
Integrating Measures of Fecal Glucocorticoid Metabolites and Giving-Up Densities to Assess Adrenocortical Activity and Well-Being in Zoo-Housed Three-Banded Armadillos (Tolypeutes matacus)
 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Fecal Sample Collection and Processing
2.3. Hormone Analysis
2.4. Foraging Patch Procedure and Experiments
2.4.1. Substrate Quantity
2.4.2. Patch Quality
2.4.3. Cover Quantity
2.5. Integrating FGMs and GUDs into States of Stress and Well-Being
2.6. Data Analysis
2.6.1. Adrenocortical Activity
2.6.2. Foraging Behavior
2.6.3. Relationship between FGMs and GUDs
3. Results
3.1. Adrenocortical Activity
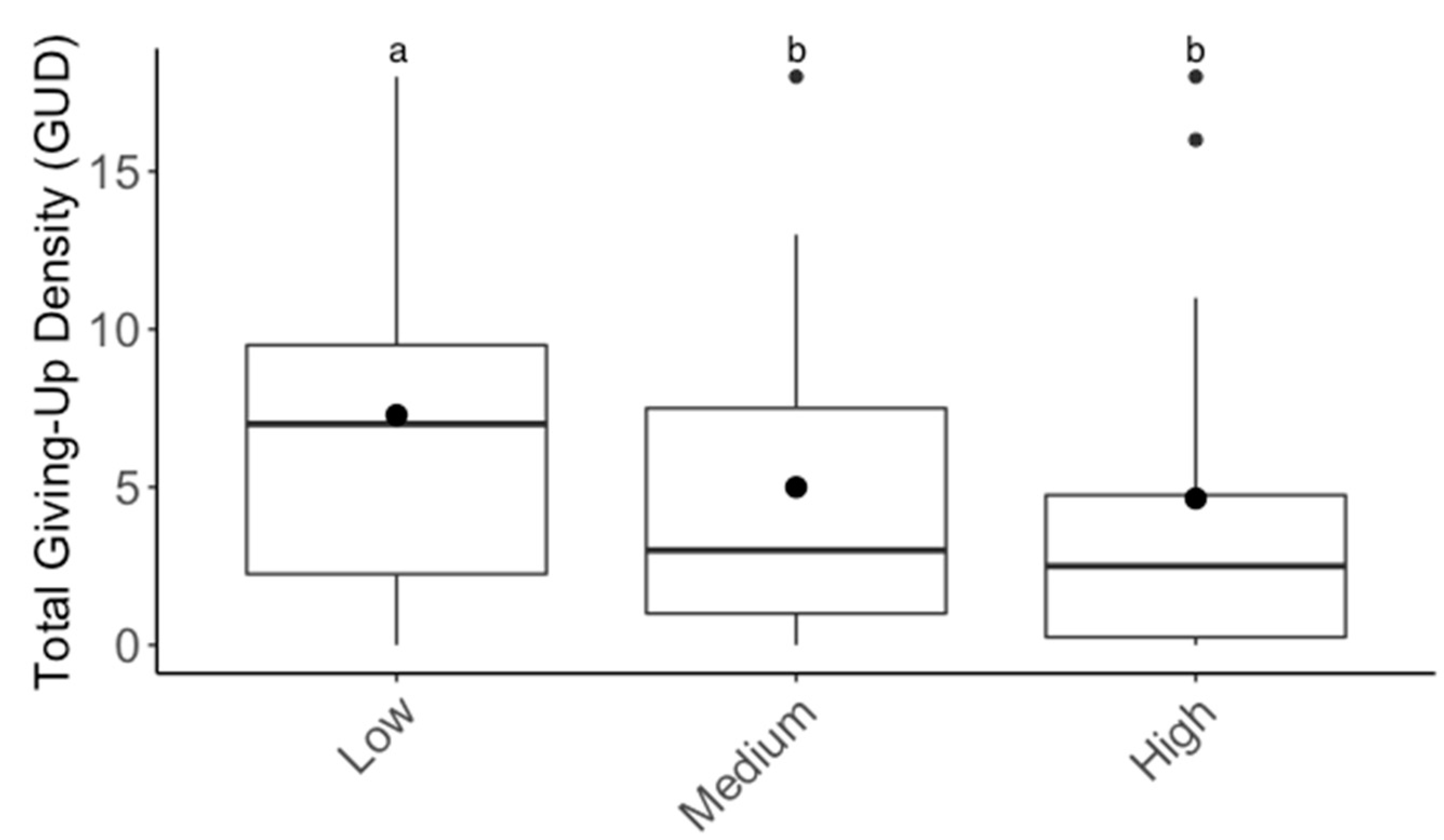
3.2. Foraging Behavior
3.3. Relationship between FGMs and GUDs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Edwards, K.L.; Edes, A.N.; Brown, J.L. Stress, Well-Being and Reproductive Success. In Reproductive Sciences in Animal Conservation. Advances in Experimental Medicine and Biology; Comizzoli, P., Brown, J., Holt, W., Eds.; Springer: Cham, Switzerland; New York, NY, USA, 2019; Volume 1200, pp. 91–162. [Google Scholar] [CrossRef]
- MacDougall-Shackleton, S.A.; Bonier, F.; Romero, L.M.; Moore, I.T. Glucocorticoids and “stress” are not synonymous. Integr. Org. Biol. 2019, 1, obz017. [Google Scholar] [CrossRef] [PubMed]
- Adkins-Regan, E. Hormones and Animal Social Behavior; Princeton University Press: Princeton, NJ, USA; Oxford, UK, 2005. [Google Scholar] [CrossRef]
- Kudielka, B.M.; Kirschbaum, C. Sex differences in HPA axis responses to stress: A review. Biol. Psychol. 2005, 69, 113–132. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Vale, W.W. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. Dialogues Clin. Neurosci. 2006, 8, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Moberg, G.P. Biological Response to Stress: Implications for Animal Welfare. In The Biology of Animal Stress: Basic Principles and Implications for Animal Welfare; CABI Pub: New York, NY, USA, 2000; pp. 1–21. [Google Scholar]
- McEwen, B.S.; Stellar, E. Stress and the individual: Mechanisms leading to disease. Arch. Intern. Med. 1993, 153, 2093–2101. [Google Scholar] [CrossRef]
- Nussey, S.S.; Whitehead, S.A. Endocrinology: An Integrated Approach; BIOS Scientific: Oxford, UK, 2001; ISBN 0-203-45043-4. [Google Scholar]
- Wingfield, J.C.; Hunt, K.; Breuner, C.; Dunlap, K.; Fowler, G.S.; Freed, L.; Lepson, J. Environmental Stress, Field Endocrinology, and Conservation Biology. In Behavioral Approaches to Conservation in the Wild; Cambridge University Press: Cambridge, UK, 1997; pp. 95–131. [Google Scholar]
- Wingfield, J.C.; Sapolsky, R.M. Reproduction and resistance to stress: When and how. J. Neuroendocrinol. 2003, 15, 711–724. [Google Scholar] [CrossRef]
- Brown, J.L.; Wildt, D.E.; Graham, L.H.; Byers, A.P.; Collins, L.; Barrett, S.; Howard, J. Natural versus chorionic gonadotropin-induced ovarian responses in the clouded leopard (Neofelis nebulosa) assessed by fecal steroid analysis. Biol. Reprod. 1995, 53, 93–102. [Google Scholar] [CrossRef]
- Touma, C.; Palme, R. Measuring fecal glucocorticoid metabolites in mammals and birds: The importance of validation. Ann. N. Y. Acad. Sci. 2005, 1046, 54–74. [Google Scholar] [CrossRef]
- Young, K.M.; Walker, S.L.; Lanthier, C.; Waddell, W.T.; Monfort, S.L.; Brown, J.L. Noninvasive monitoring of adrenocortical activity in carnivores by fecal glucocorticoid analyses. Gen. Comp. Endocrinol. 2004, 137, 148–165. [Google Scholar] [CrossRef]
- Palme, R. Measuring fecal steroids: Guidelines for practical application. Ann. N. Y. Acad. Sci. 2005, 1046, 75–80. [Google Scholar] [CrossRef]
- Monfort, S.L. Non-Invasive Endocrine Measures of Reproduction and Stress in Wild Populations. In Reproductive Science and Integrated Conservation; Holt, W.V., Ed.; Cambridge University Press: Cambridge, UK, 2003; pp. 147–165. [Google Scholar]
- Brown, J.L.; Somerville, M.; Riddle, H.S.; Keele, M.; Duer, C.K.; Freeman, E.W. Comparative endocrinology of testicular, adrenal and thyroid function in captive Asian and African elephant bulls. Gen. Comp. Endocrinol. 2007, 151, 153–162. [Google Scholar] [CrossRef]
- Swaisgood, R.R. Current status and future directions of applied behavioral research for animal welfare and conservation. Appl. Anim. Behav. Sci. 2007, 102, 139–162. [Google Scholar] [CrossRef]
- Kotler, B.P.; Morris, D.W.; Brown, J.S. Direct Behavioral Indicators as a Conservation and Management Tool. In Conservation Behavior; Berger-Tal, O., Saltz, D., Eds.; Cambridge University Press: Cambridge, UK, 2016; pp. 307–351. ISBN 978-1-107-69041-7. [Google Scholar]
- Hogan, L.A.; Johnston, S.D.; Lisle, A.; Horsup, A.B.; Janssen, T.; Phillips, C.J.C. Stereotypies and environmental enrichment in captive southern hairy-nosed wombats, Lasiorhinus latifrons. Appl. Anim. Behav. Sci. 2010, 126, 85–95. [Google Scholar] [CrossRef]
- Shivik, J.A.; Palmer, G.L.; Gese, E.M.; Osthaus, B. Captive coyotes compared to their counterparts in the wild: Does environmental enrichment help? J. Appl. Anim. Welf. Sci. 2009, 12, 223–235. [Google Scholar] [CrossRef]
- Cummings, D.; Brown, J.L.; Rodden, M.D.; Songsasen, N. Behavioral and physiologic responses to environmental enrichment in the maned wolf (Chrysocyon brachyurus). Zoo Biol. 2007, 26, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Brown, J. Patch use as an indicator of habitat preference, predation risk, and competition. Behav. Ecol. Sociobiol. 1988, 22, 37–47. [Google Scholar] [CrossRef]
- Mogerman, J.E. Zoo Foraging Ecology: Patch Use and Giving-Up Densities as Tools for Animal Care. Ph.D. Thesis, University of Illinois at Chicago, Chicago, IL, USA, 2011. [Google Scholar]
- Troxell-Smith, S.; Watters, J.; Whelan, C.; Brown, J. Zoo foraging ecology: Preference and welfare assessment of two okapi (Okapia Johnstoni) at the Brookfield Zoo. Anim. Behav. Cogn. 2017, 4, 187–199. [Google Scholar] [CrossRef]
- Troxell-Smith, S.M.; Whelan, C.J.; Magle, S.B.; Brown, J.S. Zoo foraging ecology: Development and assessment of a welfare tool for captive animals. Anim. Welf. 2017, 26, 265–275. [Google Scholar] [CrossRef]
- Olsson, O.; Brown, J.S.; Smith, H.G. Long-and short-term state-dependent foraging under predation risk: An indication of habitat quality. Anim. Behav. 2002, 63, 981–989. [Google Scholar] [CrossRef]
- Bolković, M.L.; Caziani, S.M.; Protomastro, J.J. Food habits of the three-banded armadillo (Xenarthra: Dasypodidae) in the Dry Chaco, Argentina. J. Mammal. 1995, 76, 1199–1204. [Google Scholar] [CrossRef]
- Noss, A.; Superina, M.; Abba, A.M. Tolypeutes Matacus. The IUCN Red List of Threatened Species. 2014, p. e.T21974A47443233. Available online: https://dx.doi.org/10.2305/IUCN.UK.2014-1.RLTS.T21974A47443233.en (accessed on 1 February 2023).
- Superina, M.; Miranda, F.; Plese, T.; Vizcaíno, S.; Loughry, W. Maintenance of Xenarthra in Captivity. In The Biology of the Xenarthra; The University Press of Florida: Gainesville, FL, USA, 2008; pp. 232–243. [Google Scholar]
- Bernier, D. Population Analysis and Breeding and Transfer Recommendations: Southern Three-Banded Armadillo (Tolypeutes matacus); AZA Population Management Plan Program: Chicago, IL, USA, 2022. [Google Scholar]
- St. Juliana, J.R.; Kotler, B.P.; Wielebnowski, N.; Cox, J.G. Stress as an adaptation I: Stress hormones are correlated with optimal foraging behaviour of gerbils under the risk of predation. Evol. Ecol. Res. 2017, 18, 571–585. [Google Scholar]
- St. Juliana, J.R.; Kotler, B.P.; Pinshow, B.; Kronfeld-Schor, N. Optimal foraging and physiological responses to the risk of predation: How fecal cortisol concentrations from trapped allenby’s gerbil (Gerbillus andersoni allenbyi) relate to foraging under the risk of predation. Isr. J. Ecol. Evol. 2019, 65, 28–36. [Google Scholar] [CrossRef]
- Sargunaraj, F.; Kotler, B.P.; Juliana, J.R.S.; Wielebnowski, N. Stress as an adaptation II: Does experimental cortisol supplementation affect predation risk assessment in foraging gerbils? Evol. Ecol. Res. 2017, 18, 587–598. [Google Scholar]
- Poessel, S.A.; Biggins, D.E.; Santymire, R.M.; Livieri, T.M.; Crooks, K.R.; Angeloni, L. Environmental enrichment affects adrenocortical stress responses in the endangered black-footed ferret. Gen. Comp. Endocrinol. 2011, 172, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Howell-Stephens, J.A.; Brown, J.S.; Bernier, D.; Mulkerin, D.; Santymire, R.M. Characterizing adrenocortical activity in zoo-housed southern three-banded armadillos (Tolypeutes matacus). Gen. Comp. Endocrinol. 2012, 178, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Loeding, E.; Thomas, J.; Bernier, D.; Santymire, R. Using fecal hormonal and behavioral analyses to evaluate the introduction of two sable antelope at Lincoln Park Zoo. J. Appl. Anim. Welf. Sci. 2011, 14, 220–246. [Google Scholar] [CrossRef]
- Inglis, I.; Shepherd, D.; Smith, P.; Haynes, P.; Bull, D.; Cowan, D.; Whitehead, D. Foraging behaviour of wild rats (Rattus norvegicus) towards new foods and bait containers. Appl. Anim. Behav. Sci. 1996, 47, 175–190. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2022. Available online: https://www.r-project.org (accessed on 8 January 2022).
- Ogle, D.H.; Doll, J.C.; Wheeler, A.P.; Dinno, A. FSA: Simple Fisheries Stock Assessment Methods. 2023, R Package Version 0.9.4. Available online: https://CRAN.R-project.org/package=FSA (accessed on 8 January 2022).
- de Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. 2021, R Package Version 1.3.5. Available online: http://CRAN.R-project.org/package=agricolae (accessed on 8 January 2022).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. Available online: https://ggplot2.tidyverse.org (accessed on 10 January 2022).
- Carlstead, K. Effects of Captivity on the Behavior of Wild Mammals. In Wild Mammals In Captivity—Principles and Techniques; University of Chicago Press: Chicago, IL, USA, 1996; pp. 317–333. [Google Scholar]
- Schmidt, K.A.; Brown, J.S. Patch assessment in fox squirrels: The role of resource density, patch size, and patch boundaries. Am. Nat. 1996, 147, 360–380. [Google Scholar] [CrossRef]
- Abu Baker, M.A.; Brown, J.S. Patch area, substrate depth, and richness affect giving-up densities: A test with mourning doves and cottontail rabbits. Oikos 2009, 118, 1721–1731. [Google Scholar] [CrossRef]
- Valone, T.J.; Brown, J.S. Measuring patch assessment abilities of desert granivores. Ecology 1989, 70, 1800–1810. [Google Scholar] [CrossRef]
- Vizcaíno, S.F.; Loughry, W.J. The Biology of the Xenarthra; University Press of Florida: Gainesville, FL, USA, 2008; ISBN 0-8130-3165-6. [Google Scholar]
- Kotler, B.P.; Brown, J.S. Mechanisms of coexistence of optimal foragers as determinants of local abundances and distributions of desert granivores. J. Mammal. 1999, 80, 361–374. [Google Scholar] [CrossRef]
- Cassini, M.H.; Kacelnik, A.; Segura, E.T. The tale of the screaming hairy armadillo, the guinea pig and the marginal value theorem. Anim. Behav. 1990, 39, 1030–1050. [Google Scholar] [CrossRef]
- Meritt, D.A. Xenarthrans on the Paraguayan Chaco. In The Biology of the Xenarthra; The University Press of Florida: Gainesville, FL, USA, 2008; pp. 294–299. [Google Scholar]
- Vollrath, M. Personality and stress. Scand. J. Psychol. 2001, 42, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Carere, C.; Eens, M. Unravelling animal personalities: How and why individuals consistently differ. Behaviour 2005, 142, 1149–1157. [Google Scholar] [CrossRef]
- Dingemanse, N.J.; Réale, D. Natural selection and animal personality. Behaviour 2005, 142, 1159–1184. [Google Scholar] [CrossRef]
- Réale, D.; Reader, S.M.; Sol, D.; McDougall, P.T.; Dingemanse, N.J. Integrating animal temperament within ecology and evolution. Biol. Rev. 2007, 82, 291–318. [Google Scholar] [CrossRef] [PubMed]
- Baird, B.A.; Kuhar, C.W.; Lukas, K.E.; Amendolagine, L.A.; Fuller, G.A.; Nemet, J.; Willis, M.A.; Schook, M.W. Program animal welfare: Using behavioral and physiological measures to sssess the well-being of animals used for education programs in zoos. Appl. Anim. Behav. Sci. 2016, 176, 150–162. [Google Scholar] [CrossRef]
- Maple, T.L.; Perkins, L.A. Enclosure Furnishings and Structural Environmental Enrichment. In Wild Mammals in Captivity; The University of Chicago Press: Chicago, IL, USA, 1996; pp. 212–222. [Google Scholar]
- Mellen, J.; Sevenich MacPhee, M. Philosophy of environmental enrichment: Past, present, and future. Zoo Biol. 2001, 20, 211–226. [Google Scholar] [CrossRef]
- Swaisgood, R.R.; Ellis, S.; Forthman, D.L.; Shepherdson, D.J. Commentary: Improving well-being for captive giant pandas: Theoretical and practical issues. Zoo Biol. 2003, 22, 347–354. [Google Scholar] [CrossRef]
- Stoinski, T.S.; Daniel, E.; Maple, T.L. A preliminary study of the behavioral effects of feeding enrichment on African elephants. Zoo Biol. 2000, 19, 485–493. [Google Scholar] [CrossRef]
- Bashaw, M.J.; Bloomsmith, M.A.; Marr, M.J.; Maple, T.L. To hunt or not to hunt? A feeding enrichment experiment with captive large felids. Zoo Biol. 2003, 22, 189–198. [Google Scholar] [CrossRef]
- Brown, J.S.; Kotler, B.P. Hazardous duty pay and the foraging cost of predation: Foraging cost of predation. Ecol. Lett. 2004, 7, 999–1014. [Google Scholar] [CrossRef]
- McDonough, C.M.; Loughry, W.J. Behavioral Ecology of Armadillos. In The Biology of the Xenarthra; The University Press of Florida: Gainesville, FL, USA, 2008; pp. 281–293. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Low FGM | High FGM | |
|---|---|---|
| Low GUD | Ideal Low foraging costs and risk; low adrenocortical activity | Positive Low foraging costs; high adrenocortical activity (i.e., eustress) |
| High GUD | Neutral High foraging costs; low adrenocortical activity | Negative High foraging costs; high adrenocortical activity (i.e., distress) |
| ID | Pre | Substrate | Patch | Cover | Post | Overall | |
|---|---|---|---|---|---|---|---|
| Males | 6474 a | 13,130.51 ± 1100.32 | 12,709.31 ± 1434.52 | 18,196.97 ± 4084.78 | 14,757.69 ± 3632.16 | 17,442.22 ± 2644.83 | 14,145.06 ± 946.17 |
| 20200 d | 742.37 ± 46.65 | 1004.24 ± 110.44 | 904.85 ± 97.02 | 843.49 ± 137.89 | 1020.97 ± 188.67 | 848.19 ± 41.27 | |
| 20202 b | 2504.59 ± 324.92 | 2646.27 ± 439.17 | 3451.29 ± 329.20 | 1935.20 ± 301.07 | 1412.37 ± 128.09 | 2457.38 ± 189.83 | |
| Females | 9338 c | 1161.40 ± 80.39 | 1231.13 ± 110.66 | 994.27 ± 79.75 | 1073.29 ± 167.89 | 1061.03 ± 184.70 | 1132.60 ± 51.21 |
| 9717 c | 1112.98 ± 283.66 | 1023.73 ± 160.44 | 1294.27 ± 189.11 | 864.11 ± 124.55 | 1006.01 ± 181.97 | 1119.47 ± 143.70 | |
| 20441 c | 995.49 ± 77.20 | 1903.70 ± 556.53 | 1080.93 ± 173.97 | 1186.18 ± 181.64 | 963.39 ± 134.78 | 1167.09 ± 102.28 | |
| 20727 d | 836.25 ± 38.82 | 788.19 ± 45.86 | 889.48 ± 81.73 | 884.01 ± 97.42 | 636.78 ± 87.26 | 830.79 ± 27.91 | |
| Overall | 3160.04 ± 345.23 | 3152.63 ± 537.41 | 3403.04 ± 805.81 | 3260.93 ± 816.95 | 2802.59 ± 888.20 | – |
| Substrate | Patch | Cover | |||||
|---|---|---|---|---|---|---|---|
| ID | FGM | GUD | FGM | GUD | FGM | GUD | |
| Males | 6474 | 7 | 4 | 7 | 2 | 7 | 5 |
| 20200 | 2 | 3 | 2 | 1 | 1 | 4 | |
| 20202 | 6 | 6 | 6 | 6 | 6 | 6 | |
| Females | 9338 | 4 | 5 | 3 | 5 | 4 | 7 |
| 9717 | 3 | 7 | 5 | 7 | 2 | 3 | |
| 20441 | 5 | 2 | 4 | 4 | 5 | 2 | |
| 20727 | 1 | 1 | 1 | 3 | 3 | 1 | |
| ID | Substrate | Patch | Cover | |
|---|---|---|---|---|
| Males | 6474 | Low | Low | Low |
| 20200 | Low | Low | Low | |
| 20202 * | High | High | Low | |
| Females | 9338 | High | High | High |
| 9717 * | High | High | Low | |
| 20441 | Low | Low | Low | |
| 20727 | Low | Low | Low |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Howell-Stephens, J.; Potratz, E.J.; Brown, J.S.; Bernier, D.; Santymire, R.M. Integrating Measures of Fecal Glucocorticoid Metabolites and Giving-Up Densities to Assess Adrenocortical Activity and Well-Being in Zoo-Housed Three-Banded Armadillos (Tolypeutes matacus). Animals 2023, 13, 1975. https://doi.org/10.3390/ani13121975
Howell-Stephens J, Potratz EJ, Brown JS, Bernier D, Santymire RM. Integrating Measures of Fecal Glucocorticoid Metabolites and Giving-Up Densities to Assess Adrenocortical Activity and Well-Being in Zoo-Housed Three-Banded Armadillos (Tolypeutes matacus). Animals. 2023; 13(12):1975. https://doi.org/10.3390/ani13121975
Chicago/Turabian StyleHowell-Stephens, Jennifer, Emily J. Potratz, Joel S. Brown, Dave Bernier, and Rachel M. Santymire. 2023. "Integrating Measures of Fecal Glucocorticoid Metabolites and Giving-Up Densities to Assess Adrenocortical Activity and Well-Being in Zoo-Housed Three-Banded Armadillos (Tolypeutes matacus)" Animals 13, no. 12: 1975. https://doi.org/10.3390/ani13121975
APA StyleHowell-Stephens, J., Potratz, E. J., Brown, J. S., Bernier, D., & Santymire, R. M. (2023). Integrating Measures of Fecal Glucocorticoid Metabolites and Giving-Up Densities to Assess Adrenocortical Activity and Well-Being in Zoo-Housed Three-Banded Armadillos (Tolypeutes matacus). Animals, 13(12), 1975. https://doi.org/10.3390/ani13121975