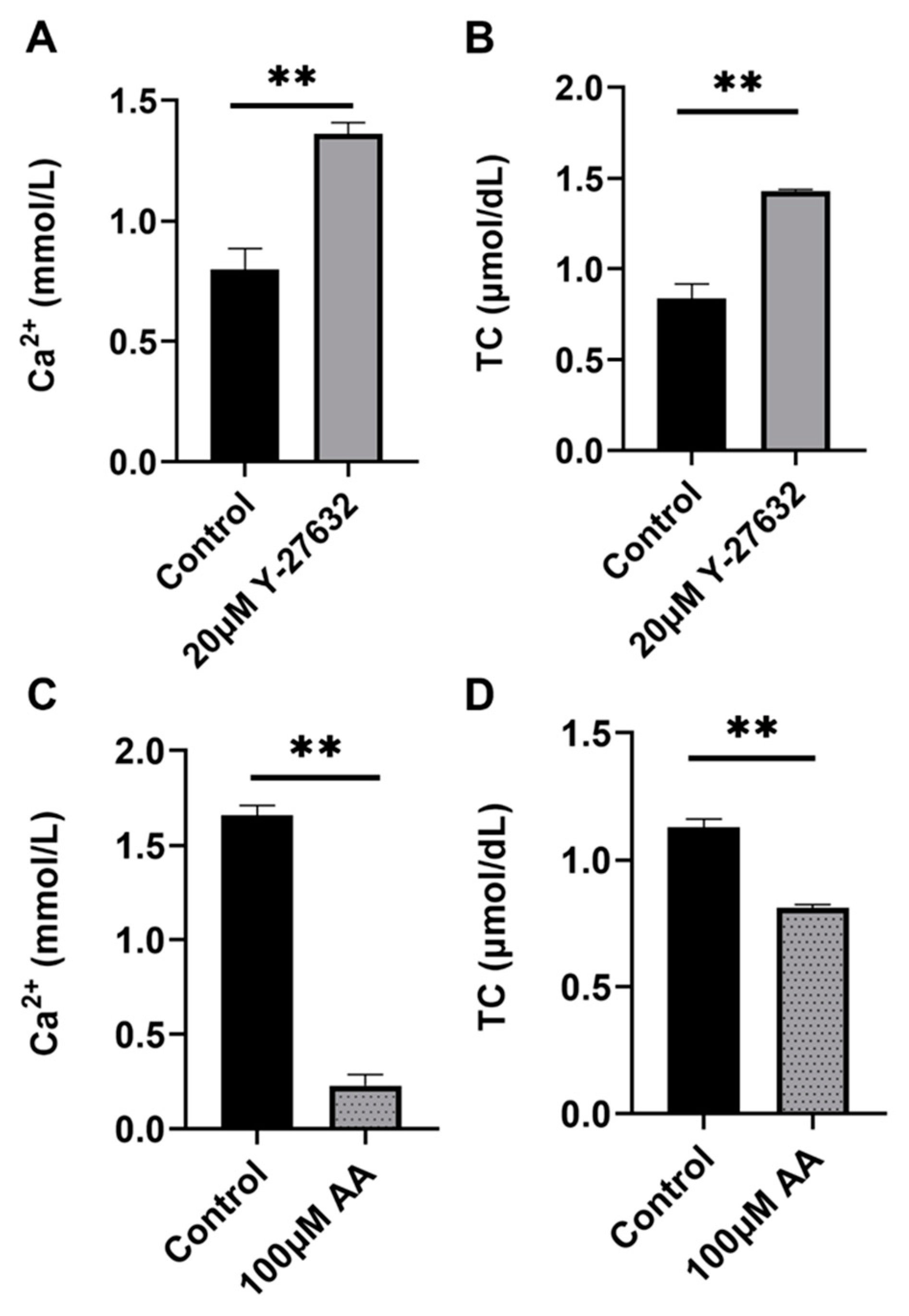
3.4. Effects of Y-27632 (Day 9) and AA (Day 5) on Ca2+ and TC in Goat Sperm
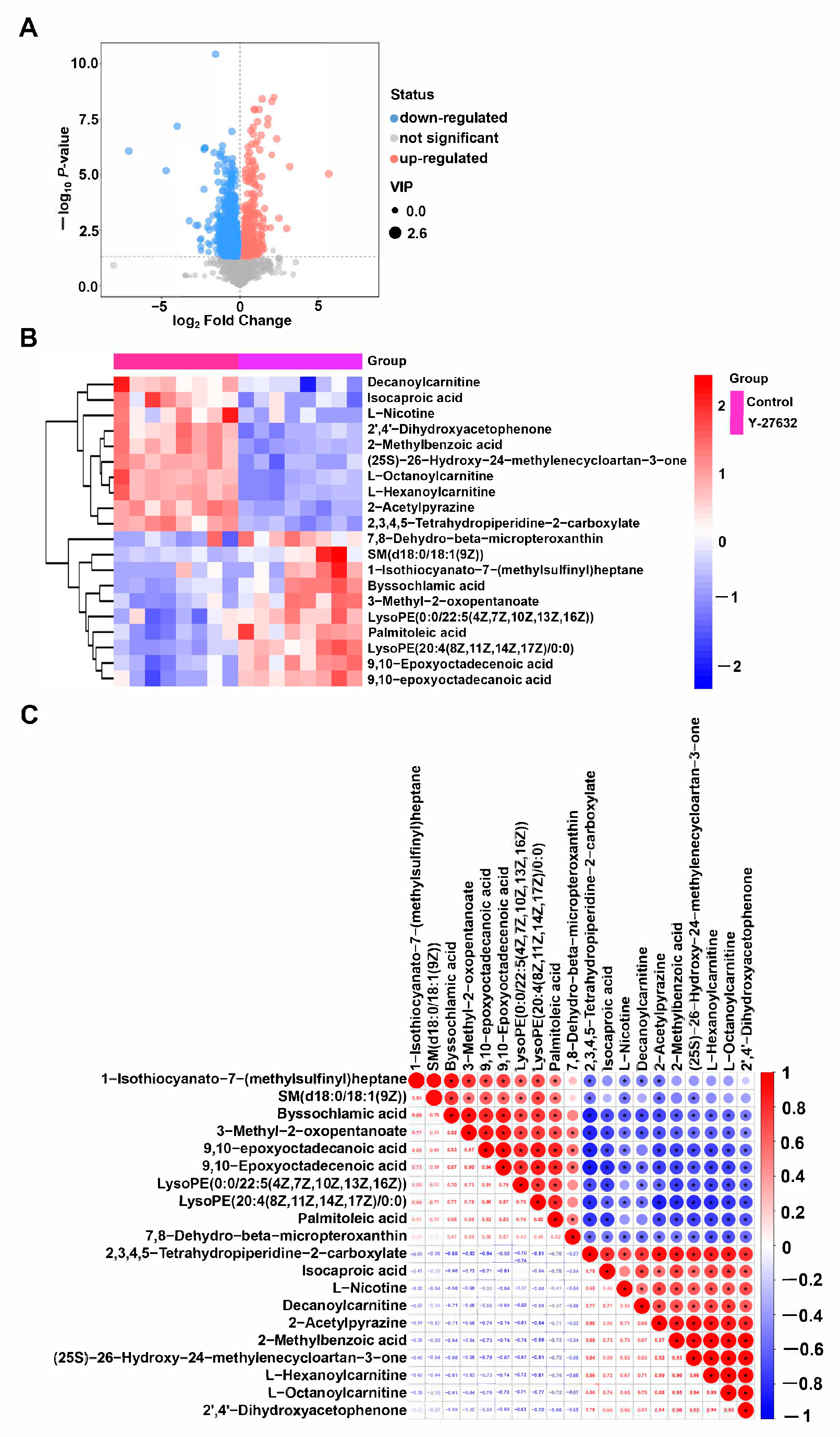
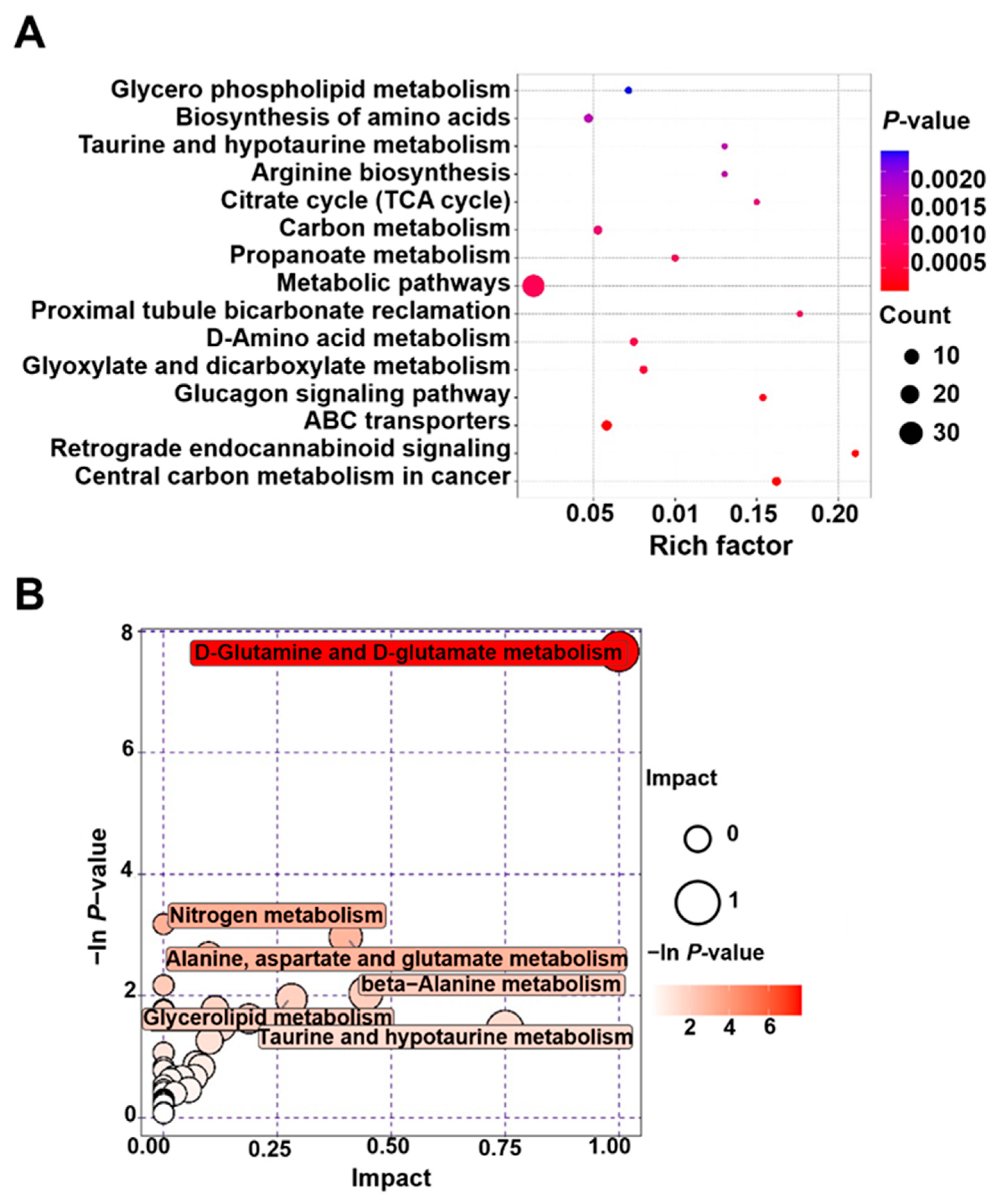
3.5. Metabolomics Analysis of Y-27632 Goat Sperm
4. Fertility Analysis
5. Discussion
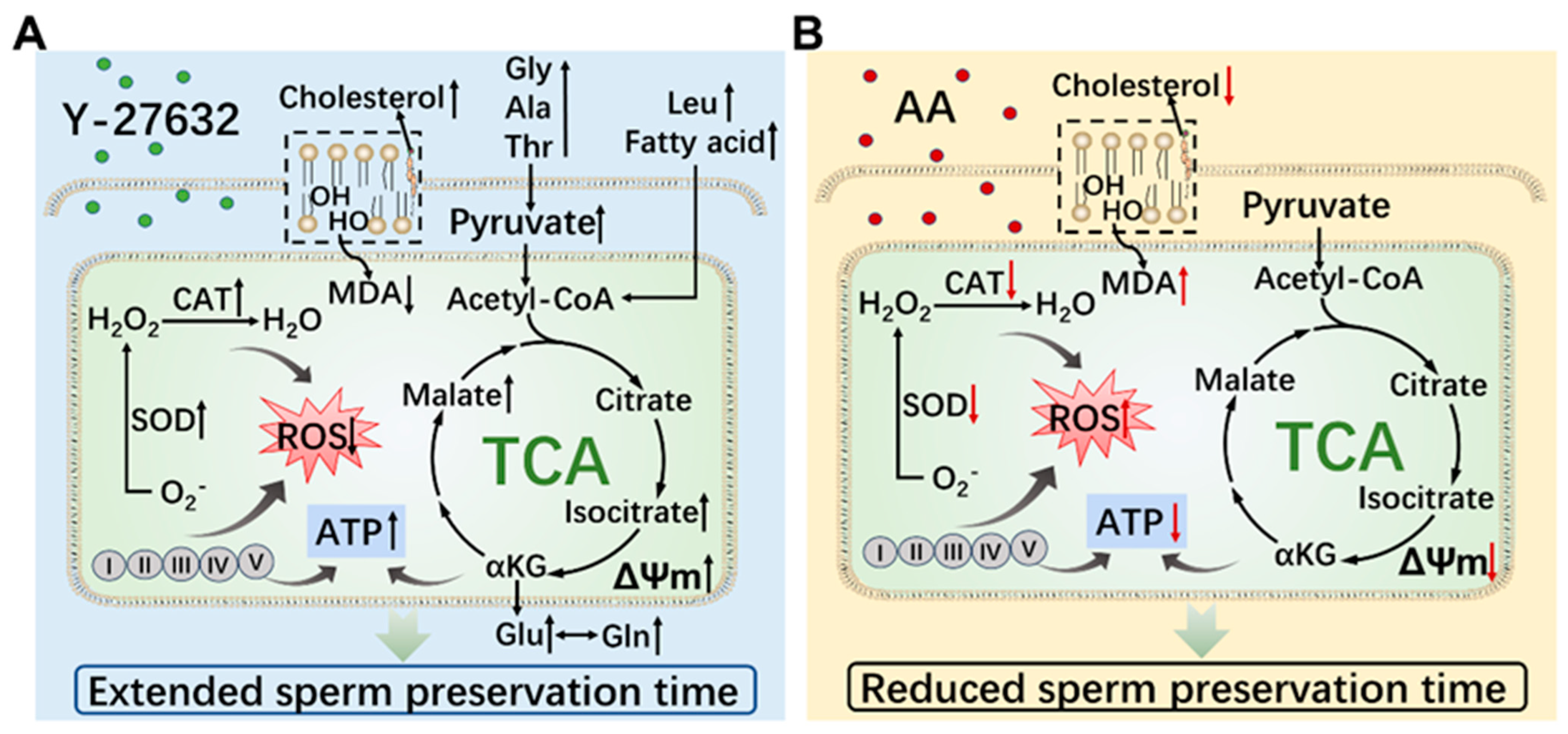
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, Z.; He, W.; Leung, T.C.N.; Chung, H.Y. Immortalization and Characterization of Rat Lingual Keratinocytes in a High-Calcium and Feeder-Free Culture System Using ROCK Inhibitor Y-27632. Int. J. Mol. Sci. 2021, 22, 6782. [Google Scholar] [CrossRef]
- Guan, G.; Cannon, R.D.; Coates, D.E.; Mei, L. Effect of the Rho-Kinase/ROCK Signaling Pathway on Cytoskeleton Components. Genes 2023, 14, 272. [Google Scholar] [CrossRef]
- Reynolds, S.D.; Rios, C.; Wesolowska-Andersen, A.; Zhuang, Y.; Pinter, M.; Happoldt, C.; Hill, C.L.; Lallier, S.W.; Cosgrove, G.P.; Solomon, G.M.; et al. Airway Progenitor Clone Formation Is Enhanced by Y-27632–Dependent Changes in the Transcriptome. Am. J. Respir. Cell Mol. Biol. 2016, 55, 323–336. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-C.; Liu, H.-F.; Hsu, K.-C.; Yang, J.-M.; Chen, C.; Liu, K.-K.; Hsu, T.-S.; Chao, J.-I. 7-Chloro-6-piperidin-1-yl-quinoline-5,8-dione (PT-262), a novel ROCK inhibitor blocks cytoskeleton function and cell migration. Biochem. Pharmacol. 2011, 81, 856–865. [Google Scholar] [CrossRef] [PubMed]
- Heng, N.; Zhao, Z.-X.; Guo, Y.; Gao, S.; Cai, D.-L.; Fu, B.-F.; Sheng, X.-H.; Wang, X.-G.; Xing, K.; Xiao, L.-F.; et al. RhoA improves cryopreservation of rooster sperm through the Rho/RhoA-associated kinase/cofilin pathway. Poult. Sci. 2022, 101, 102051. [Google Scholar] [CrossRef]
- Feng, Z.-H.; Zhang, X.-H.; Wang, X.-Y. The ROCK pathway inhibitor Y-27632 mitigates hypoxia and oxidative stress-induced injury to retinal Müller cells. Neural Regen. Res. 2018, 13, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Zepeta-Flores, N.; Valverde, M.; Lopez-Saavedra, A.; Rojas, E. Glutathione depletion triggers actin cytoskeleton changes via actin-binding proteins. Genet. Mol. Biol. 2018, 41, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Zhang, C.; Yu, S.; Liu, S.; Wang, G.; Lan, H.; Zheng, X.; Li, S. Glycine ameliorates MBP-induced meiotic abnormalities and apoptosis by regulating mitochondrial-endoplasmic reticulum interactions in porcine oocytes. Environ. Pollut. 2022, 309, 119756. [Google Scholar] [CrossRef]
- Pierozan, P.; Biasibetti, H.; Schmitz, F.; Ávila, H.; Fernandes, C.G.; Pessoa-Pureur, R.; Wyse, A.T. Neurotoxicity of Methylmercury in Isolated Astrocytes and Neurons: The Cytoskeleton as a Main Target. Mol. Neurobiol. 2017, 54, 5752–5767. [Google Scholar] [CrossRef]
- Noiles, E.E.; Thompson, K.A.; Storey, B.T. Water permeability, Lp, of the mouse sperm plasma membrane and its activation energy are strongly dependent on interaction of the plasma membrane with the sperm cytoskeleton. Cryobiology 1997, 35, 79–92. [Google Scholar] [CrossRef]
- Osman, R.; Lee, S.; Almubarak, A.; Han, J.I.; Yu, I.J.; Jeon, Y. Antioxidant Effects of Myo-Inositol Improve the Function and Fertility of Cryopreserved Boar Semen. Antioxidants 2023, 12, 1673. [Google Scholar] [CrossRef] [PubMed]
- Shan, M.-M.; Sun, S.-C. The multiple roles of RAB GTPases in female and male meiosis. Hum. Reprod. Updat. 2021, 27, 1013–1029. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Dewry, R.; Srivastava, R.; Nath, S.; Mohanty, T. Targeted antioxidant delivery modulates mitochondrial functions, ameliorates oxidative stress and preserve sperm quality during cryopreservation. Theriogenology 2022, 179, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Dunleavy, J.E.M.; O’bryan, M.K.; Stanton, P.G.; O’donnell, L. The cytoskeleton in spermatogenesis. Reproduction 2019, 157, R53–R72. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.C.; Ray, D.M.; Lackford, B.; Rubino, M.; Olden, K.; Roberts, J.D. Arachidonic acid stimulates cell adhesion through a novel p38 MAPK-RhoA signaling pathway that involves heat shock protein 27. J. Biol. Chem. 2009, 284, 20936–20945. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ren, C.; Chen, Y.; Wang, M.; Tang, J.; Zhang, Y.; Wang, Q.; Zhang, Z. Changes on proteomic and metabolomic profiling of cryopreserved sperm effected by melatonin. J. Proteom. 2023, 273, 104791. [Google Scholar] [CrossRef]
- Bezerra, L.G.; Silva, A.M.; Dantas, M.R.; Santos, R.P.; Moreira, S.S.; Pereira, A.G.; Oliveira, M.F.; Comizzoli, P.; Silva, A.R. Development of assays for the characterization of sperm motility parameters, viability, and membrane integrity in the epididymis and vas deferens of the greater rhea (Rhea americana). Anim. Reprod. 2023, 20, e20230113. [Google Scholar] [CrossRef]
- Bai, H.; Zhang, Z.; Shen, W.; Fu, Y.; Cao, Z.; Liu, Z.; Yang, C.; Sun, S.; Wang, L.; Ling, Y.; et al. Metabolomics Analysis of Sodium Salicylate Improving the Preservation Quality of Ram Sperm. Molecules 2023, 29, 188. [Google Scholar] [CrossRef]
- Valcarce, D.G.; Robles, V. Evaluation of intracellular location of reactive oxygen species in Solea Senegalensis Spermatozoa. JoVE J. Vis. Exp. 2018, e55323. [Google Scholar] [CrossRef]
- D’Ovidio, C.; Locatelli, M.; Perrucci, M.; Ciriolo, L.; Furton, K.G.; Gazioglu, I.; Kabir, A.; Merone, G.M.; de Grazia, U.; Ali, I.; et al. LC-MS/MS Application in Pharmacotoxicological Field: Current State and New Applications. Molecules 2023, 28, 2127. [Google Scholar] [CrossRef]
- Ai, J.; Wu, Q.; Battino, M.; Bai, W.; Tian, L. Using untargeted metabolomics to profile the changes in roselle (Hibiscus sabdariffa L.) anthocyanins during wine fermentation. Food Chem. 2021, 364, 130425. [Google Scholar] [CrossRef] [PubMed]
- Antonouli, S.; Di Nisio, V.; Messini, C.; Samara, M.; Salumets, A.; Daponte, A.; Anifandis, G. Sperm plasma membrane ion transporters and male fertility potential: A perspective under the prism of cryopreservation. Cryobiology 2024, 114, 104845. [Google Scholar] [CrossRef] [PubMed]
- Inanc, M.E.; Güngör, Ş.; Avdatek, F.; Yeni, D.; Gülhan, M.F.; Olğaç, K.T.; Denk, B.; Taşdemir, U. Thymoquinone improves motility, plasma membrane integrity and DNA integrity of frozen–thawed ram semen. Andrologia 2022, 54, e14547. [Google Scholar] [CrossRef] [PubMed]
- Khawar, M.B.; Gao, H.; Li, W. Mechanism of Acrosome Biogenesis in Mammals. Front. Cell Dev. Biol. 2019, 7, 195. [Google Scholar] [CrossRef] [PubMed]
- Karaşör, F.; Bucak, M.N.; Cenariu, M.; Bodu, M.; Taşpınar, M.; Taşpınar, F. The Effects of Different Doses of ROCK Inhibitor, Antifreeze Protein III, and Boron Added to Semen Extender on Semen Freezeability of Ankara Bucks. Molecules 2022, 27, 8070. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Majzoub, A.; Agarwal, A. Oxidative stress and sperm function: A systematic review on evaluation and management. Arab. J. Urol. 2019, 17, 87–97. [Google Scholar] [CrossRef]
- Kowalczyk, A. The Role of the Natural Antioxidant Mechanism in Sperm Cells. Reprod. Sci. 2022, 29, 1387–1394. [Google Scholar] [CrossRef]
- Yan, L.; Liu, J.; Wu, S.; Zhang, S.; Ji, G.; Gu, A. Seminal superoxide dismutase activity and its relationship with semen quality and SOD gene polymorphism. J. Assist. Reprod. Genet. 2014, 31, 549–554. [Google Scholar] [CrossRef]
- Agarwal, A.; Durairajanayagam, D.; Du Plessis, S.S. Utility of antioxidants during assisted reproductive techniques: An evidence based review. Reprod. Biol. Endocrinol. 2014, 12, 112. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Niederberger, C. Re: Mystery of idiopathic male infertility: Is oxidative stress an actual risk? J. Urol. 2013, 190, 1842–1843. [Google Scholar] [CrossRef] [PubMed]
- Traini, G.; Tamburrino, L.; Vignozzi, L.; Baldi, E.; Marchiani, S. Is oxidative stress evaluated in viable human spermatozoa a marker of good semen quality? Front. Endocrinol. 2022, 13, 1012416. [Google Scholar] [CrossRef] [PubMed]
- Van de Hoek, M.; Rickard, J.P.; de Graaf, S.P. Motility assessment of ram spermatozoa. Biology 2022, 11, 1715. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Duan, X.; Xiong, B.; Cui, X.S.; Kim, N.H.; Rui, R.; Sun, S.C. ROCK inhibitor Y-27632 prevents porcine oocyte maturation. Theriogenology 2014, 82, 49–56. [Google Scholar] [CrossRef]
- Banihani, S.A.; Khasawneh, F.H. Effect of lansoprazole on human sperm motility, sperm viability, seminal nitric oxide production, and seminal calcium chelation. Res. Pharm. Sci. 2018, 13, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Mata-Martínez, E.; Sánchez-Tusie, A.A.; Darszon, A.; Mayorga, L.S.; Treviño, C.L.; De Blas, G.A. Epac activation induces an extracellular Ca(2+)-independent Ca(2+) wave that triggers acrosome reaction in human spermatozoa. Andrology 2021, 9, 1227–1241. [Google Scholar] [CrossRef] [PubMed]
- Stiavnicka, M.; Hosek, P.; Abril-Parreno, L.; Kenny, D.A.; Lonergan, P.; Fair, S. Membrane remodulation and hyperactivation are impaired in frozen- thawed sperm of low-fertility bulls. Theriogenology 2023, 195, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Green, C.; Watson, P. Comparison of the capacitation-like state of cooled boar spermatozoa with true capacitation. Reproduction 2001, 122, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Shen, J.; Wang, Y.; Pan, C.; Pang, W.; Diao, H.; Dong, W. Boar seminal plasma exosomes maintain sperm function by infiltrating into the sperm membrane. Oncotarget 2016, 7, 58832. [Google Scholar] [CrossRef]
- Stanishevskaya, O.I.; Silyukova, Y.; Fedorova, E.; Pleshanov, N.; Kurochkin, A.; Tereshina, V.M.; Ianutsevich, E. Effects of Trehalose Supplementation on Lipid Composition of Rooster Spermatozoa Membranes in a Freeze/Thaw Protocol. Animals 2023, 13, 1023. [Google Scholar] [CrossRef]
- White, I. Lipids and calcium uptake of sperm in relation to cold shock and preservation: A review. Reprod. Fertil. Dev. 1993, 5, 639–658. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.I.; Squires, E.L.; Graham, J.K. Adding cholesterol to the stallion sperm plasma membrane improves cryosurvival. Cryobiology 2005, 51, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Rajoriya, J.S.; Prasad, J.K.; Ramteke, S.S.; Perumal, P.; De, A.K.; Ghosh, S.K.; Bag, S.; Raje, A.; Singh, M.; Kumar, A.; et al. Exogenous cholesterol prevents cryocapacitation-like changes, membrane fluidity, and enhances in vitro fertility in bubaline spermatozoa. Reprod. Domest. Anim. 2020, 55, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Batissaco, L.; de Arruda, R.P.; Alves, M.B.; Torres, M.A.; Lemes, K.M.; Prado-Filho, R.R.; de Almeida, T.G.; de Andrade, A.F.; Celeghini, E.C. Cholesterol-loaded cyclodextrin is efficient in preserving sperm quality of cryopreserved ram semen with low freezability. Reprod. Biol. 2020, 20, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Goñi, F.M. Sphingomyelin: What is it good for? Biochem. Biophys. Res. Commun. 2022, 633, 23–25. [Google Scholar] [CrossRef] [PubMed]
- van der Veen, J.N.; Kennelly, J.P.; Wan, S.; Vance, J.E.; Vance, D.E.; Jacobs, R.L. The critical role of phosphatidylcholine and phosphatidylethanolamine metabolism in health and disease. Biochim. Et Biophys. Acta (BBA) Biomembr. 2017, 1859, 1558–1572. [Google Scholar] [CrossRef] [PubMed]
- Ahumada-Gutierrez, H.; Peñalva, D.; Enriz, R.; Antollini, S.; Cascales, J.L. Mechanical properties of bilayers containing sperm sphingomyelins and ceramides with very long-chain polyunsaturated fatty acids. Chem. Phys. Lipids 2019, 218, 178–186. [Google Scholar] [CrossRef]
- Lucio, C.; Brito, M.; Angrimani, D.; Belaz, K.; Morais, D.; Zampieri, D.; Losano, J.; Assumpção, M.; Nichi, M.; Eberlin, M.; et al. Lipid composition of the canine sperm plasma membrane as markers of sperm motility. Reprod. Domest. Anim. 2017, 52, 208–213. [Google Scholar] [CrossRef]
- Pfeiffer, R.L.; Marc, R.E.; Jones, B.W. Müller Cell Metabolic Signatures: Evolutionary Conservation and Disruption in Disease. Trends Endocrinol. Metab. 2020, 31, 320–329. [Google Scholar] [CrossRef]
- Yoo, H.C.; Yu, Y.C.; Sung, Y.; Han, J.M. Glutamine reliance in cell metabolism. Exp. Mol. Med. 2020, 52, 1496–1516. [Google Scholar] [CrossRef]
- Bansal, A.; Celeste Simon, M. Glutathione metabolism in cancer progression and treatment resistance. J. Cell Biol. 2018, 217, 2291–2298. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | VIP | p-Value | FC |
|---|---|---|---|
| Isocaproic acid | 2.10472657 | 0.001404849 | 1.640539 |
| 2’,4’-Dihydroxyacetophenone | 2.360230926 | 0.000002668 | 1.706914 |
| Decanoylcarnitine | 1.626249997 | 0.002329103 | 1.707014 |
| L-Octanoylcarnitine | 2.462864583 | 0.000000073 | 1.789571 |
| L-Hexanoylcarnitine | 2.458776918 | 0.000000011 | 1.876355 |
| (25S)-26-Hydroxy-24-methylenecycloartan-3-one | 2.285585719 | 0.000000041 | 2.173294 |
| 2-Acetylpyrazine | 2.44681402 | 0.000000012 | 2.336872 |
| 2-Methylbenzoic acid | 2.544358081 | 0.000000769 | 2.397188 |
| L-Nicotine | 1.950733563 | 0.002509318 | 3.111003 |
| 2,3,4,5-Tetrahydropiperidine-2-carboxylate | 2.464528754 | 0.000000057 | 3.380712 |
| 1-Isothiocyanato-7-(methylsulfinyl)heptane | 1.198204213 | 0.030862102 | 0.248183 |
| LysoPE(20:4(8Z, 11Z, 14Z, 17Z)/0:0) | 2.326823113 | 0.000003663 | 0.376789 |
| SM(d18:0/18:1(9Z)) | 1.79944416 | 0.036462866 | 0.462591 |
| LysoPE(0:0/22:5(4Z, 7Z, 10Z, 13Z, 16Z)) | 1.862054009 | 0.000668867 | 0.499072 |
| 9,10-Epoxyoctadecenoic acid | 2.165526982 | 0.000034265 | 0.508835 |
| Byssochlamic acid | 2.118297325 | 0.001568542 | 0.548432 |
| Palmitoleic acid | 2.132674871 | 0.000147640 | 0.548844 |
| 9,10-epoxyoctadecanoic acid | 2.159295557 | 0.000027289 | 0.597584 |
| 7,8-Dehydro-beta-micropteroxanthin | 1.642921181 | 0.010008768 | 0.614358 |
| 3-Methyl-2-oxopentanoate | 2.101091738 | 0.000441510 | 0.650236 |
| Name | p-Value | Number |
|---|---|---|
| Glycerophospholipid metabolism | 0.002131231 | 4 |
| Biosynthesis of amino acids | 0.001475389 | 6 |
| Taurine and hypotaurine metabolism | 0.001407924 | 3 |
| Arginine biosynthesis | 0.001407924 | 3 |
| Citrate cycle (TCA cycle) | 0.000925305 | 3 |
| Carbon metabolism | 0.000804546 | 6 |
| Propanoate metabolism | 0.000595049 | 4 |
| Metabolic pathways | 0.000571736 | 39 |
| Proximal tubule bicarbonate reclamation | 0.000563529 | 3 |
| D-Amino acid metabolism | 0.000466421 | 5 |
| Glyoxylate and dicarboxylate metabolism | 0.000324201 | 5 |
| Glucagon signaling pathway | 0.000107687 | 4 |
| ABC transporters | 0.000049142 | 8 |
| Retrograde endocannabinoid signaling | 0.000029368 | 4 |
| Central carbon metabolism in cancer | 0.000001238 | 6 |
| Pathway | Total | Hits | Raw p | Holm Adjust | Impact |
|---|---|---|---|---|---|
| D-Glutamine and D-glutamate metabolism | 5 | 3 | 0.000468 | 0.037905 | 1 |
| Nitrogen metabolism | 9 | 2 | 0.041713 | 1 | 0 |
| Alanine, aspartate, and glutamate metabolism | 23 | 3 | 0.05148 | 1 | 0.4 |
| beta-Alanine metabolism | 17 | 2 | 0.13046 | 1 | 0.44444 |
| Glycerolipid metabolism | 18 | 2 | 0.14339 | 1 | 0.28098 |
| Taurine and hypotaurine metabolism | 7 | 1 | 0.23455 | 1 | 0.75 |
| Semen Type | Fertilized Ewes (n) | Fertility (%) |
|---|---|---|
| Fresh semen | 50 | 43.2 ± 0.26 c |
| Semen with 20 μM Y-27632 | 50 | 42.5 ± 0.15 bc |
| Semen without 20 μM Y-27632 | 50 | 31.2 ± 0.23 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Sun, S.; Fu, Y.; Wang, L.; Ren, C.; Ling, Y.; Zhang, Z.; Cao, H. Positive In Vitro Effect of ROCK Pathway Inhibitor Y-27632 on Qualitative Characteristics of Goat Sperm Stored at Low Temperatures. Animals 2024, 14, 1441. https://doi.org/10.3390/ani14101441
Xu Y, Sun S, Fu Y, Wang L, Ren C, Ling Y, Zhang Z, Cao H. Positive In Vitro Effect of ROCK Pathway Inhibitor Y-27632 on Qualitative Characteristics of Goat Sperm Stored at Low Temperatures. Animals. 2024; 14(10):1441. https://doi.org/10.3390/ani14101441
Chicago/Turabian StyleXu, Yongjie, Shixin Sun, Yu Fu, Lei Wang, Chunhuan Ren, Yinghui Ling, Zijun Zhang, and Hongguo Cao. 2024. "Positive In Vitro Effect of ROCK Pathway Inhibitor Y-27632 on Qualitative Characteristics of Goat Sperm Stored at Low Temperatures" Animals 14, no. 10: 1441. https://doi.org/10.3390/ani14101441
APA StyleXu, Y., Sun, S., Fu, Y., Wang, L., Ren, C., Ling, Y., Zhang, Z., & Cao, H. (2024). Positive In Vitro Effect of ROCK Pathway Inhibitor Y-27632 on Qualitative Characteristics of Goat Sperm Stored at Low Temperatures. Animals, 14(10), 1441. https://doi.org/10.3390/ani14101441