
Skin Cortisol and Acoustic Activity: Potential Tools to Evaluate Stress and Welfare in Captive Cetaceans

 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
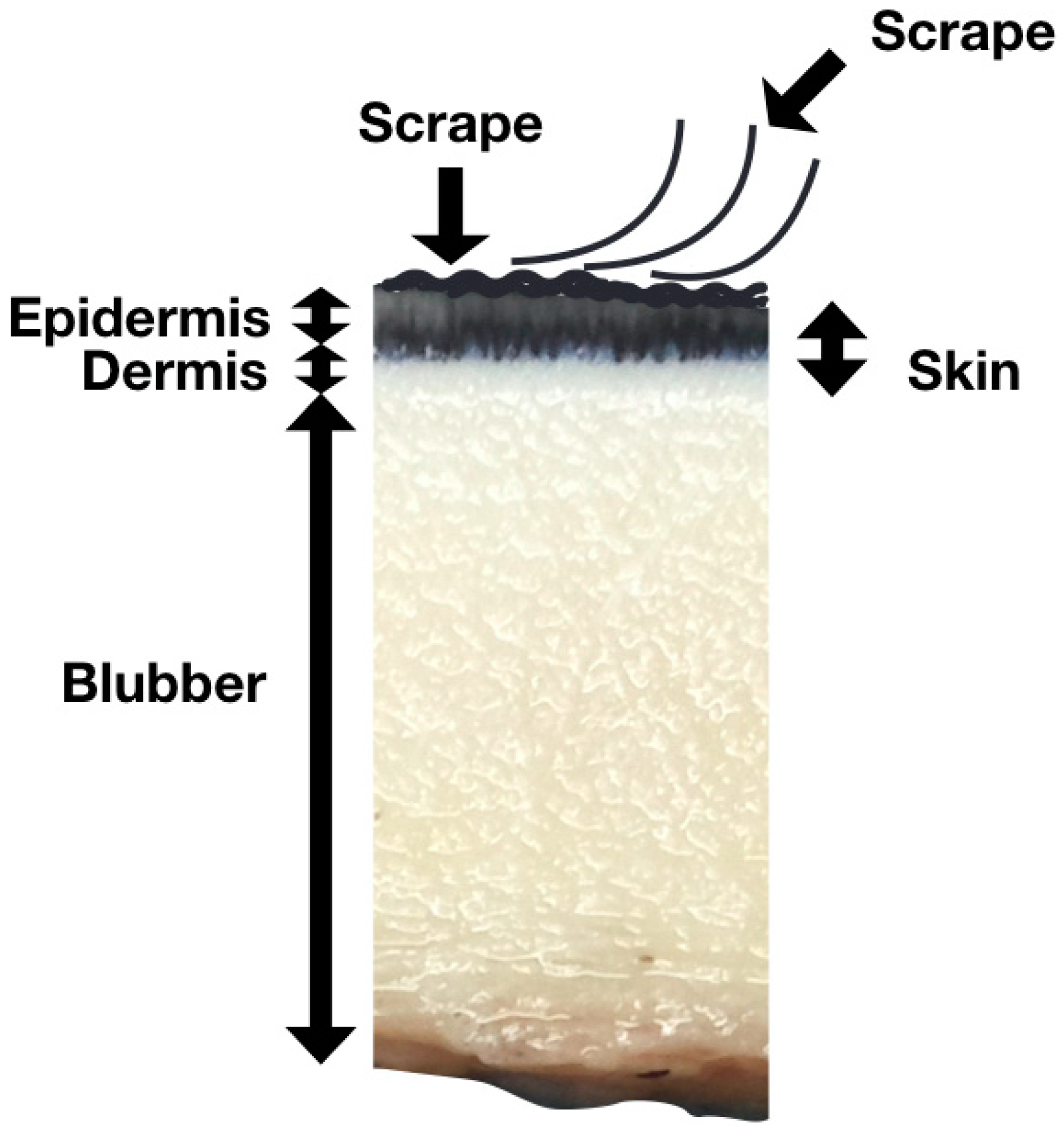
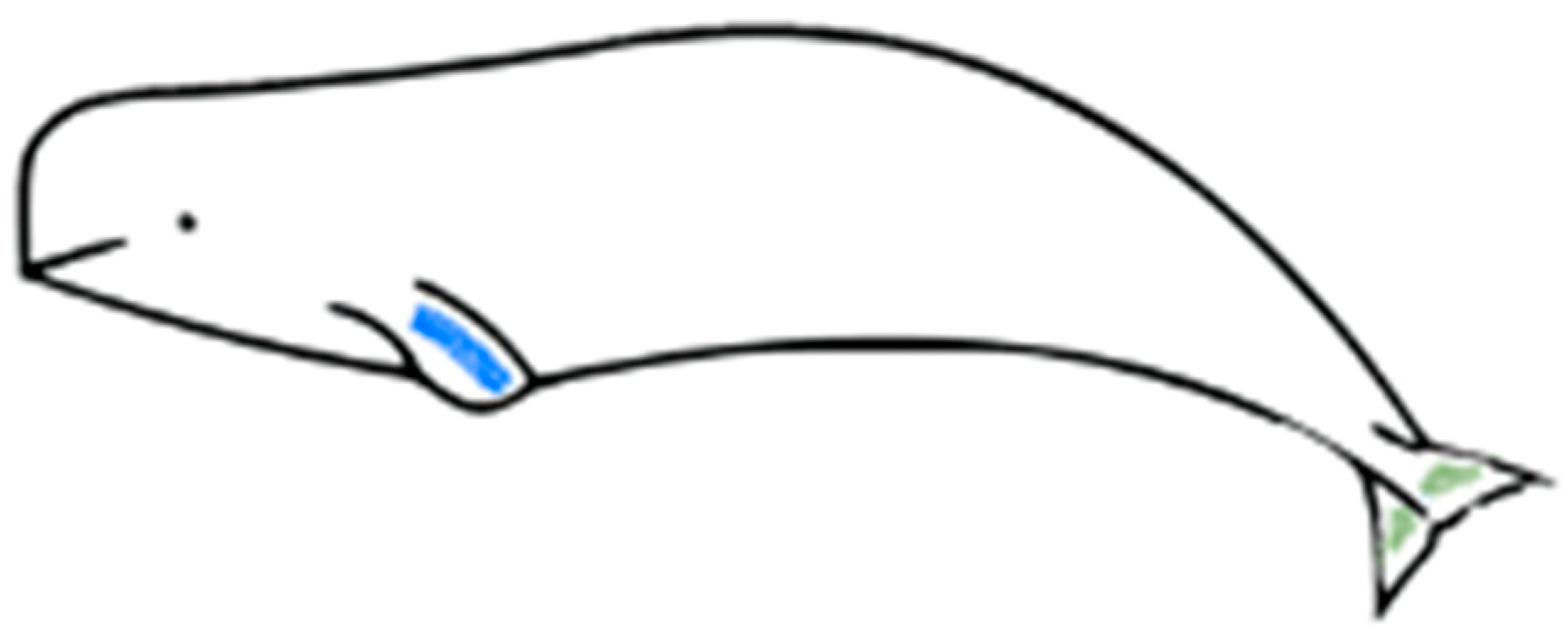
2.1. Scrape Samples Collection
2.2. Hormone Extraction
2.3. Cortisol Detection
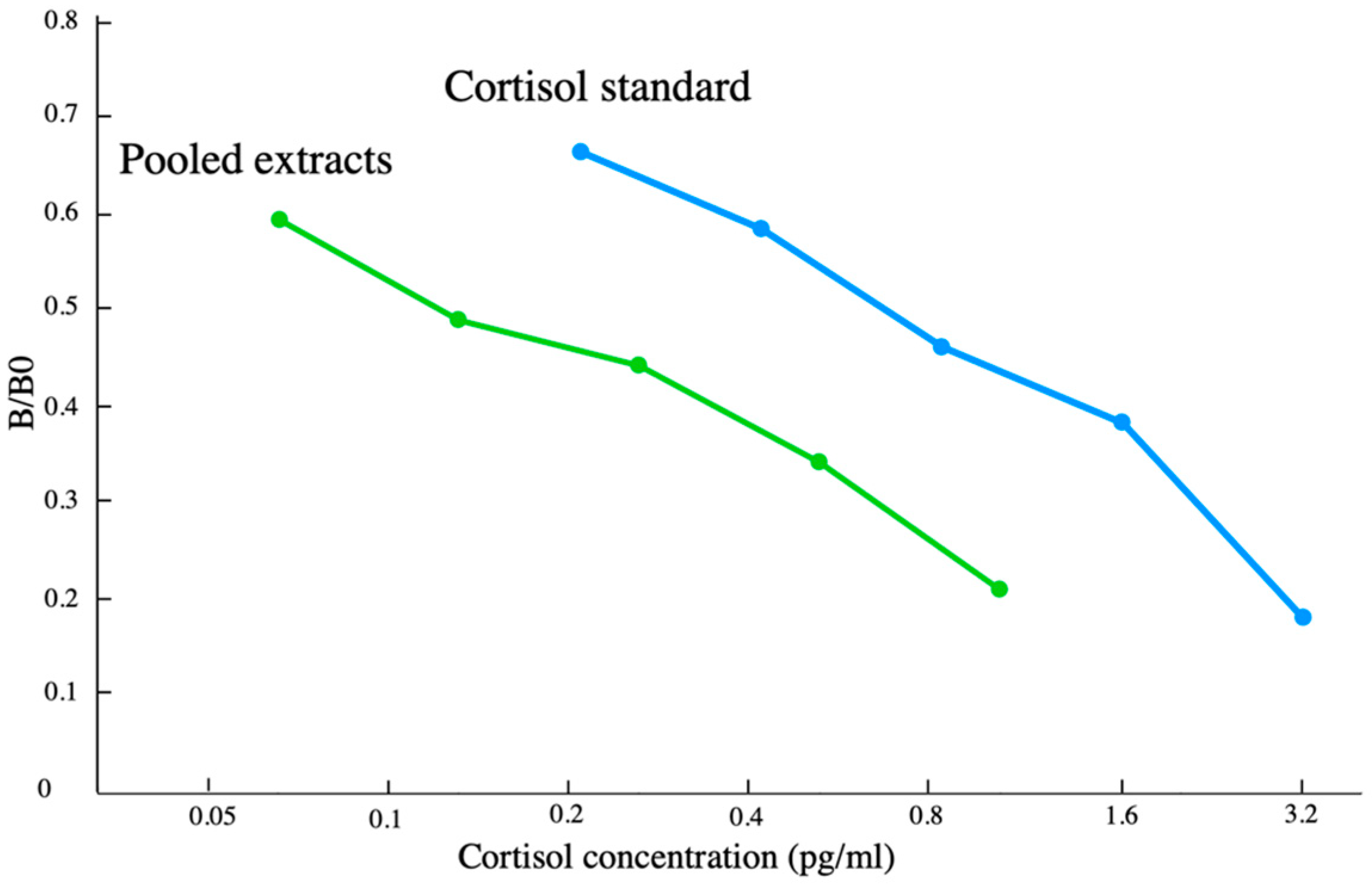
2.4. Parallelism and Matrix Effects Analyses
2.5. Time Lag Estimation
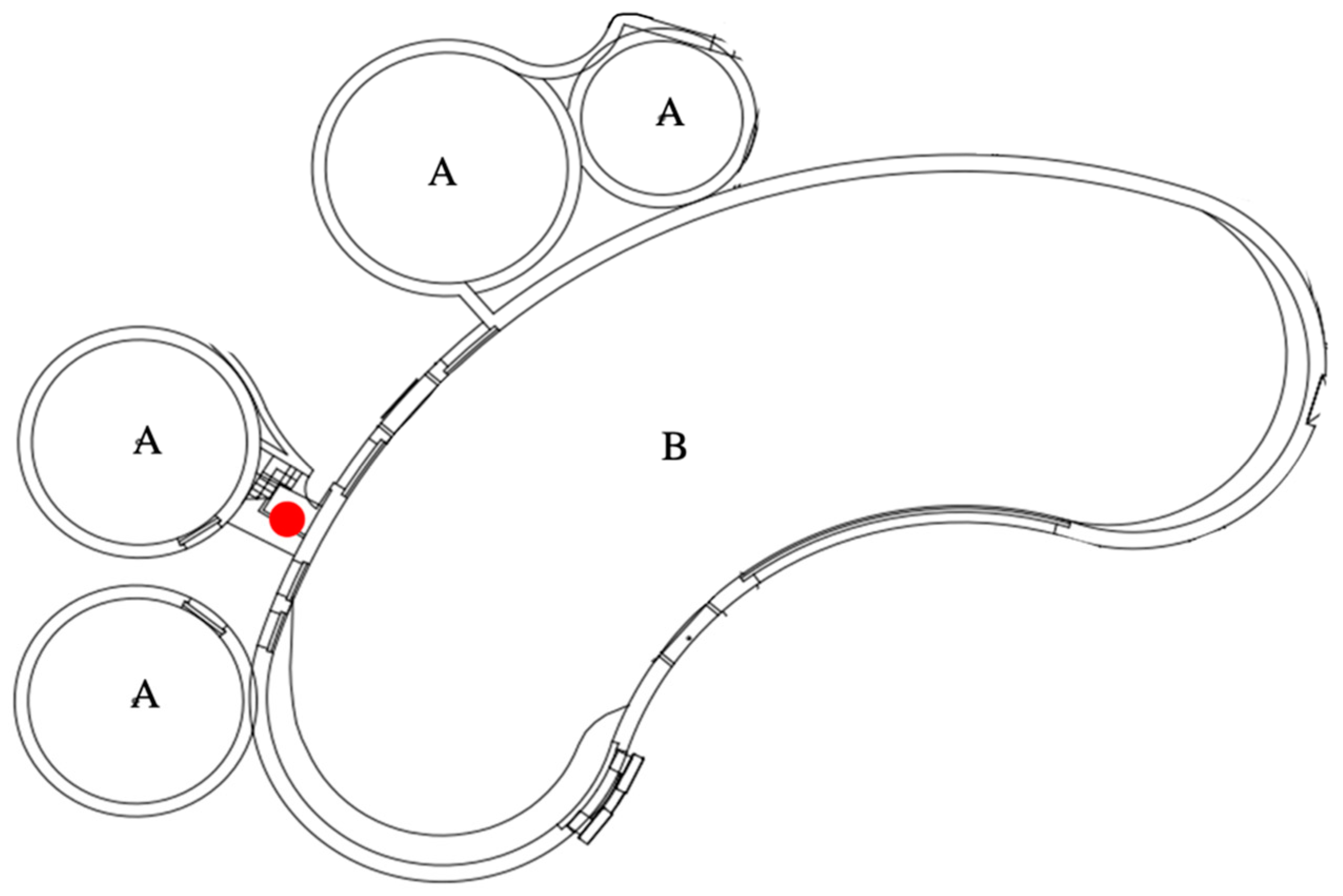
2.6. Acoustic Activities and Analyses
2.7. Statistical Analysis
3. Result
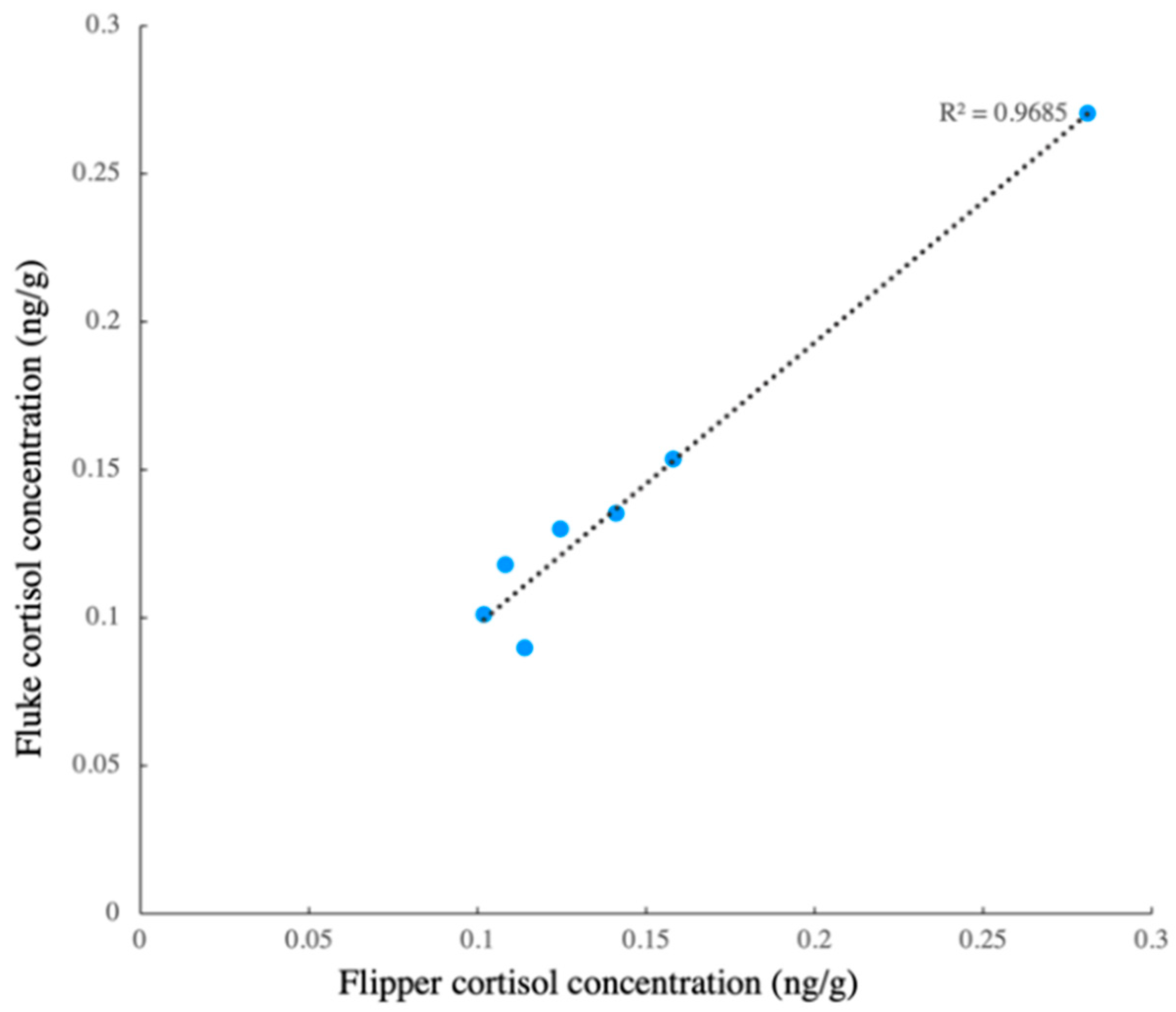
3.1. EIA Performance
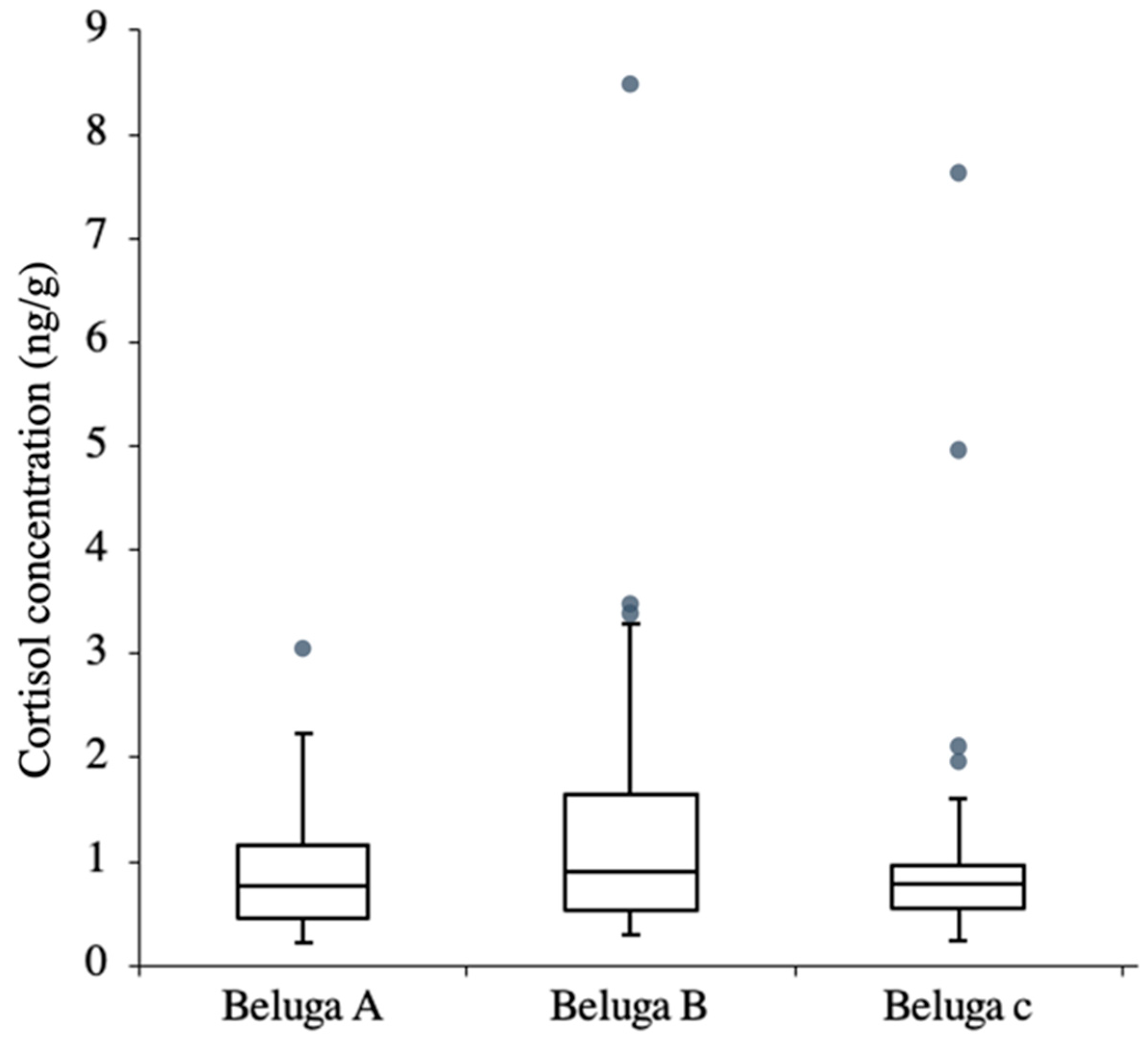
3.2. Scrape Cortisol
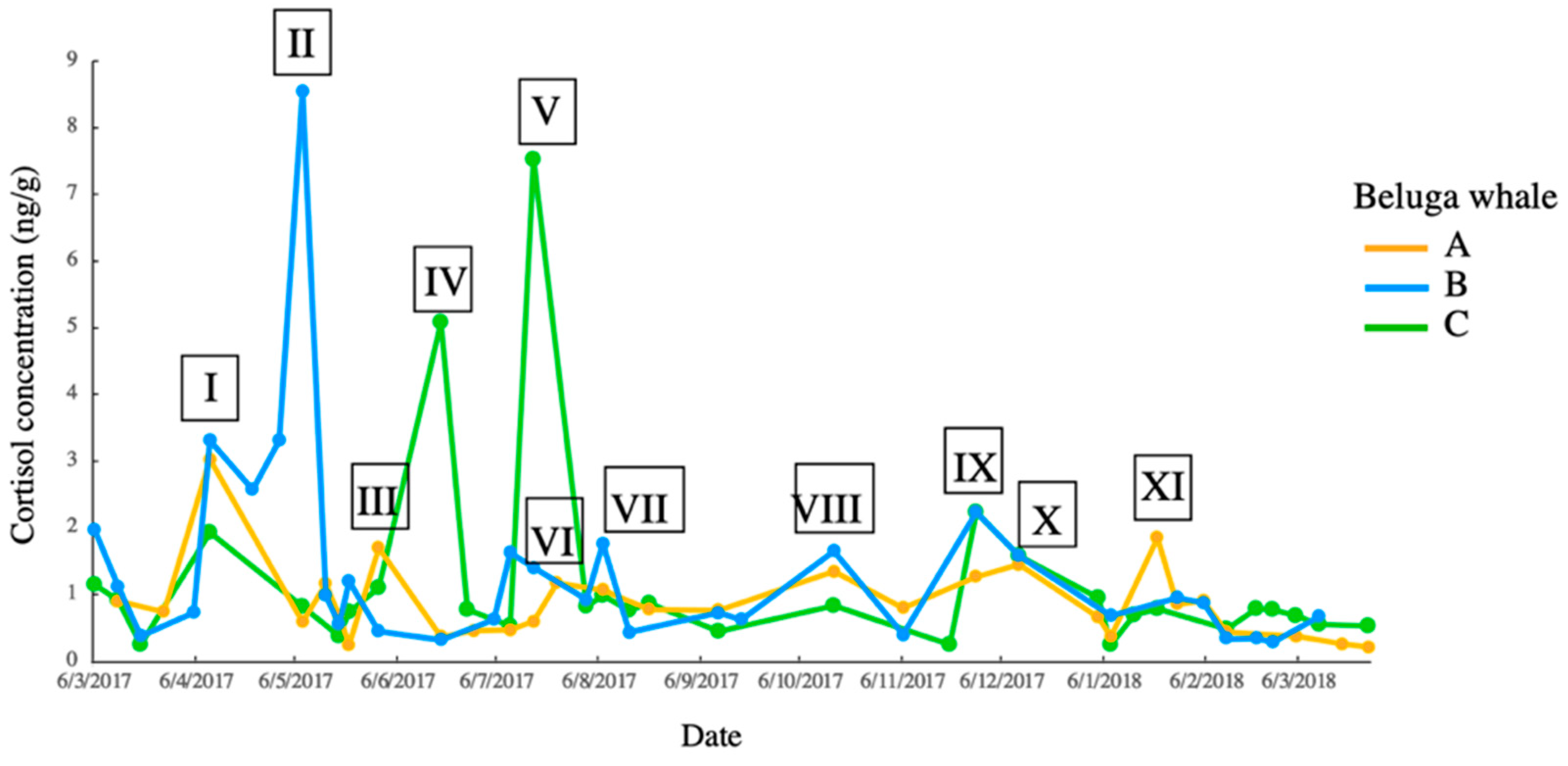
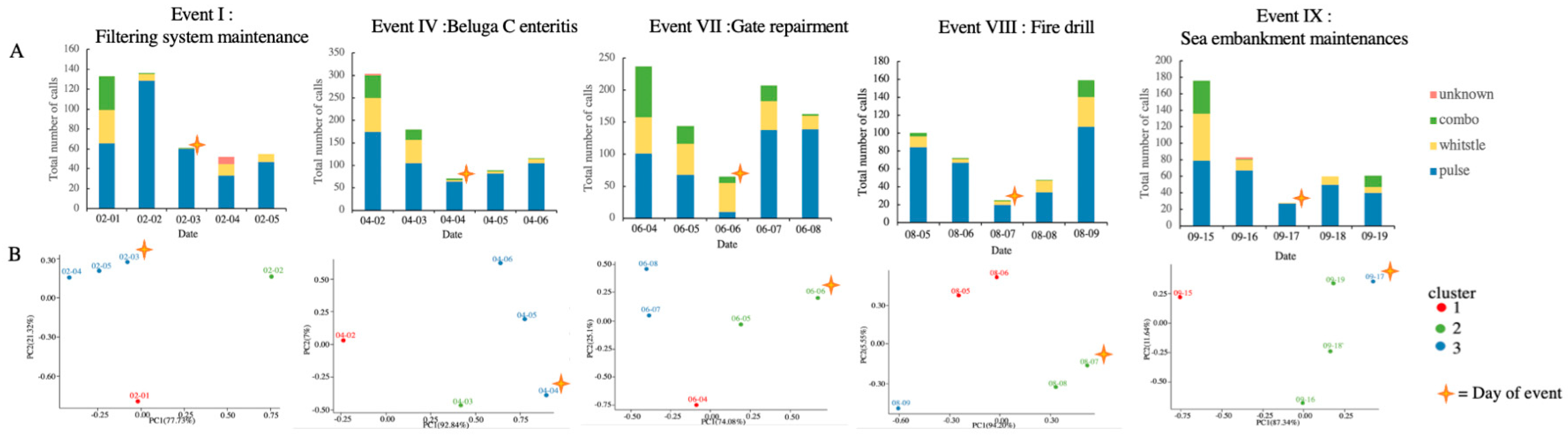
3.3. Acoustic Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mellor, D.; Reid, C. Concepts of animal well-being and predicting the impact of procedures on experimental animals. In Improving the Well-Being of Animals in the Research Environment; Baker, R.M., Jenkin, G., Mellor, D.J., Eds.; Australian and New Zealand Council for the Care of Animals in Research and Teaching: Glen Osmond, Australia, 1994; pp. 3–18. ISBN 0646181165. [Google Scholar]
- Mellor, D.J.; Beausoleil, N.J.; Littlewood, K.E.; McLean, A.N.; McGreevy, P.D.; Jones, B.; Wilkins, C. The 2020 five domains model: Including human–animal interactions in assessments of animal welfare. Animals 2020, 10, 1870. [Google Scholar] [CrossRef] [PubMed]
- Waples, K.A.; Gales, N.J. Evaluating and minimising social stress in the care of captive bottlenose dolphins (Tursiops aduncus). Zoo Biol. 2002, 21, 5–26. [Google Scholar] [CrossRef]
- Moberg, G.P. A model for assessing the impact of behavioral stress on domestic animals. J. Anim. Sci. 1987, 65, 1228–1235. [Google Scholar] [CrossRef] [PubMed]
- Wiepkema, P.; Koolhaas, J. Stress and Animal Welfare. Anim. Welf. 1993, 2, 195–218. [Google Scholar] [CrossRef]
- Broom, D.M.; Johnson, K.G. One Welfare, One Health, One Stress: Humans and Other Animals, Animal Welfare, 2nd ed.; Springer International Publishing: Cham, Switzerland, 2019; Volume 19, ISBN 978-3-030-32152-9. [Google Scholar]
- National Research Council. Stress and Distress: Definitions; National Academies Press: Washington, DC, USA, 2008; ISBN 978-0-309-10817-1. [Google Scholar]
- Golbidi, S.; Frisbee, J.C.; Laher, I. Chronic stress impacts the cardiovascular system: Animal models and clinical outcomes. Am. J. Physiol. Circ. Physiol. 2015, 308, H1476–H1498. [Google Scholar] [CrossRef]
- Veissier, I.; Boissy, A. Stress and welfare: Two complementary concepts that are intrinsically related to the animal’s point of view. Physiol. Behav. 2007, 92, 429–433. [Google Scholar] [CrossRef]
- Lee, C.; Colditz, I.G.; Campbell, D.L.M. A Framework to Assess the Impact of New Animal Management Technologies on Welfare: A Case Study of Virtual Fencing. Front. Vet. Sci. 2018, 5, 187. [Google Scholar] [CrossRef] [PubMed]
- Bayazit, V. Evaluation of Cortisol and Stress in Captive Animals. Aust. J. Basic Appl. Sci. 2009, 3, 1022–1031. [Google Scholar]
- Trumble, S.J.; Norman, S.A.; Crain, D.D.; Mansouri, F.; Winfield, Z.C.; Sabin, R.; Potter, C.W.; Gabriele, C.M.; Usenko, S. Baleen whale cortisol levels reveal a physiological response to 20th century whaling. Nat. Commun. 2018, 9, 4587. [Google Scholar] [CrossRef]
- Champagne, C.D.; Kellar, N.M.; Trego, M.L.; Delehanty, B.; Boonstra, R.; Wasser, S.K.; Booth, R.K.; Crocker, D.E.; Houser, D.S. Comprehensive endocrine response to acute stress in the bottlenose dolphin from serum, blubber, and feces. Gen. Comp. Endocrinol. 2018, 266, 178–193. [Google Scholar] [CrossRef]
- Heimbürge, S.; Kanitz, E.; Otten, W. The use of hair cortisol for the assessment of stress in animals. Gen. Comp. Endocrinol. 2019, 270, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Möstl, E.; Palme, R. Hormones as indicators of stress. Domest. Anim. Endocrinol. 2002, 23, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Kershaw, J.L.; Hall, A.J. Seasonal variation in harbour seal (Phoca vitulina) blubber cortisol—A novel indicator of physiological state? Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sheriff, M.J.; Dantzer, B.; Delehanty, B.; Palme, R.; Boonstra, R. Measuring stress in wildlife: Techniques for quantifying glucocorticoids. Oecologia 2011, 166, 869–887. [Google Scholar] [CrossRef] [PubMed]
- Monreal-Pawlowsky, T.; Carbajal, A.; Tallo-Parra, O.; Sabés-Alsina, M.; Monclús, L.; Almunia, J.; Fernández-Bellon, H.; Lopez-Bejar, M. Daily salivary cortisol levels in response to stress factors in captive common bottlenose dolphins (Tursiops truncatus): A potential welfare indicator. Vet. Rec. 2017, 180, 593. [Google Scholar] [CrossRef]
- Nemeth, M.; Pschernig, E.; Wallner, B.; Millesi, E. Non-invasive cortisol measurements as indicators of physiological stress responses in guinea pigs. PeerJ 2016, 4, e1590. [Google Scholar] [CrossRef] [PubMed]
- Ovejero, R.; Novillo, A.; Soto-Gamboa, M.; Mosca-Torres, M.E.; Cuello, P.; Gregório, P.; Jahn, G.; Carmanchahi, P. Do cortisol and corticosterone play the same role in coping with stressors? Measuring glucocorticoid serum in free-ranging guanacos (Lama guanicoe). J. Exp. Zool. Part A Ecol. Genet. Physiol. 2013, 319, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Hadinger, U.; Haymerle, A.; Knauer, F.; Schwarzenberger, F.; Walzer, C. Faecal cortisol metabolites to assess stress in wildlife: Evaluation of a field method in free-ranging chamois. Methods Ecol. Evol. 2015, 6, 1349–1357. [Google Scholar] [CrossRef]
- Villalba, J.J.; Manteca, X. A Case for Eustress in Grazing Animals. Front. Vet. Sci. 2019, 6, 303. [Google Scholar] [CrossRef] [PubMed]
- Koolhaas, J.M.; Bartolomucci, A.; Buwalda, B.; de Boer, S.F.; Flügge, G.; Korte, S.M.; Meerlo, P.; Murison, R.; Olivier, B.; Palanza, P.; et al. Stress revisited: A critical evaluation of the stress concept. Neurosci. Biobehav. Rev. 2011, 35, 1291–1301. [Google Scholar] [CrossRef]
- Clegg, I.L.K.; Boys, R.M.; Stockin, K.A. Increasing the Awareness of Animal Welfare Science in Marine Mammal Conservation: Addressing Language, Translation and Reception Issues. Animals 2021, 11, 1596. [Google Scholar] [CrossRef] [PubMed]
- Clegg, I.L.K.; Borger-Turner, J.L.; Eskelinen, H.C. C-Well: The development of a welfare assessment index for captive bottlenose dolphins (Tursiops truncatus). Anim. Welf. 2015, 24, 267–282. [Google Scholar] [CrossRef]
- Clegg, I.L.K.; Rödel, H.G.; Mercera, B.; van der Heul, S.; Schrijvers, T.; de Laender, P.; Gojceta, R.; Zimmitti, M.; Verhoeven, E.; Burger, E.; et al. Dolphins’ Willingness to Participate (WtP) in positive reinforcement training as a potential welfare indicator, where WtP predicts early changes in health status. Front. Psychol. 2019, 10, 2112. [Google Scholar] [CrossRef]
- Ugaz, C.; Valdez, R.A.; Romano, M.C.; Galindo, F. Behavior and salivary cortisol of captive dolphins (Tursiops truncatus) kept in open and closed facilities. J. Vet. Behav. Clin. Appl. Res. 2013, 8, 285–290. [Google Scholar] [CrossRef]
- Castellote, M.; Fossa, F. Measuring acoustic activity as a method to evaluate welfare in captive beluga whales (Delphinapterus leucas). Aquat. Mamm. 2006, 32, 325–333. [Google Scholar] [CrossRef]
- Norcross, J.L.; Newman, J.D. Effects of separation and novelty on distress vocalizations and cortisol in the common marmoset (Callithrix jacchus). Am. J. Primatol. 1999, 47, 209–222. [Google Scholar] [CrossRef]
- Jefferson, T.A.; Webber, M.A.; Pitman, R. Marine Mammals of the World; Elsevier: Amsterdam, The Netherlands, 2015; ISBN 9780123838537. [Google Scholar]
- Belikov, R.A.; Bel’Kovich, V.M. Whistles of beluga whales in the reproductive gathering off Solovetskii Island in the White Sea. Acoust. Phys. 2007, 53, 528–534. [Google Scholar] [CrossRef]
- Vergara, V.; Barrett-Lennard, L.G. Vocal development in a beluga calf (Delphinapterus leucas). Aquat. Mamm. 2008, 34, 123–143. [Google Scholar] [CrossRef]
- Panova, E.M.; Belikov, R.A.; Agafonov, A.V.; Bel’kovich, V.M. The relationship between the behavioral activity and the underwater vocalization of the beluga whale (Delphinapterus leucas). Oceanology 2012, 52, 79–87. [Google Scholar] [CrossRef]
- Garland, E.C.; Castellote, M.; Berchok, C.L. Beluga whale (Delphinapterus leucas) vocalizations and call classification from the eastern Beaufort Sea population. J. Acoust. Soc. Am. 2015, 137, 3054–3067. [Google Scholar] [CrossRef]
- Chmelnitsky, E.G.; Ferguson, S.H. Beluga whale, Delphinapterus leucas, vocalizations from the Churchill River, Manitoba, Canada. J. Acoust. Soc. Am. 2012, 131, 4821–4835. [Google Scholar] [CrossRef] [PubMed]
- Karlsen, J.D.; Bisther, A.; Lydersen, C.; Haug, T.; Kovacs, K.M. Summer vocalisations of adult male white whales (Delphinapterus leucas) in Svalbard, Norway. Polar Biol. 2002, 25, 808–817. [Google Scholar] [CrossRef]
- Yang, W.C.; Chen, C.F.; Chuah, Y.C.; Zhuang, C.R.; Chen, I.H.; Mooney, T.A.; Stott, J.; Blanchard, M.; Jen, I.F.; Chou, L.S. Anthropogenic Sound Exposure-Induced Stress in Captive Dolphins and Implications for Cetacean Health. Front. Mar. Sci. 2021, 8, 1–8. [Google Scholar] [CrossRef]
- Tsai, M.-A.; Chen, I.-H.; Wang, J.-H.; Chou, S.-J.; Li, T.-H.; Leu, M.-Y.; Ho, H.-K.; Yang, W.C. A probe-based qRT-PCR method to profile immunological gene expression in blood of captive beluga whales (Delphinapterus leucas). PeerJ 2017, 5, e3840. [Google Scholar] [CrossRef] [PubMed]
- Kellar, N.M.; Catelani, K.N.; Robbins, M.N.; Trego, M.L.; Allen, C.D.; Danil, K.; Chivers, S.J. Blubber cortisol: A potential tool for assessing stress response in free-ranging dolphins without effects due to sampling. PLoS ONE 2015, 10, e0115257. [Google Scholar] [CrossRef] [PubMed]
- Hunt, K.E.; Lysiak, N.S.J.; Matthews, C.J.D.; Lowe, C.; Ajó, A.F.; Dillon, D.; Willing, C.; Heide-Jørgensen, M.P.; Ferguson, S.H.; Moore, M.J.; et al. Multi-year patterns in testosterone, cortisol and corticosterone in baleen from adult males of three whale species. Conserv. Physiol. 2018, 6, coy049. [Google Scholar] [CrossRef]
- Bechshoft, T.; Wright, A.J.; Styrishave, B.; Houser, D. Measuring and validating concentrations of steroid hormones in the skin of bottlenose dolphins (Tursiops truncatus). Conserv. Physiol. 2020, 8, coaa032. [Google Scholar] [CrossRef]
- Champagne, C.D.; Kellar, N.M.; Crocker, D.E.; Wasser, S.K.; Booth, R.K.; Trego, M.L.; Houser, D.S. Blubber cortisol qualitatively reflects circulating cortisol concentrations in bottlenose dolphins. Mar. Mammal Sci. 2017, 33, 134–153. [Google Scholar] [CrossRef]
- Loseto, L.L.; Pleskach, K.; Hoover, C.; Tomy, G.T.; Desforges, J.-P.; Halldorson, T.; Ross, P.S. Cortisol levels in beluga whales (Delphinapterus leucas): Setting a benchmark for Marine Protected Area monitoring. Arct. Sci. 2017, 372, 358–372. [Google Scholar] [CrossRef]
- Rolland, R.M.; McLellan, W.A.; Moore, M.J.; Harms, C.A.; Burgess, E.A.; Hunt, K.E. Fecal glucocorticoids and anthropogenic injury and mortality in North Atlantic right whales Eubalaena glacialis. Endanger. Species Res. 2017, 34, 417–429. [Google Scholar] [CrossRef]
- Thompson, L.A.; Spoon, T.R.; Goertz, C.E.C.; Hobbs, R.C.; Romano, T.A. Blow collection as a non-invasive method for measuring cortisol in the beluga (Delphinapterus leucas). PLoS ONE 2014, 9, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Bechshoft, T.; Wright, A.J.; Weisser, J.J.; Teilmann, J.; Dietz, R.; Hansen, M.; Björklund, E.; Styrishave, B. Developing a new research tool for use in free-ranging cetaceans: Recovering cortisol from harbour porpoise skin. Conserv. Physiol. 2015, 3, cov016. [Google Scholar] [CrossRef] [PubMed]
- Watt, C.A.; Simonee, J.; Herault, V.L.; Zhou, R.; Ferguson, S.H.; Marcoux, M.; Black, S. Cortisol levels in narwhal (Monodon monoceros) blubber from 2000 to 2019. Arct. Sci. 2021, 7, 690–698. [Google Scholar] [CrossRef]
- Trana, M.R.; Roth, J.D.; Tomy, G.T.; Anderson, W.G.; Ferguson, S.H. Increased blubber cortisol in ice-entrapped beluga whales (Delphinapterus leucas). Polar Biol. 2016, 39, 1563–1569. [Google Scholar] [CrossRef]
- Hunt, K.E.; Lysiak, N.S.; Moore, M.J.; Rolland, R.M. Longitudinal progesterone profiles in baleen from female North Atlantic right whales (Eubalaena glacialis) match known calving history. Conserv. Physiol. 2016, 4, cow014. [Google Scholar] [CrossRef]
- Aubin, D.J.S.; Smith, T.G.; Geraci, J.R. Seasonal epidermal molt in beluga whales, Delphinapterus leucas. Can. J. Zool. 1990, 68, 359–367. [Google Scholar] [CrossRef]
- Geraci, J.R.; Bruce-Allen, L.J. Slow process of wound repair in beluga whales, Delphinapterus leucas. Can. J. Fish. Aquat. Sci. 1987, 44, 1661–1665. [Google Scholar] [CrossRef]
- Ryan, C.; Mchugh, B.; O’Connor, I.; Berrow, S. Lipid content of blubber biopsies is not representative of blubber in situ for fin whales (Balaenoptera physalus). Mar. Mammal Sci. 2013, 29, 542–547. [Google Scholar] [CrossRef]
- Kershaw, J.L.; Sherrill, M.; Davison, N.J.; Brownlow, A.; Hall, A.J. Evaluating morphometric and metabolic markers of body condition in a small cetacean, the harbor porpoise (Phocoena phocoena). Ecol. Evol. 2017, 7, 3494–3506. [Google Scholar] [CrossRef]
- Candiani, D.; Salamano, G.; Mellia, E.; Doglione, L.; Bruno, R.; Toussaint, M.; Gruys, E. A combination of behavioral and physiological indicators for assessing pig welfare on the farm. J. Appl. Anim. Welf. Sci. 2008, 11, 1–13. [Google Scholar] [CrossRef]
- Rajagopal, T.; Archunan, G.; Sekar, M. Impact of zoo visitors on the fecal cortisol levels and behavior of an endangered species: Indian blackbuck (Antelope cervicapra L.). J. Appl. Anim. Welf. Sci. 2011, 14, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Haverbeke, A.; Diederich, C.; Depiereux, E.; Giffroy, J.M. Cortisol and behavioral responses of working dogs to environmental challenges. Physiol. Behav. 2008, 93, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Laws, N.; Ganswindt, A.; Heistermann, M.; Harris, M.; Harris, S.; Sherwin, C. A Case Study: Fecal Corticosteroid and Behavior as Indicators of Welfare During Relocation of an Asian Elephant. J. Appl. Anim. Welf. Sci. 2007, 10, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Salas, M.; Temple, D.; Abáigar, T.; Cuadrado, M.; Delclaux, M.; Enseñat, C.; Almagro, V.; Martínez-Nevado, E.; Quevedo, M.Á.; Carbajal, A.; et al. Aggressive behavior and hair cortisol levels in captive Dorcas gazelles (Gazella dorcas) as animal-based welfare indicators. Zoo Biol. 2016, 35, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Wolfensohn, S.; Shotton, J.; Bowley, H.; Davies, S.; Thompson, S.; Justice, W.S.M. Assessment of welfare in zoo animals: Towards optimum quality of life. Animals 2018, 8, 110. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day | Beluga Whale | Points | ||
|---|---|---|---|---|
| A | B | C | ||
| 28–32 | 3 | 2 | 2 | 7 |
| 38–42 | 4 | 4 | 2 | 10 |
| 48–52 | 5 | 2 | 3 | 10 |
| 58–62 | 5 | 3 | 2 | 10 |
| 68–72 | 5 | 5 | 4 | 14 |
| 78–82 | 4 | 3 | 2 | 9 |
| Environmental Changes | Physiological Changes | ||
|---|---|---|---|
| I | Filtering system maintenance | / | |
| II | Filtering system maintenance | / | |
| III | Minor maintenance | / | |
| IV | / | Beluga C enteritis | |
| V | Construction of false bottom in medical pool | / | |
| VI | Gate repairment | / | |
| VII | Gate repairment | / | |
| VIII | Fire drill | / | |
| IX | Impulsive sound of unknown-source from the sea embankment | / | |
| X | Sound exposure experiment on beluga A | / | |
| XI | Maintenance of false bottom | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wong, C.-H.; Tsai, M.-A.; Ko, F.-C.; Wang, J.-H.; Xue, Y.-J.; Yang, W.-C. Skin Cortisol and Acoustic Activity: Potential Tools to Evaluate Stress and Welfare in Captive Cetaceans. Animals 2023, 13, 1521. https://doi.org/10.3390/ani13091521
Wong C-H, Tsai M-A, Ko F-C, Wang J-H, Xue Y-J, Yang W-C. Skin Cortisol and Acoustic Activity: Potential Tools to Evaluate Stress and Welfare in Captive Cetaceans. Animals. 2023; 13(9):1521. https://doi.org/10.3390/ani13091521
Chicago/Turabian StyleWong, Chung-Hei, Ming-An Tsai, Fung-Chi Ko, Jiann-Hsiung Wang, Yi-Jing Xue, and Wei-Cheng Yang. 2023. "Skin Cortisol and Acoustic Activity: Potential Tools to Evaluate Stress and Welfare in Captive Cetaceans" Animals 13, no. 9: 1521. https://doi.org/10.3390/ani13091521
APA StyleWong, C.-H., Tsai, M.-A., Ko, F.-C., Wang, J.-H., Xue, Y.-J., & Yang, W.-C. (2023). Skin Cortisol and Acoustic Activity: Potential Tools to Evaluate Stress and Welfare in Captive Cetaceans. Animals, 13(9), 1521. https://doi.org/10.3390/ani13091521