
Validation of Enzyme Immunoassays via an Adrenocorticotrophic Stimulation Test for the Non-Invasive Quantification of Stress-Related Hormone Metabolites in Naked Mole-Rats

 ,
,  , and
, and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Saline and ACTH Administration
2.3. Sample Collection
2.4. Fecal Steroid Extraction
2.5. Enzyme Immunoassays
2.6. Data Analysis
3. Results
3.1. EIA Validation
3.2. Dose-Dependent Response in uGCM Concentrations
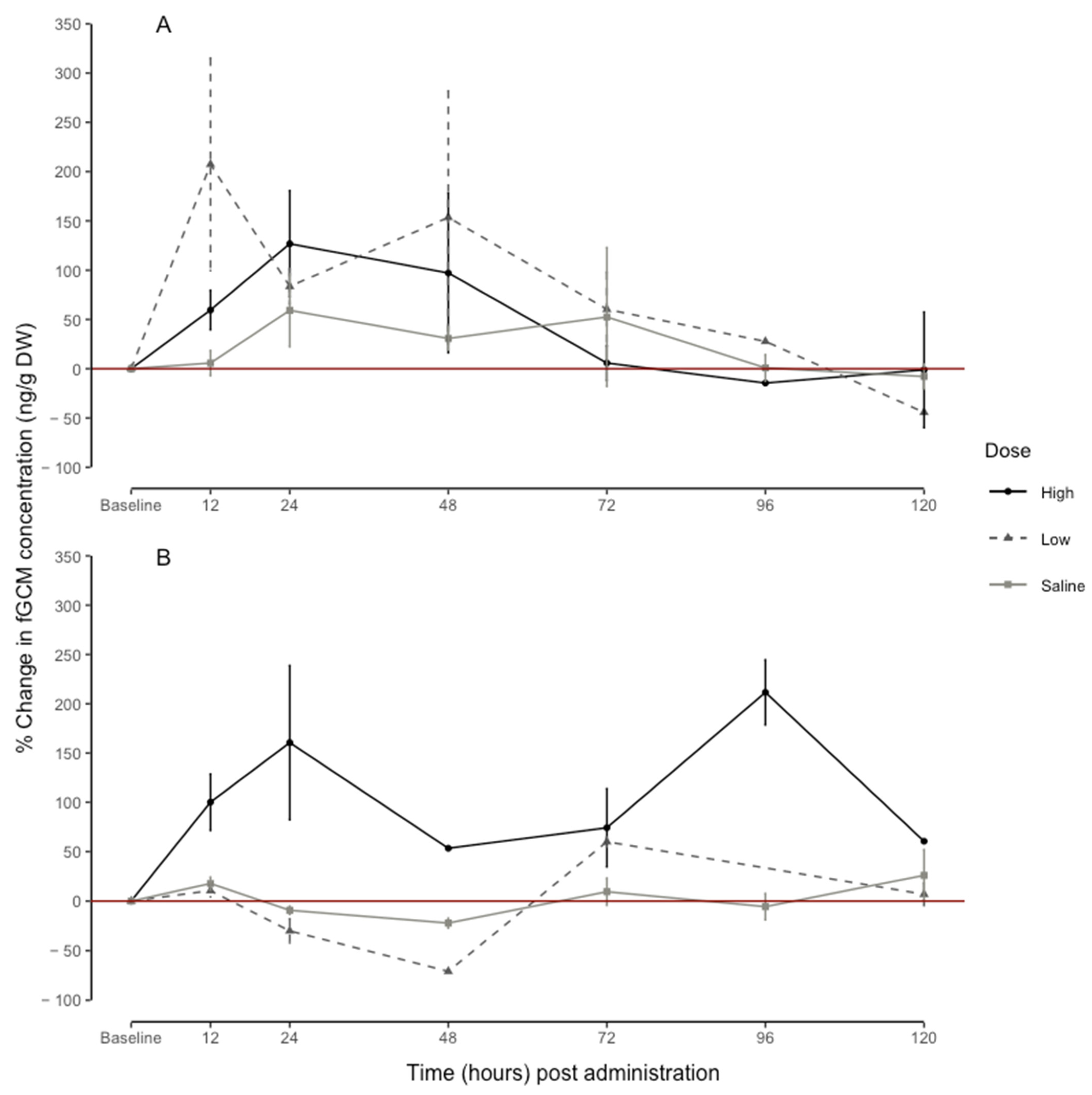
3.3. Dose-Dependent Response in fGCM Concentrations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bennett, N.C.; Faulkes, C.G. African Mole-Rats: Ecology and Eusociality; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Smith, E.S.J.; Schuhmacher, L.-N.; Husson, Z. The naked mole-rat as an animal model in biomedical research: Current perspectives. Anim. Physiol. 2015, 7, 137–148. [Google Scholar] [CrossRef]
- Touma, C.; Palme, R.; Sachser, N. Analyzing corticosterone metabolites in fecal samples of mice: A noninvasive technique to monitor stress hormones. Horm. Behav. 2003, 45, 10–22. [Google Scholar] [CrossRef]
- Selye, H. A Syndrome produced by Diverse Nocuous Agents. Nature 1936, 138, 2. [Google Scholar] [CrossRef]
- Wielebnowski, N. Stress and distress: Evaluating their impact for the well-being of zoo animals. J. Am. Veter-Med. Assoc. 2003, 223, 973–977. [Google Scholar] [CrossRef] [PubMed]
- Sapolsky, R.M.; Romero, L.M.; Munck, A.U. How Do Glucocorticoids Influence Stress Responses? Integrating Permissive, Suppressive, Stimulatory, and Preparative Actions*. Endocr. Rev. 2000, 21, 55–89. [Google Scholar] [CrossRef] [PubMed]
- Palme, R. Non-invasive measurement of glucocorticoids: Advances and problems. Physiol. Behav. 2018, 199, 229–243. [Google Scholar] [CrossRef]
- Schwarzenberger, F.; Möstl, E.; Palme, R.; Bamberg, E. Faecal steroid analysis for non-invasive monitor& of reproductive status in farm, wild and zoo animals. Anim. Reprod. Sci. 1996, 42, 515–526. [Google Scholar]
- Touma, C.; Palme, R. Measuring Fecal Glucocorticoid Metabolites in Mammals and Birds: The Importance of Validation. Ann. N. Y. Acad. Sci. 2005, 1046, 54–74. [Google Scholar] [CrossRef]
- Möstl, E.; Palme, R. Hormones as indicators of stress. Domest. Anim. Endocrinol. 2002, 23, 67–74. [Google Scholar] [CrossRef]
- von der Ohe, C.G.; Servheen, C. Measuring stress in mammals using fecal glucocorticoids: Opportunities and challenges. Wildl. Soc. Bull. 2002, 1215–1225. [Google Scholar]
- Herman, J.P. Neural control of chronic stress adaptation. Front. Behav. Neurosci. 2013, 7, 61. [Google Scholar] [CrossRef] [PubMed]
- Romero, L. Physiological stress in ecology: Lessons from biomedical research. Trends Ecol. Evol. 2004, 19, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Hunt, K.E.; Stimmelmayr, R.; George, C.; Hanns, C.; Suydam, R.S.; Brower, J.H.; Rolland, R.M. Baleen hormones: A novel tool for retrospective assessment of stress and reproduction in bowhead whales (Balaena mysticetus). Conserv. Physiol. 2014, 2, cou030. [Google Scholar] [CrossRef] [PubMed]
- Ping, X.; Liu, N.; Jiang, Z.; Xu, H.; Bai, J.; Zhang, S.; Zhong, Z.; Li, C. Assaying progesterone, estradiol and cortisol concentra-tions in hair of Père David deer hinds: An alternative way to reflect seasonality of steroid secretion. Biol. Rhythm. Res. 2018, 49, 422–430. [Google Scholar] [CrossRef]
- Edwards, P.D.; Mooney, S.J.; Bosson, C.O.; Toor, I.; Palme, R.; Holmes, M.M.; Boonstra, R. The stress of being alone: Removal from the colony, but not social subordination, increases fecal cortisol metabolite levels in eusocial naked mole-rats. Horm. Behav. 2020, 121, 104720. [Google Scholar] [CrossRef]
- Kamgang, V.W.; Bennett, N.C.; van der Goot, A.C.; Majelantle, T.L.; Ganswindt, A. Patterns of faecal glucocorticoid me-tabolite levels in captive roan antelope (Hippotragus equinus) in relation to reproductive status and season. Gen. Comp. Endocrinol. 2022, 325, 114052. [Google Scholar] [CrossRef]
- Son, Y.L.; Ubuka, T.; Tsutsui, K. Regulation of stress response on the hypothalamic-pituitary-gonadal axis via gonadotro-pin-inhibitory hormone. Front. Neuroendocrinol. 2022, 64, 100953. [Google Scholar] [CrossRef]
- Majelantle, T.L.; Mcintyre, T.; Ganswindt, A. Monitoring the effects of land transformation on African clawless otters (Aonyx capensis) using fecal glucocorticoid metabolite concentrations as a measure of stress. Integr. Zool. 2020, 15, 293–306. [Google Scholar] [CrossRef]
- Ramahlo, M.; Chimimba, C.; Pirk, C.; Ganswindt, A. Non-invasive monitoring of adrenocortical activity in free-ranging Namaqua rock mice Micaelamys namaquensis from South Africa in response to anthropogenic land use and season. Wildl. Biol. 2019, 2019, 1–6. [Google Scholar] [CrossRef]
- Crossey, B.; Chimimba, C.; du Plessis, C.; Hall, G.; Ganswindt, A. Using faecal glucocorticoid metabolite analyses to elucidate stressors of African wild dogs Lycaon pictus from South Africa. Wildl. Biol. 2020, 2020, 1. [Google Scholar] [CrossRef]
- Goymann, W. On the use of non-invasive hormone research in uncontrolled, natural environments: The problem with sex, diet, metabolic rate and the individual. Methods Ecol. Evol. 2012, 3, 757–765. [Google Scholar] [CrossRef]
- Santamaria, F.; Barlow, C.K.; Schlagloth, R.; Schittenhelm, R.B.; Palme, R.; Henning, J. Identification of Koala (Phascolarctos cinereus) Faecal Cortisol Metabolites Using Liquid Chromatography-Mass Spectrometry and Enzyme Immunoassays. Metabolites 2021, 11, 393. [Google Scholar] [CrossRef] [PubMed]
- Chelini, M.-O.M.; Otta, E.; Yamakita, C.; Palme, R. Sex differences in the excretion of fecal glucocorticoid metabolites in the Syrian hamster. J. Comp. Physiol. B 2010, 180, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, F.; Schlagloth, R.; Valenza, L.; Palme, R.; de Villiers, D.; Henning, J. The Effect of Disease and Injury on Faecal Cortisol Metabolites, as an Indicator of Stress in Wild Hospitalised Koalas, Endangered Australian Marsupials. Veter-Sci. 2023, 10, 65. [Google Scholar] [CrossRef]
- Ganswindt, A.; Tordiffe, A.S.W.; Stam, E.; Howitt, M.J.; Jori, F. Determining adrenocortical activity as a measure of stress in African buffalo (Syncerus caffer) based on faecal analysis. African Zool. 2012, 47, 261–269. [Google Scholar] [CrossRef]
- O’Riain, M.J.; Jarvis, J.U.M.; Faulkes, C.G. A dispersive morph in the naked mole-rat. Nature 1996, 380, 619–621. [Google Scholar] [CrossRef] [PubMed]
- Bennett, N.C.; Jarvis, J.U.M. Coefficients of digestibility and nutritional values of geophytes and tubers eaten by southern African mole-rats (Rodentia: Bathyergidae). J. Zool. 1995, 236, 189–198. [Google Scholar] [CrossRef]
- Fraňková, M.; Palme, R.; Frynta, D. Family affairs and experimental male replacement affect fecal glucocorticoid metabolites levels in the Egyptian spiny mouse Acomys cahirinus. Zool. Stud. 2012, 51, 277–287. [Google Scholar]
- Palme, R.; Touma, C.; Arias, N.; Dominchin, M.F.; Lepschy, M. Steroid extraction: Get the best out of faecal samples. Vet. Med. Austria. 2013, 100, 238–246. [Google Scholar]
- Millspaugh, J.J.; Washburn, B.E. Use of fecal glucocorticoid metabolite measures in conservation biology research: Con-siderations for application and interpretation. Gen. Comp. Endocrinol. 2004, 138, 189–199. [Google Scholar] [CrossRef]
- Ganswindt, A.; Münscher, S.; Henley, M.; Palme, R.; Thompson, P.; Bertschinger, H. Concentrations of faecal glucocorticoid metabolites in physically injured free-ranging African elephants Loxodonta africana. Wildl. Biol. 2010, 16, 323–332. [Google Scholar] [CrossRef]
- Ganswindt, A.; Heistermann, M.; Borragan, S.; Hodges, J.K. Assessment of testicular endocrine function in captive African elephants by measurement of urinary and fecal androgens. Zoo Biol. 2002, 21, 27–36. [Google Scholar] [CrossRef]
- Palme, R.; Mostl, E. Measurement of cortisol metabolites in feces of sheep as a parameter of cortisol concentration in blood. Mammal Biol. 1997, 2, 192–197. [Google Scholar]
- Möstl, E.; Maggs, J.L.; Schrötter, G.; Besenfelder, U.; Palme, R. Measurement of cortisol metabolites in feces of ruminants. Vet. Res. Commun. 2002, 26, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Touma, C.; Sachser, N.; Möstl, E.; Palme, R. Effects of sex and time of day on metabolism and excretion of corticosterone in urine and feces of mice. Gen. Comp. Endocrinol. 2003, 130, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Folin, O.; Morris, J. On the determination of creatinine and creatine in urine. J. Biol. Chem. 1914, 17, 469–473. [Google Scholar] [CrossRef]
- Jackson, C.R.; Bennett, N.C. Is the natal mole-rat (Cryptomys hottentotus natalensis) a spontaneous or induced ovulator? J. Mammal. 2005, 86, 1–6. [Google Scholar] [CrossRef]
- Bates, D.M.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; ISBN 3-900051-07-0. Available online: http://www.R-project.org/ (accessed on 23 June 2022).
- Palme, R.; Robia, C.; Messmann, S.; Hofer, J.; Möstl, E. Measurement of faecal cortisol metabolites in ruminants: A non-invasive parameter of adrenocortical function. Wien. Tierarztl. Monatsschr. 1999, 86, 237–241. [Google Scholar]
- Medger, K.; Bennett, N.C.; Prins, A.; Lutermann, H.; Ganswindt, A. Sex and dose-dependent responses of urinary and faecal glucocorticoid metabolite concentrations following an ACTH challenge in eastern rock sengis (Elephantulus myurus). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 245, 110696. [Google Scholar] [CrossRef]
- Crill, C.; Janz, D.M.; Kusch, J.M.; Santymire, R.M.; Heyer, G.P.; Shury, T.K.; Lane, J.E. Investigation of the utility of feces and hair as non-invasive measures of glucocorticoids in wild black-tailed prairie dogs (Cynomys ludovicianus). Gen. Comp. Endocrinol. 2019, 275, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Tilbrook, A.; Turner, A.; Clarke, I. Effects of stress on reproduction in non-rodent mammals: The role of glucocorticoids and sex differences. Rev. Reprod. 2000, 5, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Pizzutto, C.S.; Sgai, M.G.F.G.; Francischini, C.P.; Viau, P.; De Oliveira, C.A.; Guimarães, M.A.D.B.V. Physiological validation of enzyme immunoassay of fecal glucocorticoid metabolite levels and diurnal variation measured in captive Black-tufted Marmoset Callithrix penicillata (Mammalia: Primates: Callitrichidae). J. Threat. Taxa 2015, 7, 7234–7242. [Google Scholar] [CrossRef]
- Lepschy, M.; Touma, C.; Hruby, R.; Palme, R. Non-invasive measurement of adrenocortical activity in male and female rats. Lab. Anim. 2007, 41, 372–387. [Google Scholar] [CrossRef]
- Sipari, S.; Ylönen, H.; Palme, R. Excretion and measurement of corticosterone and testosterone metabolites in bank voles (Myodes glareolus). Gen. Comp. Endocrinol. 2017, 243, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Reeve, H.K.; Sherman, P.W. Intracolonial Aggression and Nepotism by the Breeding Female Naked Mole-Rat. In The Biology of the Naked Mole-Rat; Princeton University Press: Princeton, NJ, USA, 2017; pp. 337–357. [Google Scholar] [CrossRef]
- Palme, R.; Fischer, P.; Schildorfer, H.; Ismail, M.N. Excretion of infused 14C-steroid hormones via feces and urine in domestic livestock. Anim. Reprod. Sci. 1996, 43, 43–63. [Google Scholar] [CrossRef]
- Nováková, M.; Palme, R.; Kutalová, H.; Janský, L.; Frynta, D. The effects of sex, age and commensal way of life on levels of fecal glucocorticoid metabolites in spiny mice (Acomys cahirinus). Physiol. Behav. 2008, 95, 187–193. [Google Scholar] [CrossRef]
- Edwards, P.D.; Dean, E.K.; Palme, R.; Boonstra, R. Assessing space use in meadow voles: The relationship to reproduction and the stress axis. J. Mammal. 2019, 100, 4–12. [Google Scholar] [CrossRef]
- Buffenstein, R.; Park, T.; Hanes, M.; Artwohl, J.E. Naked Mole Rat. The Laboratory Rabbit, Guinea Pig, Hamster, and Other Rodents; Elsevier: Amsterdam, The Netherlands, 2012; pp. 1055–1074. [Google Scholar]
- Medger, K. Stress in an underground empire. Biol. Lett. 2022, 18, 20220012. [Google Scholar] [CrossRef]
- Edwards, P.D. For the physiology of cooperative breeding, it’s time to move beyond stress: A Comment on:‘Stress in an un-derground empire’(2022) by Medger. Biol. Lett. 2022, 18, 20220375. [Google Scholar] [CrossRef]
- Hart, D.W.; Bennett, N.C.; Voigt, C. Social stress is unlikely to play a major role in reproductive suppression of female subordinate naked mole-rats and Damaraland mole-rats. Biol. Lett. 2022, 18, 20220292. [Google Scholar] [CrossRef] [PubMed]
- Blecher, A.S.; Bennett, N.C.; Medger, K.; Hagenah, N.; Ganswindt, A.; Oosthuizen, M.K. Effect of colony disruption and social isolation on naked mole-rat endocrine correlates. Gen. Comp. Endocrinol. 2020, 295, 113520. [Google Scholar] [CrossRef]
- Wassif, W.S.; McLoughlin, D.M.; Vincent, R.P.; Conroy, S.; Russell, G.F.; Taylor, N.F. Steroid metabolism and excretion in severe anorexia nervosa: Effects of refeeding. Am. J. Clin. Nutr. 2011, 93, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Lewicka, S.; Nowicki, M.; Vecsei, P. Effect of sodium restriction on urinary excretion of cortisol and its metabolites in humans. Steroids 1998, 63, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Baudrand, R.; Campino, C.; Carvajal, C.A.; Olivieri, O.; Guidi, G.; Faccini, G.; Vöhringer, P.A.; Cerda, J.; Owen, G.; Kalergis, A.M.; et al. High sodium intake is associated with increased glucocorticoid production, insulin resistance and metabolic syndrome. Clin. Endocrinol. 2014, 80, 677–684. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dose | ID | EIA | ||||
|---|---|---|---|---|---|---|
| Cortisol | Corticosterone | 5α-Pregnane | Oxoaetio | Oxoaetio | ||
| Cholanolone I | Cholanolone II | |||||
| Low | F1 | −42 | −7 | 40 ^ | 174 ^ | 11 |
| F2 | 60 ^ | −23 * | 46 * | 26 * | 38 * | |
| F3 | 325 | 193 | 19 | 130 * | −23 * | |
| High | F5 | 29 | −13 | −15 | −5 | 170 |
| F6 | 106 * | 105 * | −2 * | 28 * | 71 * | |
| Low | M1 | 17 ^ | 69 ^ | 115 ^ | −5 ^ | 26 |
| M2 | 4 * | −58 * | 131 * | 88 * | 88 * | |
| M3 | 1545 ^ | 457 ^ | 338 ^ | 424 ^ | 367 ^ | |
| High | M4 | 261 | 91 | 295 ^ | 73 ^ | 71 |
| M5 | 995 ^ | 612 ^ | 113 ^ | 91 ^ | 90 ^ | |
| Dose | ID | EIA | ||||
|---|---|---|---|---|---|---|
| Cortisol | Corticosterone | 5α-Pregnane | Oxoaetio | Oxoaetio | ||
| Cholanolone I | Cholanolone II | |||||
| Low | F1 | 36 * | 36 * | −10 * | 76 * | −2 * |
| F2 | 38 | 73 * | 83 | 195 | 218 * | |
| F3 | 29 * | −1 * | −12 * | 64 * | −8 * | |
| High | F4 | 79 | 16 | 13 | 392 | −5 |
| F5 | 74 | 27 * | 19 | 160 | 38 | |
| F6 | 73 | 110 | 120 * | 233 * | −29 | |
| Low | M1 | 230 | 77 | 7 | 315 | −1 * |
| M2 | 166 | 64 | 87 | 172 * | 97 | |
| M3 | 54 | 8 | −5 | 65 | 27 | |
| High | M4 | 56 * | 80 | 40 | 309 | 9 * |
| M5 | 99 | 94 | *64 | 147 | 68 * | |
| M6 | 296 | 92 | 39 | 274 | 72 | |
| Species | Dose | EIA | Citation |
|---|---|---|---|
| Cattle (Bos taurus taurus) | 0.25 mg, 0.5 mg, and 1 mg | 11-oxoetiocholanolone I | [41] |
| Mice (Mus musculus f. domesticus) | 20 μg and 60 μg/100 g | 5α-pregnane-3β,11β,21-triol-20-one EIA | [3] |
| Black-tailed prairie dogs (Cynomys ludovicianus) | * 4 μg and 12 μg/100 g | Cortisol- and Corticosterone- EIAs | [43] |
| Eastern rock sengis (Elephantulus myurus) | ` 20 μg and 60 μg/100 g | 5α-pregnane-3β,11β,21-triol-20-one EIA | [42] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majelantle, T.L.; Bennett, N.C.; Ganswindt, S.B.; Hart, D.W.; Ganswindt, A. Validation of Enzyme Immunoassays via an Adrenocorticotrophic Stimulation Test for the Non-Invasive Quantification of Stress-Related Hormone Metabolites in Naked Mole-Rats. Animals 2023, 13, 1424. https://doi.org/10.3390/ani13081424
Majelantle TL, Bennett NC, Ganswindt SB, Hart DW, Ganswindt A. Validation of Enzyme Immunoassays via an Adrenocorticotrophic Stimulation Test for the Non-Invasive Quantification of Stress-Related Hormone Metabolites in Naked Mole-Rats. Animals. 2023; 13(8):1424. https://doi.org/10.3390/ani13081424
Chicago/Turabian StyleMajelantle, Tshepiso Lesedi, Nigel Charles Bennett, Stefanie Birgit Ganswindt, Daniel William Hart, and Andre Ganswindt. 2023. "Validation of Enzyme Immunoassays via an Adrenocorticotrophic Stimulation Test for the Non-Invasive Quantification of Stress-Related Hormone Metabolites in Naked Mole-Rats" Animals 13, no. 8: 1424. https://doi.org/10.3390/ani13081424
APA StyleMajelantle, T. L., Bennett, N. C., Ganswindt, S. B., Hart, D. W., & Ganswindt, A. (2023). Validation of Enzyme Immunoassays via an Adrenocorticotrophic Stimulation Test for the Non-Invasive Quantification of Stress-Related Hormone Metabolites in Naked Mole-Rats. Animals, 13(8), 1424. https://doi.org/10.3390/ani13081424