
RNA-Seq Study on the Longissimus thoracis Muscle of Italian Large White Pigs Fed Extruded Linseed with or without Antioxidants and Polyphenols
 , , ,
, , ,  , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
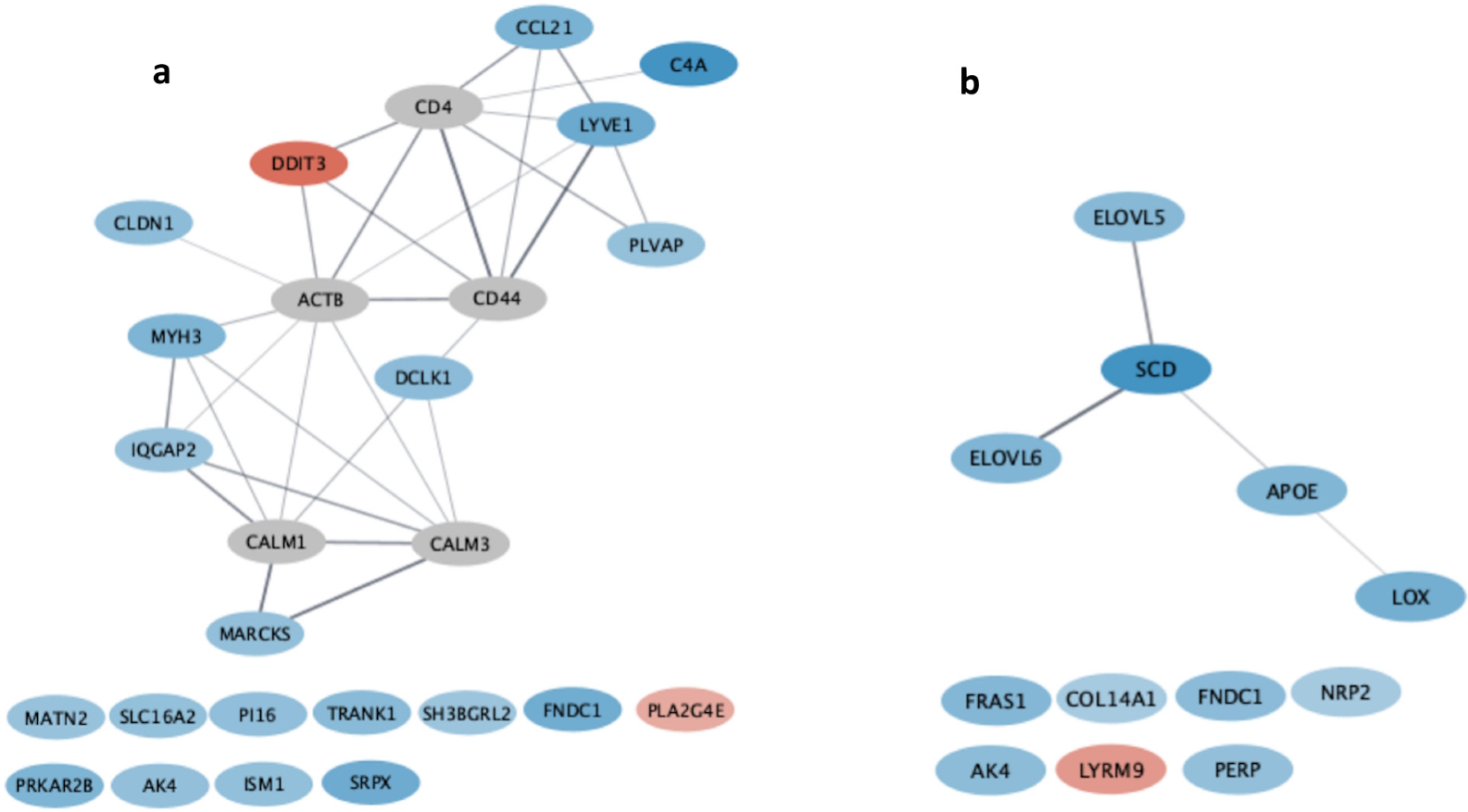
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rocchetti, G.; Vitali, M.; Zappaterra, M.; Righetti, L.; Sirri, R.; Lucini, L.; Dall’Asta, C.; Davoli, R.; Galaverna, G. A molecular insight into the lipid changes of pig Longissimus thoracis muscle following dietary supplementation with functional ingredients. PLoS ONE 2022, 17, e0264953. [Google Scholar] [CrossRef] [PubMed]
- Vitali, M.; Sirri, R.; Zappaterra, M.; Zambonelli, P.; Giannini, G.; Lo Fiego, D.P.; Davoli, R. Functional analysis finds differences in the muscle transcriptome of pigs fed an n-3 PUFA-enriched diet with or without antioxidant supplementations. PLoS ONE 2019, 14, e0212449. [Google Scholar] [CrossRef] [PubMed]
- ANAS. Available online: https://www.anas.it/ (accessed on 29 January 2023).
- Sirri, R.; Vitali, M.; Zambonelli, P.; Giannini, G.; Zappaterra, M.; Lo Fiego, D.P.; Sami, D.; Davoli, R. Effect of diets supplemented with linseed alone or combined with vitamin E and selenium or with plant extracts, on Longissimus thoracis transcriptome in growing-finishing Italian Large White pigs. J. Anim. Sci. Biotechnol. 2018, 9, 81. [Google Scholar] [CrossRef] [PubMed]
- ArrayExpress. Available online: https://www.ebi.ac.uk/biostudies/arrayexpress (accessed on 29 January 2023).
- FastQC v.0.11.5. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 29 January 2023).
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef]
- Trimmomatic v.0.39. Available online: http://www.usadellab.org/cms/?page=trimmomatic (accessed on 29 January 2023).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Sus Scrofa Genome Assembly Version Sscrofa11.1. Available online: https://www.ensembl.org/Sus_scrofa/Info/Index (accessed on 29 January 2023).
- STAR v.2.6.1.d. Available online: https://github.com/alexdobin/STAR (accessed on 29 January 2023).
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Cunningham, F.; Allen, J.E.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Bennett, R.; et al. Ensembl 2022. Nucleic Acids Res. 2022, 50, D988–D995. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Sva. Available online: https://bioconductor.org/packages/3.14/bioc/html/sva.html (accessed on 29 January 2023).
- Parker, H.S.; Corrada Bravo, H.; Leek, J.T. Removing batch effects for prediction problems with frozen surrogate variable analysis. PeerJ 2014, 2, e561. [Google Scholar] [CrossRef]
- Chen, Y.; Lun, A.T.L.; Smyth, G.K. From reads to genes to pathways: Differential expression analysis of RNA-Seq experiments using Rsubread and the edgeR quasi-likelihood pipeline. F1000Research 2016, 5, 1438. [Google Scholar] [CrossRef] [PubMed]
- Markussen, L.K.; Rondini, E.A.; Johansen, O.; Madsen, J.; Sustarsic, E.G.; Marcher, A.; Hansen, J.B.; Gerhart-Hines, Z.; Granneman, J.G.; Mandrup, S. Lipolysis regulates major transcriptional programs in brown adipocytes. Nat. Commun. 2022, 13, 3956. [Google Scholar] [CrossRef] [PubMed]
- BioMart-Ensembl. Available online: https://www.ensembl.org/biomart/martview/ (accessed on 29 January 2023).
- Doncheva, N.T.; Morris, J.H.; Gorodkin, J.; Jensen, L.J. Cytoscape StringApp: Network Analysis and Visualization of Proteomics Data. J. Proteome Res. 2019, 18, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Shu, Z.; Wang, L.; Wang, J.; Zhang, L.; Hou, X.; Yan, H.; Wang, L. Integrative Analysis of Nanopore and Illumina Sequencing Reveals Alternative Splicing Complexity in Pig Longissimus Dorsi Muscle. Front. Genet. 2022, 13, 877646. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, M. Complement C4, Infections, and Autoimmune Diseases. Front. Immunol. 2021, 12, 694928. [Google Scholar] [CrossRef]
- Christopherson, K.W.; Campbell, J.J.; Travers, J.B.; Hromas, R.A. Low-Molecular-Weight Heparins Inhibit CCL21-Induced T Cell Adhesion and Migration. J. Pharmacol. Exp. Ther. 2002, 302, 290–295. [Google Scholar] [CrossRef]
- Lawrance, W.; Banerji, S.; Day, A.J.; Bhattacharjee, S.; Jackson, D.G. Binding of Hyaluronan to the Native Lymphatic Vessel Endothelial Receptor LYVE-1 Is Critically Dependent on Receptor Clustering and Hyaluronan Organization. J. Biol. Chem. 2016, 291, 8014–8030. [Google Scholar] [CrossRef]
- Johnson, L.A.; Banerji, S.; Lawrance, W.; Gileadi, U.; Prota, G.; Holder, K.A.; Roshorm, Y.M.; Hanke, T.; Cerundolo, V.; Gale, N.W.; et al. Dendritic cells enter lymph vessels by hyaluronan-mediated docking to the endothelial receptor LYVE-1. Nat. Immunol. 2017, 18, 762–770. [Google Scholar] [CrossRef]
- Guo, L.; Zhang, H.; Hou, Y.; Wei, T.; Liu, J. Plasmalemma vesicle-associated protein: A crucial component of vascular homeostasis. Exp. Ther. Med. 2016, 12, 1639–1644. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes: From molecules to man. Biochem. Soc. Trans. 2017, 45, 1105–1115. [Google Scholar] [CrossRef]
- Guillou, H.; Zadravec, D.; Martin, P.G.P.; Jacobsson, A. The key roles of elongases and desaturases in mammalian fatty acid metabolism: Insights from transgenic mice. Prog. Lipid Res. 2010, 49, 186–199. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yu, L.; Schmidt, R.E.; Su, C.; Huang, X.; Gould, K.; Cao, G. Characterization of HSCD5, a novel human stearoyl-CoA desaturase unique to primates. Biochem. Biophys. Res. Commun. 2005, 332, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Goren, M.A.; Fox, B.G. Wheat germ cell-free translation, purification, and assembly of a functional human stearoyl-CoA desaturase complex. Protein Expr. Purif. 2008, 62, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Rosenblat, M.; Volkova, N.; Coleman, R.; Aviram, M. Pomegranate Byproduct Administration to Apolipoprotein E-Deficient Mice Attenuates Atherosclerosis Development as a Result of Decreased Macrophage Oxidative Stress and Reduced Cellular Uptake of Oxidized Low-Density Lipoprotein. J. Agric. Food Chem. 2006, 54, 1928–1935. [Google Scholar] [CrossRef] [PubMed]
- Aviram, M.; Volkova, N.; Coleman, R.; Dreher, M.; Reddy, M.K.; Ferreira, D.; Rosenblat, M. Pomegranate Phenolics from the Peels, Arils, and Flowers Are Antiatherogenic: Studies in Vivo in Atherosclerotic Apolipoprotein E-Deficient (E0) Mice and in Vitro in Cultured Macrophages and Lipoproteins. J. Agric. Food Chem. 2008, 56, 1148–1157. [Google Scholar] [CrossRef] [PubMed]
- Ranucci, D.; Beghelli, D.; Trabalza-Marinucci, M.; Branciari, R.; Forte, C.; Olivieri, O.; Badillo Pazmay, G.V.; Cavallucci, C.; Acuti, G. Dietary effects of a mix derived from oregano (Origanum vulgare L.) essential oil and sweet chestnut (Castanea sativa Mill.) wood extract on pig performance, oxidative status and pork quality traits. Meat Sci. 2015, 100, 319–326. [Google Scholar] [CrossRef]
- Zou, Y.; Xiang, Q.; Wang, J.; Wei, H.; Peng, J. Effects of oregano essential oil or quercetin supplementation on body weight loss, carcass characteristics, meat quality and antioxidant status in finishing pigs under transport stress. Livest. Sci. 2016, 192, 33–38. [Google Scholar] [CrossRef]
- Ishihara, T.; Yoshida, M.; Arita, M. Omega-3 fatty acid-derived mediators that control inflammation and tissue homeostasis. Int. Immunol. 2019, 31, 559–567. [Google Scholar] [CrossRef]
- Schurch, N.J.; Schofield, P.; Gierliński, M.; Cole, C.; Sherstnev, A.; Singh, V.; Wroblel, N.; Gharbi, K.; Simpson, G.G.; Owe-Hughes, T.; et al. How many biological replicates are needed in an RNA-seq experiment and which differential expression tool should you use? RNA 2016, 22, 839–851. [Google Scholar] [CrossRef]
- de Tonnac, A.; Guillevic, M.; Mourot, J. Fatty acid composition of several muscles and adipose tissues of pigs fed n-3 PUFA rich diets. Meat Sci. 2018, 140, 1–8. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Sample | Raw Reads (N) | Clean Reads (N) | Uniquely Mapped Reads (%) | Reads Assigned (N) | Reads Assigned (%) |
|---|---|---|---|---|---|
| ERR2775643 | 15,222,091 | 12,231,608 | 86.9 | 9,139,962 | 74.7 |
| ERR2775644 | 11,864,858 | 9,324,171 | 87.3 | 6,775,878 | 72.7 |
| ERR2775645 | 19,689,337 | 15,667,592 | 85.9 | 11,375,307 | 72.6 |
| ERR2775646 | 20,182,659 | 16,184,879 | 86.2 | 11,585,338 | 71.6 |
| ERR2775647 | 12,477,450 | 9,885,484 | 86.2 | 7,077,650 | 71.6 |
| ERR2775648 | 12,974,371 | 9,985,754 | 87.3 | 7,270,811 | 72.8 |
| ERR2775649 | 12,534,078 | 9,874,602 | 85.9 | 7,131,936 | 72.2 |
| ERR2775650 | 13,908,762 | 10,890,036 | 85.9 | 7,959,151 | 73.1 |
| ERR2775651 | 15,976,939 | 12,860,711 | 86.5 | 9,412,286 | 73.2 |
| ERR2775652 | 13,209,059 | 10,380,751 | 87.8 | 7,545,142 | 72.7 |
| ERR2775653 | 16,449,679 | 13,227,601 | 85.9 | 9,420,488 | 71.2 |
| ERR2775654 | 17,059,831 | 13,680,926 | 85.9 | 9,809,351 | 71.7 |
| ERR2775655 | 17,665,490 | 14,373,661 | 86.2 | 10,242,721 | 71.3 |
| ERR2775656 | 19,899,598 | 16,125,093 | 85.9 | 11,299,711 | 70.1 |
| ERR2775657 | 15,916,725 | 12,643,514 | 86.1 | 8,989,816 | 71.1 |
| ERR2775658 | 11,677,680 | 9,152,447 | 85.9 | 6,543,642 | 71.5 |
| ERR2775659 | 15,434,949 | 12,447,467 | 86.4 | 9,135,322 | 73.4 |
| ERR2775660 | 17,757,331 | 14,217,671 | 85.5 | 10,410,825 | 73.2 |
| ERR2775661 | 13,390,239 | 10,536,701 | 87.0 | 7,571,975 | 71.9 |
| ERR2775662 | 16,184,027 | 12,817,542 | 85.7 | 9,264,611 | 72.3 |
| ERR2775663 | 14,471,427 | 11,460,053 | 86.8 | 8,240,029 | 71.9 |
| ERR2775664 | 17,320,618 | 13,933,143 | 85.7 | 10,027,479 | 72.0 |
| ERR2775665 | 11,995,565 | 9,496,805 | 85.5 | 6,876,554 | 72.4 |
| ERR2775666 | 17,203,151 | 13,983,413 | 86.9 | 10,233,830 | 73.2 |
| ERR2775667 | 14,287,420 | 11,591,105 | 86.9 | 8,529,950 | 73.6 |
| ERR2775668 | 17,274,474 | 13,799,136 | 85.9 | 9,852,763 | 71.4 |
| ERR2775669 | 16,786,939 | 13,525,029 | 86.3 | 9,802,873 | 72.5 |
| ERR2775670 | 15,204,776 | 12,248,050 | 86.4 | 9,032,702 | 73.7 |
| ERR2775671 | 11,461,974 | 9,100,759 | 84.9 | 6,509,055 | 71.5 |
| ERR2775672 | 15,301,870 | 12,092,472 | 86.6 | 8,836,322 | 73.1 |
| ERR2775673 | 12,247,350 | 9,678,899 | 86.2 | 6,823,627 | 70.5 |
| ERR2775674 | 12,718,558 | 10,018,047 | 86.7 | 7,270,784 | 72.5 |
| ERR2775675 | 13,279,182 | 10,362,507 | 86.9 | 7,643,066 | 73.7 |
| ERR2775676 | 14,546,288 | 11,620,043 | 86.3 | 8,331,936 | 71.7 |
| ERR2775677 | 17,174,540 | 13,807,456 | 86.3 | 9,910,284 | 71.8 |
| ERR2775678 | 17,414,226 | 14,078,405 | 86.8 | 10,294,253 | 73.1 |
| ERR2775679 | 17,246,132 | 13,643,475 | 86.4 | 9,756,837 | 71.5 |
| ERR2775680 | 11,403,686 | 9,015,980 | 85.8 | 6,383,622 | 70.8 |
| ERR2775681 | 15,452,512 | 12,419,639 | 86.5 | 8,957,969 | 72.1 |
| ERR2775682 | 17,533,837 | 14,169,711 | 86.4 | 10,330,258 | 72.9 |
| ERR2775683 | 13,905,859 | 11,119,814 | 86.6 | 8,117,696 | 73.0 |
| ERR2775684 | 14,116,063 | 11,330,645 | 87.1 | 8,172,131 | 72.1 |
| ERR2775685 | 10,588,464 | 8,189,077 | 86.3 | 5,835,669 | 71.3 |
| ERR2775686 | 16,052,676 | 12,844,838 | 86.3 | 9,253,262 | 72.0 |
| ERR2775687 | 11,974,411 | 9,290,605 | 85.9 | 6,610,776 | 71.2 |
| ERR2775688 | 18,407,201 | 14,873,971 | 86.3 | 10,764,983 | 72.4 |
| ERR2775689 | 13,247,963 | 10,477,587 | 86.3 | 7,505,028 | 71.6 |
| ERR2775690 | 13,937,437 | 11,134,910 | 87.3 | 8,087,906 | 72.6 |
| D1 vs. D3 | ||||||
| ENSEMBL ID | D1 | D3 | log2FC | p-value | padj | Gene Symbol |
| ENSSSCG00000001427 | 51.66 | 129.03 | −1.44 | 1.19 × 10−4 | 0.034 | C4A |
| ENSSSCG00000029886 | 14.61 | 32.98 | −1.12 | 1.75 × 10−4 | 0.039 | LYVE1 |
| ENSSSCG00000012234 | 16.32 | 30.25 | −1.09 | 2.03 × 10−5 | 0.014 | SRPX |
| ENSSSCG00000004052 | 48.08 | 96.87 | −1.07 | 3.30 × 10−6 | 0.012 | FNDC1 |
| ENSSSCG00000038162 | 31.58 | 59.87 | −1.00 | 8.73 × 10−5 | 0.031 | CCL21 |
| ENSSSCG00000037706 | 12.59 | 26.32 | −0.97 | 1.90 × 10−5 | 0.014 | PRKAR2B |
| ENSSSCG00000018007 | 143.72 | 284.16 | −0.97 | 4.30 × 10−4 | 0.064 | MYH3 |
| ENSSSCG00000011239 | 15.31 | 27.81 | −0.88 | 5.14 × 10−4 | 0.071 | TRANK1 |
| ENSSSCG00000033919 | 53.23 | 91.72 | −0.86 | 9.27 × 10−4 | 0.099 | DCLK1 |
| ENSSSCG00000040904 | 19.62 | 35.12 | −0.86 | 1.39 × 10−5 | 0.014 | CLDN1 |
| ENSSSCG00000001570 | 235.46 | 402.93 | −0.82 | 3.32 × 10−4 | 0.059 | PI16 |
| ENSSSCG00000007073 | 10.58 | 18.38 | −0.81 | 5.45 × 10−4 | 0.074 | ISM1 |
| ENSSSCG00000037803 | 98.72 | 184.50 | −0.80 | 6.47 × 10−4 | 0.080 | MARCKS |
| ENSSSCG00000040337 | 49.27 | 83.66 | −0.80 | 5.12 × 10−4 | 0.071 | AK4 |
| ENSSSCG00000029458 | 16.47 | 27.36 | −0.80 | 3.95 × 10−5 | 0.019 | SLC16A2 |
| ENSSSCG00000037025 | 76.02 | 123.59 | −0.79 | 3.41 × 10−5 | 0.017 | PLVAP |
| ENSSSCG00000014088 | 24.90 | 43.19 | −0.76 | 3.18 × 10−4 | 0.059 | IQGAP2 |
| ENSSSCG00000006082 | 86.84 | 152.88 | −0.76 | 1.06 × 10−4 | 0.033 | MATN2 |
| ENSSSCG00000032015 | 12.79 | 21.14 | −0.73 | 3.64 × 10−4 | 0.061 | SH3BGRL2 |
| ENSSSCG00000037292 | 23.96 | 13.65 | 0.74 | 7.61 × 10−4 | 0.089 | PLA2G4E |
| ENSSSCG00000039921 | 18.60 | 9.49 | 0.97 | 3.98 × 10−4 | 0.064 | LOC100153854 |
| ENSSSCG00000044553 | 33.93 | 13.43 | 1.31 | 6.02 × 10−4 | 0.078 | DDIT3 |
| D1 vs. D4 | ||||||
| ENSEMBL ID | D1 | D4 | log2FC | p-Value | padj | Gene Symbol |
| ENSSSCG00000003088 | 42.11 | 90.45 | −1.03 | 2.46 × 10−4 | 0.077 | APOE |
| ENSSSCG00000004052 | 48.08 | 89.79 | −0.94 | 8.00 × 10−5 | 0.052 | FNDC1 |
| ENSSSCG00000005997 | 376.19 | 588.51 | −0.71 | 1.59 × 10−4 | 0.065 | COL14A1 |
| ENSSSCG00000008991 | 147.17 | 270.34 | −1.01 | 3.51 × 10−5 | 0.031 | FRAS1 |
| ENSSSCG00000010554 | 453.56 | 1182.12 | −1.57 | 3.47 × 10−5 | 0.031 | SCD |
| ENSSSCG00000014232 | 19.88 | 39.84 | −1.14 | 2.53 × 10−5 | 0.031 | LOX |
| ENSSSCG00000024149 | 44.81 | 80.94 | −0.98 | 5.22 × 10−6 | 0.021 | ELOVL5 |
| ENSSSCG00000026383 | 41.57 | 63.66 | −0.72 | 3.69 × 10−5 | 0.031 | NRP2 |
| ENSSSCG00000036236 | 81.34 | 160.13 | −1.04 | 2.44 × 10−4 | 0.077 | ELOVL6 |
| ENSSSCG00000038420 | 54.49 | 96.94 | −0.88 | 4.62 × 10−5 | 0.033 | PERP |
| ENSSSCG00000040337 | 49.27 | 97.58 | −0.94 | 9.07 × 10−5 | 0.053 | AK4 |
| ENSSSCG00000032450 | 38.68 | 18.34 | 0.99 | 7.54 × 10−6 | 0.021 | LYRM9 |
| D2 vs. D4 | ||||||
| ENSEMBL ID | D2 | D4 | log2FC | p-value | padj | Gene Symbol |
| ENSSSCG00000016064 | 30.39 | 17.52 | 0.92 | 6.27 × 10−6 | 0.048 | TMEFF2 |
| ENSSSCG00000025053 | 121.60 | 187.28 | −1.08 | 9.06 × 10−6 | 0.048 | RYBP |
| D1 vs. D3 | ||||
| N Genes | Category a | Description | FDR b | Gene symbol |
| 2 | GO MF | protein phosphatase activator activity | 0.0039 | CALM3|CALM1 |
| 6 | GO MF | protein kinase binding | 0.0044 | CD4|PRKAR2B|CALM3|CALM1| ACTB|MARCKS |
| 9 | GO BP | negative regulation of molecular function | 0.0108 | PRKAR2B|IQGAP2|CALM3|CALM1|ACTB|PI16|C4A|CD44|DDIT3 |
| 5 | GO BP | positive regulation of cytosolic calcium ion concentration | 0.0108 | CD4|CCL21|CALM3|CALM1|DDIT3 |
| 2 | GO BP | hyaluronan catabolic process | 0.0112 | LYVE1|CD44 |
| 2 | GO BP | regulation of cellular extravasation | 0.0161 | PLVAP|CCL21 |
| 7 | GO BP | response to biotic stimulus | 0.0176 | CD4|DCLK1|CCL21|CLDN1|C4A| CD44|DDIT3 |
| 6 | GO BP | cell adhesion | 0.0176 | CD4|LYVE1|CCL21|CLDN1|SRPX| CD44 |
| 3 | GO MF | actin filament binding | 0.0177 | IQGAP2|MYH3|MARCKS |
| 4 | GO MF | enzyme inhibitor activity | 0.0191 | PRKAR2B|IQGAP2|PI16|C4A |
| 5 | GO MF | protein domain-specific binding | 0.0226 | PRKAR2B|CALM3|CALM1| SH3BGRL2|DDIT3 |
| 2 | GO BP | release of sequestered calcium ion into cytosol | 0.0255 | CCL21|DDIT3 |
| 14 | GO BP | localization | 0.0336 | CD4|MATN2|DCLK1|CCL21|PRKAR2B|IQGAP2|CALM3|CLDN1|CALM1|ACTB|SRPX|CD44|DDIT3|SLC16A2 |
| 2 | GO BP | multicellular organismal water homeostasis | 0.0348 | PRKAR2B|CLDN1 |
| 3 | GO BP | negative regulation of peptidase activity | 0.0394 | PI16|C4A|CD44 |
| 2 | GO MF | virus receptor activity | 0.0412 | CD4|CLDN1 |
| 2 | GO BP | positive regulation of actin filament polymerization | 0.0432 | CCL21|IQGAP2 |
| 2 | GO BP | response to fatty acid | 0.0436 | CCL21|CLDN1 |
| 5 | GO BP | positive regulation of immune system process | 0.0437 | CD4|PLVAP|CCL21|ACTB|C4A |
| 5 | GO BP | cell motility | 0.0474 | MATN2|DCLK1|CCL21|ACTB|CD44 |
| 2 | GO BP | dendrite development | 0.0474 | MATN2|DCLK1 |
| 7 | GO BP | small molecule metabolic process | 0.0492 | LYVE1|PRKAR2B|CALM3|CALM1| AK4|CD44|MYH3 |
| D1 vs. D4 | ||||
| N Genes | Category | Description | FDR | Gene symbol |
| 3 | Reactome Pathways | Fatty acyl-CoA biosynthesis | 0.0034 | SCD|ELOVL5|ELOVL6 |
| 3 | GO BP | Unsaturated fatty acid biosynthetic process | 0.0146 | SCD|ELOVL5|ELOVL6 |
| 4 | GO BP | Purine ribonucleotide biosynthetic process | 0.0146 | SCD|ELOVL5|ELOVL6|AK4 |
| 2 | GO BP | Fatty acid elongation, saturated fatty acid | 0.0146 | ELOVL5|ELOVL6 |
| 2 | GO BP | Fatty acid elongation, monounsaturated fatty acid | 0.0146 | ELOVL5|ELOVL6 |
| 2 | GO BP | Fatty acid elongation, polyunsaturated fatty acid | 0.0146 | ELOVL5|ELOVL6 |
| 2 | GO BP | Very long-chain fatty acid biosynthetic process | 0.0146 | ELOVL5|ELOVL6 |
| 3 | GO BP | Regulation of cholesterol biosynthetic process | 0.0146 | APOE|SCD|ELOVL6 |
| 4 | GO BP | Regulation of lipid biosynthetic process | 0.0146 | APOE|SCD|ELOVL5|ELOVL6 |
| 3 | GO BP | fatty-acyl-CoA biosynthetic process | 0.0146 | SCD|ELOVL5|ELOVL6 |
| 5 | GO BP | Regulation of small molecule metabolic process | 0.0146 | APOE|SCD|ELOVL5|ELOVL6|AK4 |
| 2 | GO BP | Long-chain fatty-acyl-coa biosynthetic process | 0.0215 | ELOVL5|ELOVL6 |
| 2 | GO MF | Fatty acid elongase activity | 0.0400 | ELOVL5|ELOVL6 |
| 2 | GO MF | 3-oxo-arachidoyl-CoA synthase activity | 0.0400 | ELOVL5|ELOVL6 |
| 2 | GO MF | 3-oxo-cerotoyl-CoA synthase activity | 0.0400 | ELOVL5|ELOVL6 |
| 2 | GO MF | 3-oxo-lignoceronyl-CoA synthase activity | 0.0400 | ELOVL5|ELOVL6 |
| 2 | GO MF | Very-long-chain 3-ketoacyl-coa synthase activity | 0.0400 | ELOVL5|ELOVL6 |
| 2 | KEGG Pathways | Biosynthesis of unsaturated fatty acids | 0.0434 | SCD|ELOVL6 |
| Comparisons | DEGs | Gene Function |
|---|---|---|
| D1 vs. D3 | C4A | C4A (complement C4A) gene favors the reduction of susceptibility to infections as a deficiency of C4A and C4B proteins was associated with an increase in susceptibility to infections [23]. |
| CCL21 | CCL21 (C-C motif chemokine ligand 21) expresses proteins that are part of and promote immune cell migration processes. CCL21 stimulates the migration of T cells and dendritic cells to specific regions of the node in secondary lymphoid organs, where antigen presentation can occur [24]. | |
| LYVE1 | LYVE1 (lymphatic vessel endothelial hyaluronan receptor 1) expresses proteins that are part of and promote immune cell migration processes. LYVE1 expresses a receptor that binds hyaluronic acid present on the membrane of dendritic cells, allowing passage of these cells through lymphatic vessels [25,26]. | |
| PLVAP | PLVAP (plasmalemma vesicle-associated protein) expresses proteins that are part of and promote immune cell migration processes. PLVAP expresses a protein that acts as a physical filter for regulating the entry of lymphocytes and soluble antigens into the parenchyma [27]. | |
| D1 vs. D4 | ELOVL5 | ELOVL5 (ELOVL Fatty Acid Elongase 5) is part of the enzymes group called Elongation of very-long-chain fatty acids (ELOVLs) that catalyze the elongation of two carbon atoms to polyunsaturated fatty acids (PUFAs). ELOVL5 acts in the pathway that leads from alpha-linoleic acid, a polyunsaturated fatty acid of the omega-3 series and found in greater amounts in extruded linseed, to the synthesis of eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) [28]. |
| ELOVL6 | ELOVL6 (ELOVL Fatty Acid Elongase 6) is part of the enzymes group called Elongation of very-long-chain fatty acids (ELOVLs) that catalyzes the elongation of two carbon atoms into saturated and monounsaturated fatty acids [29]. | |
| SCD | SCD (stearoyl-CoA desaturase) is a key enzyme in unsaturated fatty acid biosynthesis, since it catalyzes the insertion of the first double bond into saturated fatty acyl-CoA substrates (palmitoyl-CoA and stearoyl-CoA) at the delta-9 position [30,31]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vegni, J.; Sun, Y.; Seemann, S.E.; Zappaterra, M.; Davoli, R.; Dall’Olio, S.; Gorodkin, J.; Zambonelli, P. RNA-Seq Study on the Longissimus thoracis Muscle of Italian Large White Pigs Fed Extruded Linseed with or without Antioxidants and Polyphenols. Animals 2023, 13, 1187. https://doi.org/10.3390/ani13071187
Vegni J, Sun Y, Seemann SE, Zappaterra M, Davoli R, Dall’Olio S, Gorodkin J, Zambonelli P. RNA-Seq Study on the Longissimus thoracis Muscle of Italian Large White Pigs Fed Extruded Linseed with or without Antioxidants and Polyphenols. Animals. 2023; 13(7):1187. https://doi.org/10.3390/ani13071187
Chicago/Turabian StyleVegni, Jacopo, Ying Sun, Stefan E. Seemann, Martina Zappaterra, Roberta Davoli, Stefania Dall’Olio, Jan Gorodkin, and Paolo Zambonelli. 2023. "RNA-Seq Study on the Longissimus thoracis Muscle of Italian Large White Pigs Fed Extruded Linseed with or without Antioxidants and Polyphenols" Animals 13, no. 7: 1187. https://doi.org/10.3390/ani13071187
APA StyleVegni, J., Sun, Y., Seemann, S. E., Zappaterra, M., Davoli, R., Dall’Olio, S., Gorodkin, J., & Zambonelli, P. (2023). RNA-Seq Study on the Longissimus thoracis Muscle of Italian Large White Pigs Fed Extruded Linseed with or without Antioxidants and Polyphenols. Animals, 13(7), 1187. https://doi.org/10.3390/ani13071187