
Temporal Response of Mesocarnivores to Human Activity and Infrastructure in Taihang Mountains, Central North China: Shifts in Activity Patterns and Their Overlap

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
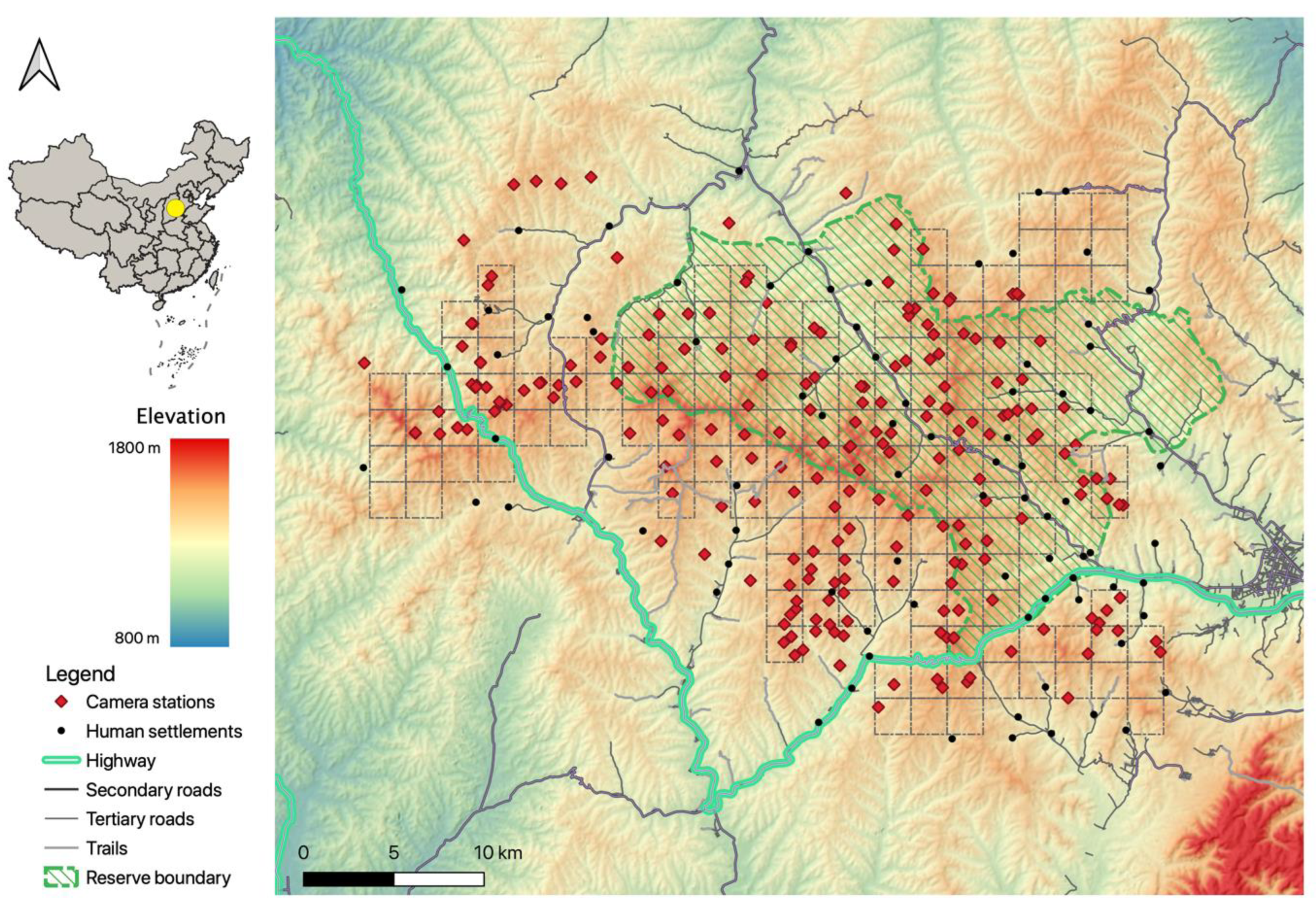
2.1. Study Area
2.2. Data Collection
2.3. Human Disturbance Factors
2.4. Data Analysis
2.4.1. Activity Overlaps under Large-Scale Human Disturbance
2.4.2. Activity Overlaps under Fine-Scale Human Disturbance
3. Results
3.1. Large-Scale Niche Partitioning between Mesocarnivore Pairs
3.2. Fine-Scale Niche Partitioning between Mesocarnivore Pairs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Watson, J.E.; Shanahan, D.F.; Di Marco, M.; Allan, J.; Laurance, W.F.; Sanderson, E.W.; Mackey, B.; Venter, O. Catastrophic declines in wilderness areas undermine global environment targets. Curr. Biol. 2016, 26, 2929–2934. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.; Lindenmayer, D.B. Landscape modification and habitat fragmentation: A synthesis. Glob. Ecol. Biogeogr. 2007, 16, 265–280. [Google Scholar] [CrossRef]
- Smith, J.A.; Thomas, A.C.; Levi, T.; Wang, Y.; Wilmers, C.C. Human activity reduces niche partitioning among three widespread mesocarnivores. Oikos 2018, 127, 890–901. [Google Scholar] [CrossRef]
- Ellis, E.C. Anthropogenic transformation of the terrestrial biosphere. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2011, 369, 1010–1035. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.J.; Knight, R.L.; Marzluff, J.; Powell, S.; Brown, K.; Gude, P.H.; Jones, K. Effects of exurban development on biodiversity: Patterns, mechanisms, and research needs. Ecol. Appl. 2005, 15, 1893–1905. [Google Scholar] [CrossRef]
- Llaneza, L.; López-Bao, J.V.; Sazatornil, V. Insights into wolf presence in human-dominated landscapes: The relative role of food availability, humans and landscape attributes. Divers. Distrib. 2011, 18, 459–469. [Google Scholar] [CrossRef]
- Oriol-Cotterill, A.; Valeix, M.; Frank, L.G.; Riginos, C.; Macdonald, D.W. Landscapes of coexistence for terrestrial carnivores: The ecological consequences of being downgraded from ultimate to penultimate predator by humans. Oikos 2015, 124, 1263–1273. [Google Scholar] [CrossRef]
- Sévêque, A.; Gentle, L.K.; López-Bao, J.V.; Yarnell, R.W.; Uzal, A. Impact of human disturbance on temporal partitioning within carnivore communities. Mammal Rev. 2021, 52, 67–81. [Google Scholar] [CrossRef]
- Moll, R.J.; Cepek, J.D.; Lorch, P.D.; Dennis, P.M.; Robison, T.; Millspaugh, J.J.; Montgomery, R.A. Humans and urban development mediate the sympatry of competing carnivores. Urban Ecosyst. 2018, 21, 765–778. [Google Scholar] [CrossRef]
- Gaynor, K.M.; Brown, J.S.; Middleton, A.D.; Power, M.E.; Brashares, J.S. Landscapes of fear: Spatial patterns of risk perception and response. Trends Ecol. Evol. 2019, 34, 355–368. [Google Scholar] [CrossRef]
- Alberti, M.; Marzluff, J.M.; Shulenberger, E.; Bradley, G.; Ryan, C.; Zumbrunnen, C. Integrating humans into ecology: Opportunities and challenges for studying urban ecosystems. BioScience 2003, 53, 1169–1179. [Google Scholar] [CrossRef]
- Dorresteijn, I.; Schultner, J.; Nimmo, D.G.; Fischer, J.; Hanspach, J.; Kuemmerle, T.; Kehoe, L.; Ritchie, E. Incorporating anthropogenic effects into trophic ecology: Predator–prey interactions in a human-dominated landscape. Proc. R. Soc. B Boil. Sci. 2015, 282, 20151602. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, R.; Levins, R. The limiting similarity, convergence, and divergence of coexisting species. Am. Nat. 1967, 101, 377–385. [Google Scholar] [CrossRef]
- Schoener, T.W. Resource partitioning in ecological communities. Science 1974, 185, 27–39. [Google Scholar] [CrossRef]
- Apps, C.D.; McLellan, B.N.; Woods, J.G. Landscape partitioning and spatial inferences of competition between black and grizzly bears. Ecography 2006, 29, 561–572. [Google Scholar] [CrossRef]
- Easter, T.; Bouley, P.; Carter, N. Intraguild dynamics of understudied carnivores in a human-altered landscape. Ecol. Evol. 2020, 10, 5476–5488. [Google Scholar] [CrossRef] [PubMed]
- Palei, H.S.; Sahu, H.K.; Nayak, A.K. Competition versus opportunism: Diet and trophic niche relationship of two sympatric apex carnivores in a tropical forest. Acta Ecol. Sin. 2021, 43, 99–105. [Google Scholar] [CrossRef]
- Frey, S.; Volpe, J.P.; Heim, N.A.; Paczkowski, J.; Fisher, J.T. Move to nocturnality not a universal trend in carnivore species on disturbed landscapes. Oikos 2020, 129, 1128–1140. [Google Scholar] [CrossRef]
- Kronfeld-Schor, N.; Dayan, T. Partitioning of time as an ecological resource. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 153–181. [Google Scholar] [CrossRef]
- Fedriani, J.M.; Fuller, T.K.; Sauvajot, R.M.; York, E.C. Competition and intraguild predation among three sympatric carnivores. Oecologia 2000, 125, 258–270. [Google Scholar] [CrossRef]
- Karanth, K.U.; Srivathsa, A.; Vasudev, D.; Puri, M.; Parameshwaran, R.; Kumar, N.S. Spatio-temporal interactions facilitate large carnivore sympatry across a resource gradient. Proc. R. Soc. B Boil. Sci. 2017, 284, 20161860. [Google Scholar] [CrossRef] [PubMed]
- Tsunoda, H.; Newman, C.; Peeva, S.; Raichev, E.; Buesching, C.D.; Kaneko, Y. Spatio-temporal partitioning facilitates mesocarnivore sympatry in the Stara Planina Mountains, Bulgaria. Zoology 2020, 141, 125801. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Allen, M.L.; Wilmers, C.C. Mesopredator spatial and temporal responses to large predators and human development in the Santa Cruz Mountains of California. Biol. Conserv. 2015, 190, 23–33. [Google Scholar] [CrossRef]
- Manlick, P.J.; Pauli, J.N. Human disturbance increases trophic niche overlap in terrestrial carnivore communities. Proc. Natl. Acad. Sci. USA 2020, 117, 26842–26848. [Google Scholar] [CrossRef] [PubMed]
- Polis, G.A.; Holt, R.D. Intraguild predation: The dynamics of complex trophic interactions. Trends Ecol. Evol. 1992, 7, 151–154. [Google Scholar] [CrossRef]
- Gaynor, K.M.; Hojnowski, C.E.; Carter, N.H.; Brashares, J.S. The influence of human disturbance on wildlife nocturnality. Science 2018, 360, 1232–1235. [Google Scholar] [CrossRef] [PubMed]
- Sévêque, A.; Gentle, L.K.; López-Bao, J.V.; Yarnell, R.W.; Uzal, A. Human disturbance has contrasting effects on niche partitioning within carnivore communities. Biol. Rev. 2020, 95, 1689–1705. [Google Scholar] [CrossRef]
- Roemer, G.W.; Gompper, M.E.; Van Valkenburgh, B. The ecological role of the mammalian mesocarnivore. Bioscience 2009, 59, 165–173. [Google Scholar] [CrossRef]
- Suraci, J.P.; Clinchy, M.; Dill, L.M.; Roberts, D.; Zanette, L.Y. Fear of large carnivores causes a trophic cascade. Nat. Commun. 2016, 7, 10698. [Google Scholar] [CrossRef]
- Schuette, P.; Wagner, A.P.; Wagner, M.E.; Creel, S. Occupancy patterns and niche partitioning within a diverse carnivore community exposed to anthropogenic pressures. Biol. Conserv. 2013, 158, 301–312. [Google Scholar] [CrossRef]
- Bu, H.; Wang, F.; McShea, W.J.; Lu, Z.; Wang, D.; Li, S. Spatial co-occurrence and activity patterns of mesocarnivores in the temperate forests of Southwest China. PLoS ONE 2016, 11, e0164271. [Google Scholar] [CrossRef] [PubMed]
- Suraci, J.P.; Clinchy, M.; Zanette, L.Y.; Wilmers, C.C. Fear of humans as apex predators has landscape-scale impacts from mountain lions to mice. Ecol. Lett. 2019, 22, 1578–1586. [Google Scholar] [CrossRef] [PubMed]
- Vitekere, K.; Wang, J.; Karanja, H.; Consolée, K.T.; Jiang, G.; Hua, Y. Dynamic in species estimates of carnivores (leopard cat, red fox, and north Chinese leopard): A multi-year assessment of occupancy and coexistence in the Tieqiaoshan Nature Reserve, Shanxi Province, China. Animals 2020, 10, 1333. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Vitekere, K.; Wang, J.; Zhu, M.; Zaman, M.; Jiang, G. Coexistence of sympatric carnivores in a relatively homogenous landscape and the effects of environmental factors on site occupation. Ann. Zoöl. Fenn. 2020, 57, 47. [Google Scholar] [CrossRef]
- Shao, X.; Song, D.; Huang, Q.; Li, S.; Yao, M. Fast surveys and molecular diet analysis of carnivores based on fecal DNA and metabarcoding. Biodivers. Sci. 2019, 27, 543–556. [Google Scholar]
- Zhao, G.; Yang, H.; Xie, B.; Gong, Y.; Ge, J.; Feng, L. Spatio-temporal coexistence of sympatric mesocarnivores with a single apex carnivore in a fine-scale landscape. Glob. Ecol. Conserv. 2020, 21, e00897. [Google Scholar] [CrossRef]
- Zheng, J.G.; Zhang, Y.F.; Wang, Y.; Dong, D.P. The characteristics of plant distribution and diversity in the middle section of Taihang Mountain. Henan Sci. 2009, 27, 292–294. [Google Scholar]
- Shi, L.; Feng, Y.; Gao, L. The method of territorial spatial development suitability evaluation in the Yangtze River Delta: A case study of Changxing County. ACTA Ecol. Sin. 2020, 40, 6495–6504. [Google Scholar]
- Zhao, D. Development direction of Tieqiaoshan Nature Reserve, Shanxi. Shanxi For. 2018, 1, 26–27. [Google Scholar] [CrossRef]
- Liu, Y.; Song, D.; Liu, B.; Xia, F.; Chen, Y.; Wang, Y.; Huang, Q. Overview of the Camera-trapping Platform for Felid Species in China: Data integration by a conservation NGO. Biodivers. Sci. 2020, 28, 1067–1074. [Google Scholar] [CrossRef]
- Vitekere, K. North China Leopard Conservation Status and Coexistence Patterns with Red Fox and Leopard Cat in Tieqiaoshan Nature Reserve, Shanxi. Ph.D. Thesis, Northeast Forestry University, Harbin, China, 2021. [Google Scholar]
- Weng, Y.; McShea, W.; Diao, Y.; Yang, H.; Zhang, X.; Gu, B.; Bu, H.; Wang, F. The incursion of free-ranging dogs into protected areas: A spatio-temporal analysis in a network of giant panda reserves. Biol. Conserv. 2021, 265, 109423. [Google Scholar] [CrossRef]
- Gompper, M.E.; Kays Roland, W.; Ray Justin, A.C.; Scott, D.; Bogan Daniel, A.; Cryan Jason, R. A comparison of noninvasive techniques to survey carnivore communities in northeastern North America. Wildl. Soc. Bull. 2006, 34, 1142–1151. [Google Scholar] [CrossRef]
- O’Brien, T.G.; Kinnaird, M.F.; Wibisono, H.T. Crouching tigers, hidden prey: Sumatran tiger and prey populations in a tropical forest landscape. Anim. Conserv. 2003, 6, 131–139. [Google Scholar] [CrossRef]
- Brook, L.A.; Johnson, C.N.; Ritchie, E.G. Effects of predator control on behaviour of an apex predator and indirect consequences for mesopredator suppression. J. Appl. Ecol. 2012, 49, 1278–1286. [Google Scholar] [CrossRef]
- Palmer, M.S.; Swanson, A.; Kosmala, M.; Arnold, T.; Packer, C. Evaluating relative abundance indices for terrestrial herbivores from large-scale camera trap surveys. Afr. J. Ecol. 2018, 56, 791–803. [Google Scholar] [CrossRef]
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation Project. 2022. Available online: http://qgis.osgeo.org (accessed on 28 March 2022).
- Van der Weyde, L.K.; Mbisana, C.; Klein, R. Multi-species occupancy modelling of a carnivore guild in wildlife management areas in the Kalahari. Biol. Conserv. 2018, 220, 21–28. [Google Scholar] [CrossRef]
- Hartigan, J.A.; Wong, M.A. Algorithm AS 136: A K-means clustering algorithm. J. R. Stat. Soc. Ser. Appl. Stat. 1979, 28, 100–108. [Google Scholar] [CrossRef]
- Kabacoff, R. Cluster Analysis. In R in Action; essay; Manning Publications: Shelter Island, NY, USA, 2022; pp. 369–388. [Google Scholar]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R package version 1.0.7. 2020. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 7 November 2021).
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K. Cluster: Cluster Analysis Basics and Extensions. R package version 1.14.4.K. 2013. Available online: https://CRAN.R-project.org/web/packages/cluster/index.html (accessed on 7 November 2021).
- Charrad, M.; Ghazzali, N.; Boiteau, V.; Niknafs, A. NbClust: An R package for determining the relevant number of clusters in a data set. J. Stat. Softw. 2014, 61, 1–36. [Google Scholar] [CrossRef]
- R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; R Core Development Team: Vienna, Austria; Available online: https://www.R-project.org/ (accessed on 5 July 2021).
- Ridout, M.S.; Linkie, M. Estimating overlap of daily activity patterns from camera trap data. J. Agric. Biol. Environ. Stat. 2009, 14, 322–337. [Google Scholar] [CrossRef]
- Meredith, M.; Ridout Martin, S. The Overlap Package. R package version 0.3.4. 2014. Available online: https://CRAN.R-project.org/web/packages/overlap/ (accessed on 10 July 2021).
- Zimmerman, D.W.; Zumbo, B.D. Rank transformations and the power of the Student t test and Welch t’ test for non-normal populations with unequal variances. Can. J. Exp. Psychol./Rev. Can. De Psychol. Expérimentale 1993, 47, 523–539. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D. nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-161. 2022. Available online: https://CRAN.R-project.org/web/packages/nlme/ (accessed on 3 January 2023).
- Johnson, J.B.; Omland, K.S. Model selection in ecology and evolution. Trends Ecol. Evol. 2004, 19, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Zuur, A.F.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R, 1st ed.; Springer: New York, NY, USA, 2009; pp. 209–259. [Google Scholar]
- Li, T.; Meng, D.; Liwei, T.; Si, Y.; Zhang, Z.; Liu, Z. Activity rhythm of red fox in Luoshan National Nature Reserve based on infrared camera technology. Chin. J. Wildl. 2020, 41, 891–896. [Google Scholar]
- Zhou, W.; Wei, W. Activity rhythms and distribution of natal dens for red foxes. Acta Theriol. Sin. 1995, 15, 267–272. [Google Scholar]
- Hu, Q.; Lin, H.; Dai, Q.; Yang, Z.; He, L.; Zhang, W.; Shi, X. Niche differentiation among three middle-sized carnivores in Wolong Nature Reserve. Chin. J. Zool. 2020, 55, 685–691. [Google Scholar] [CrossRef]
- Zagainova, O.S.; Markov, N. The diet of Asian badger, Meles leucurus Hodgson, 1847, in Samarovskii Chugas Nature Park, Western Siberia. Russ. J. Ecol. 2011, 42, 414–420. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, W.; Kaneko, Y.; Newman, C.; Liao, Z.; Zhu, X.; Buesching, C.D.; Xie, Z.; Macdonald, D.W. Seasonal dietary shifts and food resource exploitation by the hog badger (Arctonyx collaris) in a Chinese subtropical forest. Eur. J. Wildl. Res. 2014, 61, 125–133. [Google Scholar] [CrossRef]
- Nickel, B.A.; Suraci, J.P.; Allen, M.L.; Wilmers, C.C. Human presence and human footprint have non-equivalent effects on wildlife spatiotemporal habitat use. Biol. Conserv. 2019, 241, 108383. [Google Scholar] [CrossRef]
- Wu, J.; Wang, J.; Zhu, Y.; Bu, X.; Xiang, R.; Lu, Q.; Cui, S.; Hao, Y.; Sheng, Y.; Meng, X. Summer habitat selection and impacts of human disturbance on leopard cats (Prionailurus bengalensis). Ecosyst. Health Sustain. 2020, 6, 1856630. [Google Scholar] [CrossRef]
- Berger, J. Fear, human shields and the redistribution of prey and predators in protected areas. Biol. Lett. 2007, 3, 620–623. [Google Scholar] [CrossRef]
- Lee, O.; Lee, S.; Nam, D.-H.; Lee, H.Y. Food habits of the leopard cat (Prionailurus bengalensis euptilurus) in Korea. Mammal Study 2014, 39, 43–46. [Google Scholar] [CrossRef]
- Wang, J.; Ji, S.; Wu, J.; Shrestha, T.K.; Bu, X.; Zhu, Y.; Xiang, R.; Sheng, Y.; Meng, X. Away from the city: Habitat selection of badgers in mountainous area around Beijing. Biologia 2021, 76, 1737–1746. [Google Scholar] [CrossRef]
- Bae, H.-K.; Lee, J.-K.; Eom, T.-K.; Lee, D.-H.; Rhim, S.-J. Ecological factors influencing the selection of sett location by the Asian badger Meles leucurus. Wildl. Biol. 2021, 2021, wlb.00910. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, Y.-B.; Newman, C.; Kaneko, Y.; Macdonald, D.; Jiang, P.-P.; Ding, P. Niche overlap and sett-site resource partitioning for two sympatric species of badger. Ethol. Ecol. Evol. 2009, 21, 89–100. [Google Scholar] [CrossRef]
- Bao, W.; Li, X.; Shi, Y. Comparative analysis of food habits in carnivores from three areas of Beijing. Zool. Res. 2005, 26, 118–122. [Google Scholar]
- Zhang, L.; Wang, A.; Li, Y.; Bao, W.; Yang, Y.; Bater. Preliminary comparison of diet composition of four small sized carnivores at Saihanwula Nature Reserve, Inner Mongolia. Acta Theriol. Sin. 2011, 31, 55–61. [Google Scholar] [CrossRef]
- Díaz-Ruiz, F.; Delibes-Mateos, M.; García-Moreno, J.L.; López-Martín, J.M.; Ferreira, C.; Ferreras, P. Biogeographical patterns in the diet of an opportunistic predator: The red fox Vulpes vulpes in the Iberian Peninsula. Mammal Rev. 2011, 43, 59–70. [Google Scholar] [CrossRef]
- Castañeda, I.; Doherty, T.S.; Fleming, P.A.; Stobo-Wilson, A.M.; Woinarski, J.C.Z.; Newsome, T.M. Variation in red fox Vulpes vulpes diet in five continents. Mammal Rev. 2022, 52, 328–342. [Google Scholar] [CrossRef]
- Soe, E.; Davison, J.; Süld, K.; Valdmann, H.; Laurimaa, L.; Saarma, U. Europe-wide biogeographical patterns in the diet of an ecologically and epidemiologically important mesopredator, the red fox Vulpes vulpes: A quantitative review. Mammal Rev. 2017, 47, 198–211. [Google Scholar] [CrossRef]
- Seryodkin, I.V.; Burkovskiy, O.A. Food habit analysis of the Amur leopard cat Prionailurus bengalensis euptilurus in the Russian Far East. Biol. Bull. 2019, 46, 648–653. [Google Scholar] [CrossRef]
- Li, Z.; Tang, Z.; Xu, Y.; Wang, Y.; Duan, Z.; Liu, X.; Wang, P.; Yang, J.; Chen, W.; Prins, H.H.T. Habitat use and activity patterns of mammals and birds in relation to temperature and vegetation cover in the alpine ecosystem of Southwestern China with camera-trapping monitoring. Animals 2021, 11, 3377. [Google Scholar] [CrossRef]
- Gause, G.F. The Struggle for Existence; Williams and Wilkins: Baltimore, MD, USA, 1934. [Google Scholar]
- Arjo, W.M.; Pletscher, D.H. Behavioral responses of coyotes to wolf recolonization in northwestern Montana. Can. J. Zoöl. 1999, 77, 1919–1927. [Google Scholar] [CrossRef]
- Iwama, M.; Yamazaki, K.; Matsuyama, M.; Hoshino, Y.; Hisano, M.; Newman, C.; Kaneko, Y. Masked palm civet Paguma larvata summer diet differs between sexes in a suburban area of central Japan. Mammal Study 2017, 42, 185–190. [Google Scholar] [CrossRef]
- Wang, H.; Fuller, T. Food habits of four sympatric carnivores in southeastern China. Mammalia 2003, 67, 513–520. [Google Scholar] [CrossRef]
- Gasparatos, A.; Doll, C.N.; Esteban, M.; Ahmed, A.; Olang, T.A. Renewable energy and biodiversity: Implications for transitioning to a Green Economy. Renew. Sustain. Energy Rev. 2017, 70, 161–184. [Google Scholar] [CrossRef]
- Ayram, C.A.C.; Mendoza, M.E.; Etter, A.; Salicrup, D.R.P. Anthropogenic impact on habitat connectivity: A multidimensional human footprint index evaluated in a highly biodiverse landscape of Mexico. Ecol. Indic. 2017, 72, 895–909. [Google Scholar] [CrossRef]
- Frey, S.; Fisher, J.T.; Burton, A.C.; Volpe, J.P. Investigating animal activity patterns and temporal niche partitioning using camera-trap data: Challenges and opportunities. Remote. Sens. Ecol. Conserv. 2017, 3, 123–132. [Google Scholar] [CrossRef]
- Moore, H.A.; Valentine, L.E.; Dunlop, J.A.; Nimmo, D.G. The effect of camera orientation on the detectability of wildlife: A case study from north-western Australia. Remote. Sens. Ecol. Conserv. 2020, 6, 546–556. [Google Scholar] [CrossRef]
- Tourani, M.; Brøste, E.N.; Bakken, S.; Odden, J.; Bischof, R. Sooner, closer, or longer: Detectability of mesocarnivores at camera traps. J. Zoöl. 2020, 312, 259–270. [Google Scholar] [CrossRef]
- Archaux, F.; Henry, P.-Y.; Gimenez, O. When can we ignore the problem of imperfect detection in comparative studies? Methods Ecol. Evol. 2011, 3, 188–194. [Google Scholar] [CrossRef]
- Ferreras, P.; Ruiz, F.D.; Monterroso, P. Improving mesocarnivore detectability with lures in camera-trapping studies. Wildl. Res. 2018, 45, 505. [Google Scholar] [CrossRef]
- Cardillo, M.; Mace, G.M.; Jones, K.E.; Bielby, J.; Bininda-Emonds, O.R.P.; Sechrest, W.; Orme, C.D.L.; Purvis, A. Multiple causes of high extinction risk in large mammal species. Science 2005, 309, 1239–1241. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Pair | Difference | p-Value | ||
|---|---|---|---|---|
| Badger–human | 0.41 (0.39, 0.43) | 0.16 (0.05, 0.31) | −0.25 | 0.000 |
| Leopard cat–human | 0.27 (0.25, 0.29) | 0.11 (0.03, 0.21) | −0.16 | 0.000 |
| Red fox–human | 0.27 (0.25, 0.29) | 0.27(0.13, 0.43) | 0.00 | 0.730 |
| Leopard cat–badger | 0.86 (0.83, 0.88) | 0.75(0.50, 0.94) | −0.11 | 0.324 |
| Red fox–leopard cat | 0.89 (0.87, 0.91) | 0.80(0.59, 0.97) | −0.09 | 0.365 |
| Red fox–badger | 0.84 (0.82, 0.86) | 0.76(0.53, 0.96) | −0.08 | 0.471 |
| Species Pairs | ||||||
|---|---|---|---|---|---|---|
| Leopard Cat, Red Fox | Leopard Cat, Badger | Red Fox, Badger | ||||
| Random Effects Village ID intercept Road ID intercept | ||||||
| 0.000 (0.000) | ||||||
| 0.003 (0.055) | ||||||
| Residual Fixed Effects | 0.033 (0.182) | |||||
| (SE) | t value | (SE) | t value | (SE) | t value | |
| (Intercept) Road distance Village distance Human activity | 0.606 (0.016) | 36.946 | 0.609 (0.018) | 33.773 | 0.606 (0.016) | 36.946 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Liu, B.; Fan, D.; Li, S. Temporal Response of Mesocarnivores to Human Activity and Infrastructure in Taihang Mountains, Central North China: Shifts in Activity Patterns and Their Overlap. Animals 2023, 13, 688. https://doi.org/10.3390/ani13040688
Chen Y, Liu B, Fan D, Li S. Temporal Response of Mesocarnivores to Human Activity and Infrastructure in Taihang Mountains, Central North China: Shifts in Activity Patterns and Their Overlap. Animals. 2023; 13(4):688. https://doi.org/10.3390/ani13040688
Chicago/Turabian StyleChen, Yanzhi, Beibei Liu, Deqing Fan, and Sheng Li. 2023. "Temporal Response of Mesocarnivores to Human Activity and Infrastructure in Taihang Mountains, Central North China: Shifts in Activity Patterns and Their Overlap" Animals 13, no. 4: 688. https://doi.org/10.3390/ani13040688
APA StyleChen, Y., Liu, B., Fan, D., & Li, S. (2023). Temporal Response of Mesocarnivores to Human Activity and Infrastructure in Taihang Mountains, Central North China: Shifts in Activity Patterns and Their Overlap. Animals, 13(4), 688. https://doi.org/10.3390/ani13040688