
New Behavioral Handling Test Reveals Temperament Differences in Native Japanese Chickens
 ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Note
2.2. Animals
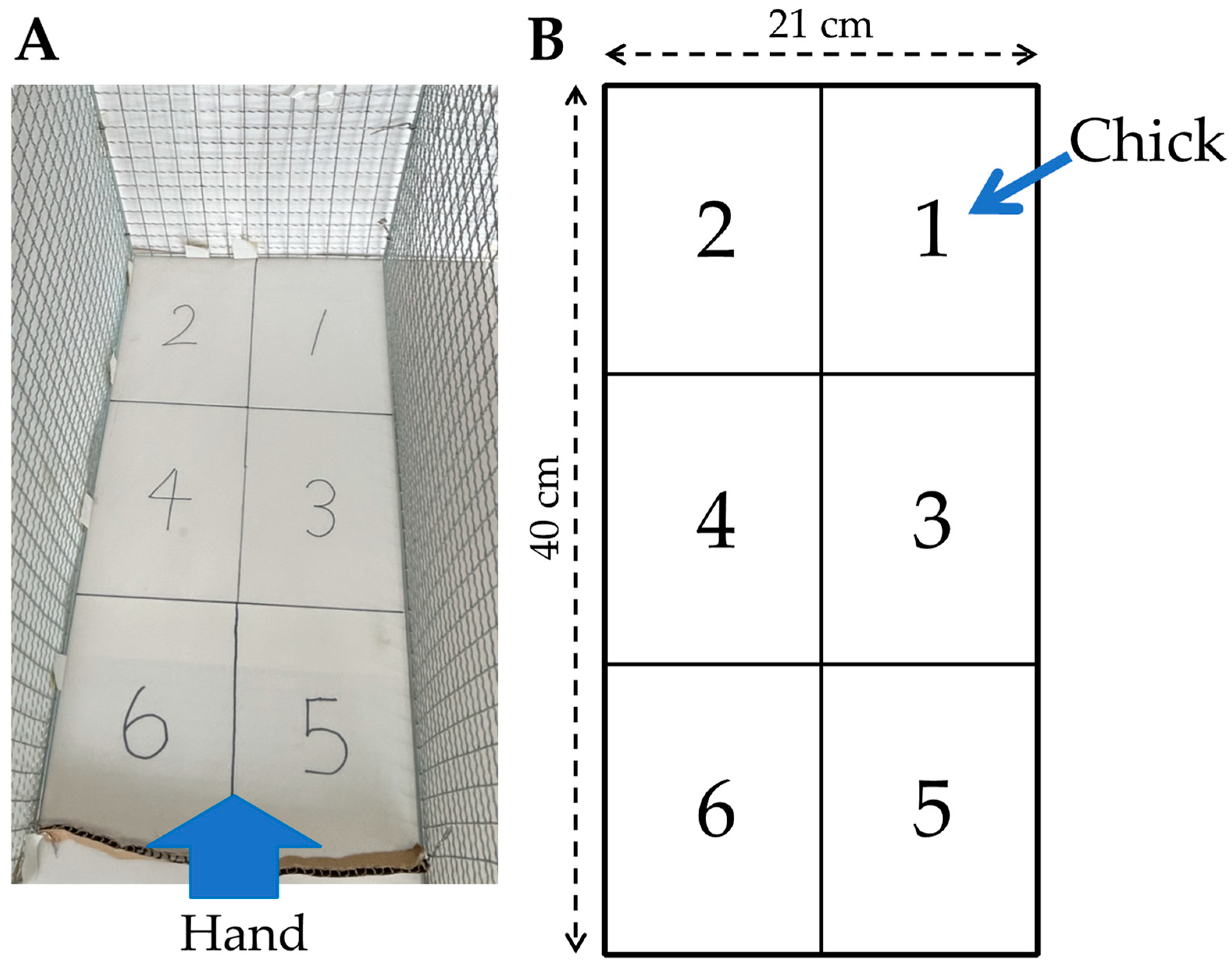
2.3. Handling Test
2.4. Sexing
2.5. Statistical Analysis
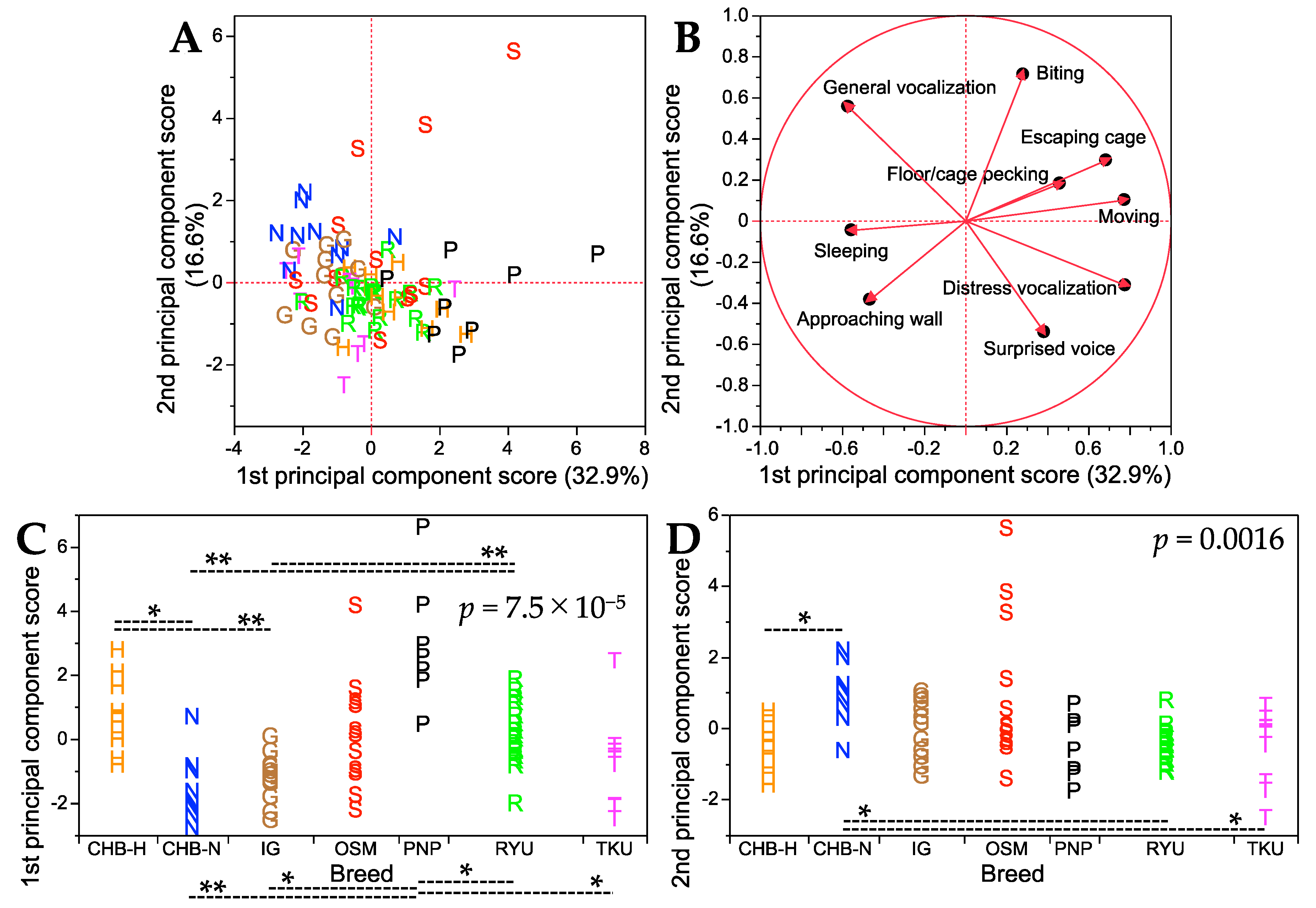
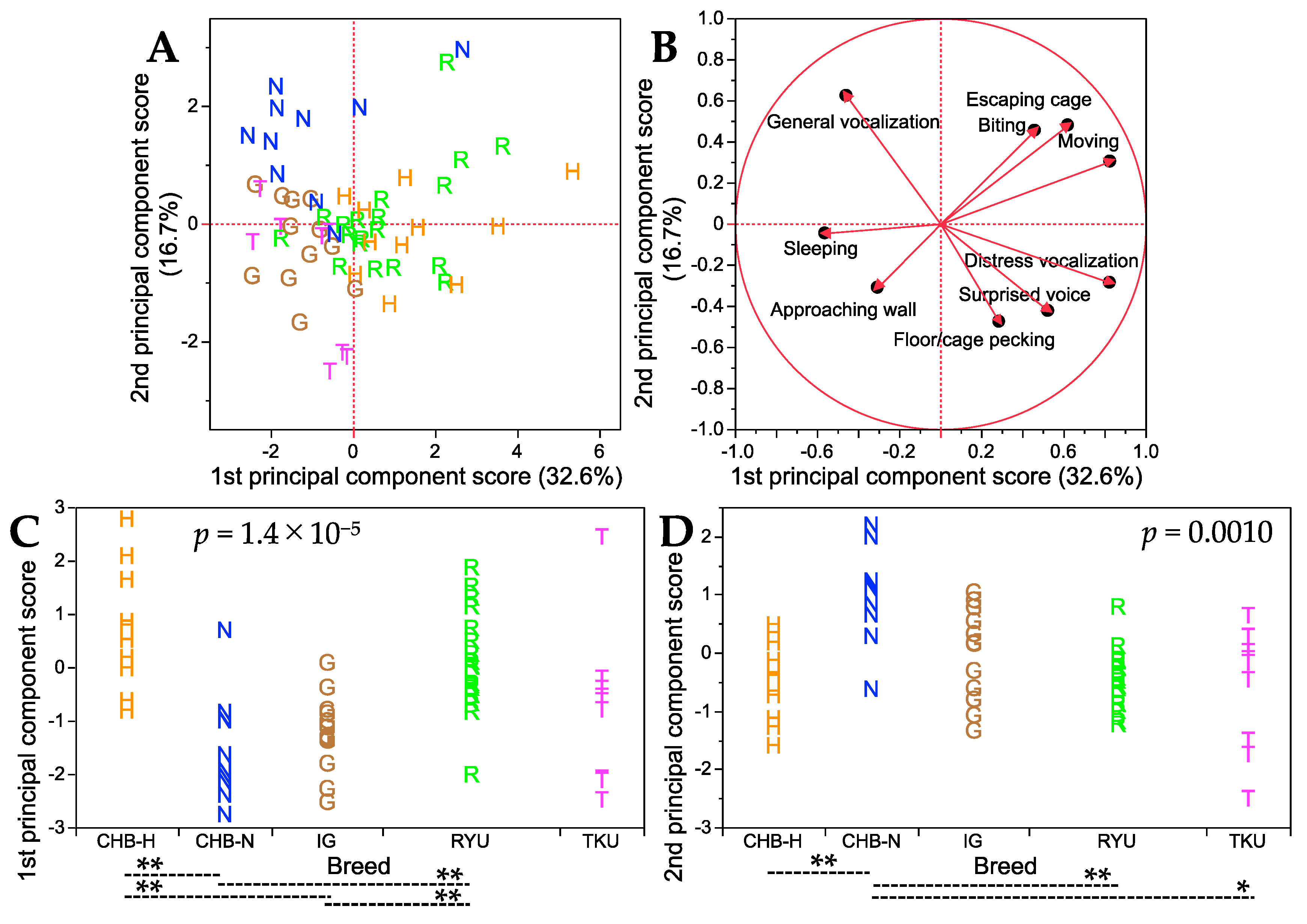
3. Results
Handling Test
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lupien, S.J.; McEwen, B.S.; Gunner, M.R.; Heim, C. Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat. Rev. Neurosci. 2009, 10, 434–445. [Google Scholar] [CrossRef]
- Hedlund, L.; Whittle, R.; Jensen, P. Effects of commercial hatchery processing on short-and long-term stress responses in laying hens. Sci. Rep. 2019, 9, 2367. [Google Scholar] [CrossRef]
- Hedlund, L.; Jensen, P. Incubation and hatching conditions of laying hen chicks explain a large part of the stress effects from commercial large-scale hatcheries. Poult. Sci. 2021, 100, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Grams, V.; Bögelein, S.; Grashorn, M.A.; Bessei, W.; Bennewitz, J. Quantitative genetic analysis of traits related to fear and feather pecking in laying hens. Behav. Genet. 2015, 45, 228–235. [Google Scholar] [CrossRef]
- Tsudzuki, M. Japanese native chickens. In The Relationships between Indigenous Animals and Humans in the APEC Region; Chang, H.-L., Huang., Y.-C., Eds.; The Chinese Society of Animal Science: Taipei, China, 2003; pp. 91–116. [Google Scholar]
- Kato, S.; Ohguchi, H.; Ando, I.; Kawamura, T.; Ohsuga, A. Studies on feeding and management for Nagoya breed: Effects of bird density and debeaking treatment on productivity of birds, and factors causing crowding accidents when rearing birds in a big flock on floor pens. Res. Bull. Aichi-Ken Agric. Res. Cent. 1991, 23, 453–457. (In Japanese) [Google Scholar]
- Forkman, B.; Boissy, A.; Meunier-Salaün, M.C.; Canali, E.; Jones, R.B. A critical review of fear tests used on cattle, pigs, sheep, poultry and horses. Physiol. Behav. 2007, 92, 340–374. [Google Scholar] [CrossRef]
- Gallup, G.G., Jr.; Ledbetter, D.H.; Maser, J.D. Strain differences among chickens in tonic immobility: Evidence for an emotionality component. J. Comp. Physiol. Psychol. 1976, 90, 1075–1081. [Google Scholar] [CrossRef]
- Jones, R.B.; Faure, J.M. Sex and strain comparisons of tonic immobility (“righting time”) in the domestic fowl and the effects of various methods of induction. Behav. Process. 1981, 6, 47–55. [Google Scholar] [CrossRef]
- Hocking, P.M.; Channing, C.E.; Waddington, D.; Jones, R.B. Age-related changes in fear, sociality and pecking behaviours in two strains of laying hen. Br. Poult. Sci. 2001, 42, 414–423. [Google Scholar] [CrossRef]
- Faure, J.M.; Jones, R.B.; Bessei, W. Fear and social motivation as factors in open-field behaviour of the domestic chick. Behav. Biol. 1983, 8, 103–116. [Google Scholar]
- Jones, R.B. Fear responses of individually caged laying hens as a function of cage level and aisle. Appl. Anim. Behav. Sci. 1985, 14, 63–74. [Google Scholar] [CrossRef]
- Barnett, J.L.; Hemsworth, P.H. Fear of humans by laying hens in different tiers of a battery: Behavioural and physiological responses. Br. Poult. Sci. 1989, 30, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Moroi, S.; Nishimura, K.; Imai, N.; Kunishige, K.; Sato, S.; Goto, T. Rapid behavioral assay using handling test provides breed and sex differences in tameness of chickens. Brain Behav. 2019, 9, e01394. [Google Scholar] [CrossRef]
- Sakaguchi, M.; Ishikawa, A. Tonic immobility and open field responses in Nagoya, White Leghorn, and White Plymouth rock chicks. J. Poult. Sci. 2020, 57, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Terashima, M.; Velasco, V.V.; Goto, N.; Tsudzuki, M.; Ishikawa, A. Differences in innate fear behaviour in native Japanese chickens. Br. Poult. Sci. 2023, 64, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Yoshidome, K.; Fukano, N.; Ouchi, Y.; Tomonaga, S.; Cockrem, J.F.; Bungo, T. The use of behavioral tests of fearfulness in chicks to distinguish between the Japanese native chicken breeds, Tosa-Kukin and Yakido. Anim. Sci. J. 2021, 92, e13507. [Google Scholar] [CrossRef] [PubMed]
- Nunome, M.; Kinoshita, K.; Ishishita, S.; Ohmori, Y.; Murai, A.; Matsuda, Y. Genetic diversity of 21 experimental chicken lines with diverse origins and genetic backgrounds. Exp. Anim. 2019, 68, 177–193. [Google Scholar] [CrossRef] [PubMed]
- Hashiguchi, T.; Fujisaki, T.; Maeda, Y.; Taketomi, M. Genetics of the hypoplasia of feathers of the tail region in the Ingie fowl. Br. Poult. Sci. 1978, 19, 487–490. [Google Scholar] [CrossRef]
- Oka, T.; Fujihara, M.; Fukushima, M.; Bungo, T.; Tsudzuki, M. Genetic diversity and structure of the Ryujin-Jidori, a critically endangered native Japanese chicken breed, based on microsatellite DNA analysis. J. Poult. Sci. 2014, 51, 387–391. [Google Scholar] [CrossRef]
- Herborn, K.A.; McElligott, A.G.; Mitchell, M.A.; Sandilands, V.; Bradshaw, B.; Asher, L. Spectral entropy of early-life distress calls as an iceberg indicator of chicken welfare. J. R. Soc. Interface 2020, 17, 20200086. [Google Scholar] [CrossRef]
- Suzuki, S.; Kobayashi, M.; Murai, A.; Tsudzuki, M.; Ishikawa, A. Characterization of growth, fat deposition, and lipid me- tabolism-related gene expression in lean and obese meat-type chickens. J. Poult. Sci. 2019, 56, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Hata, A.; Takenouchi, A.; Kinoshita, K.; Hirokawa, M.; Igawa, T.; Nunome, M.; Suzuki, T.; Tsudzuki, M. Geographic origin and genetic characteristics of Japanese indigenous chickens inferred from mitochondrial D-loop region and microsatellite DNA markers. Animals 2020, 10, 2074. [Google Scholar] [CrossRef] [PubMed]
- Osman, S.A.M.; Sekino, M.; Nishihata, A.; Kobayashi, Y.; Takenaka, W.; Kinoshita, K.; Kuwayama, T.; Nishibori, M.; Yamamoto, Y.; Tsudzuki, M. The genetic variability and relationships of Japanese and foreign chickens assessed by microsatellite DNA Profiling. Asian-Australas. J. Ani. Sci. 2006, 19, 1369–1378. [Google Scholar] [CrossRef]
- Hu, Z.-L.; Park, C.A.; Reecy, J.M. Bringing the Animal QTLdb and CorrDB into the future: Meeting new challenges and providing updated services. Nucleic Acids Res. 2022, 50, D956–D961. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, A.; Sakaguchi, M.; Nagano, A.J.; Suzuki, S. Genetic architecture of innate fear behavior in chickens. Behav. Genet. 2020, 50, 411–422. [Google Scholar] [CrossRef]
- Ochiai, T.; Sakaguchi, M.; Kawakami, S.-I.; Ishikawa, A. Identification of candidate genes responsible for innate fear behavior in the chicken. G3 2023, 13, jkac316. [Google Scholar] [CrossRef] [PubMed]
- Velasco, V.V.; Ochiai, T.; Tsudzuk, M.; Goto, N.; Ishikawa, A. Quantitative trait loci mapping of innate fear behavior in day-old F2 chickens of Japanese Oh-Shamo and White Leghorn breeds using restriction site-associated DNA sequencing. Poult. Sci. 2023, 103228. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Animals | Source and Origin of Breed 1 | |||
|---|---|---|---|---|
| Breed or Line | Abbreviation | Male | Female | |
| Chabo H line | CHB-H | 8 | 3 | JABPC |
| Chabo N line | CHB -N | 5 | 5 | ABRC [18] |
| Ingie | IG | 4 | 8 | JABPC [19] |
| Ryujin-Jidori | RYU | 11 | 13 | LPLESWP [20] |
| Oh-Shamo | OSM | 7 | 5 | JABPC [5] |
| Tosa-Kukin | TKU | 2 | 7 | JABPC [5] |
| PNP | PNP | 14 | 4 | ABRC [18] |
| Trait | Abbreviation | Criterion |
|---|---|---|
| Traits related to bustle | ||
| Number of distress vocalizations | Distress vocalization | More vocalizations indicate more clamor |
| Number of movements across the line | Moving | Number of times the bird crossed the line during the 30 s observation period after stimulation; more moves indicate more noise |
| Number of escape attempts from the cage | Escaping cage | More attempts indicate more clamor |
| Number of general vocalizations other than distress vocalizations | General vocalization | More vocalizations indicate relatively fewer distress vocalizations and less noise |
| Number of sleeps | Sleeping | Number of sleeps in each stimulus phase; more sleep indicates less bustle |
| Traits related to aggression | ||
| Number of times the floor or cage was pecked | Floor/cage pecking | More pecks indicate more aggression |
| Number of bites | Biting | More bites indicate more aggression |
| Traits related to timidity | ||
| Number of voices raised or surprised when stimulated | Surprised voice | More voices indicate more timidity |
| Number of escape attempts when stimulated | Escaping stimulus | More attempts indicate more timidity |
| Number of times approaching the cage wall | Approaching wall | More approaches indicate more timidity |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishikawa, A.; Takanuma, T.; Hashimoto, N.; Goto, T.; Tsudzuki, M. New Behavioral Handling Test Reveals Temperament Differences in Native Japanese Chickens. Animals 2023, 13, 3556. https://doi.org/10.3390/ani13223556
Ishikawa A, Takanuma T, Hashimoto N, Goto T, Tsudzuki M. New Behavioral Handling Test Reveals Temperament Differences in Native Japanese Chickens. Animals. 2023; 13(22):3556. https://doi.org/10.3390/ani13223556
Chicago/Turabian StyleIshikawa, Akira, Tomoka Takanuma, Norikazu Hashimoto, Tatsuhiko Goto, and Masaoki Tsudzuki. 2023. "New Behavioral Handling Test Reveals Temperament Differences in Native Japanese Chickens" Animals 13, no. 22: 3556. https://doi.org/10.3390/ani13223556
APA StyleIshikawa, A., Takanuma, T., Hashimoto, N., Goto, T., & Tsudzuki, M. (2023). New Behavioral Handling Test Reveals Temperament Differences in Native Japanese Chickens. Animals, 13(22), 3556. https://doi.org/10.3390/ani13223556