
The Oral Microbiome across Oral Sites in Cats with Chronic Gingivostomatitis, Periodontal Disease, and Tooth Resorption Compared with Healthy Cats


 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Ethics Statement
2.2. Clinical Evaluations of Feline Participants
2.3. Microbiome Sampling
2.4. DNA Extraction and Full-Length 16S rRNA Amplicon Gene Sequencing
2.5. Bioinformatic Processing of PacBio CSS Reads
2.6. Statistical Analyses of Microbiome data: Healthy Cats Compared with Cats Diagnosed with FCGS
2.7. Statistical Analyses of Microbiome Data: Cats Diagnosed with FCGS, TR, or SPD
2.8. Statistical Analyses of Microbiome Data: Swab Samples vs. Plaque Samples
3. Results
3.1. Feline Participants
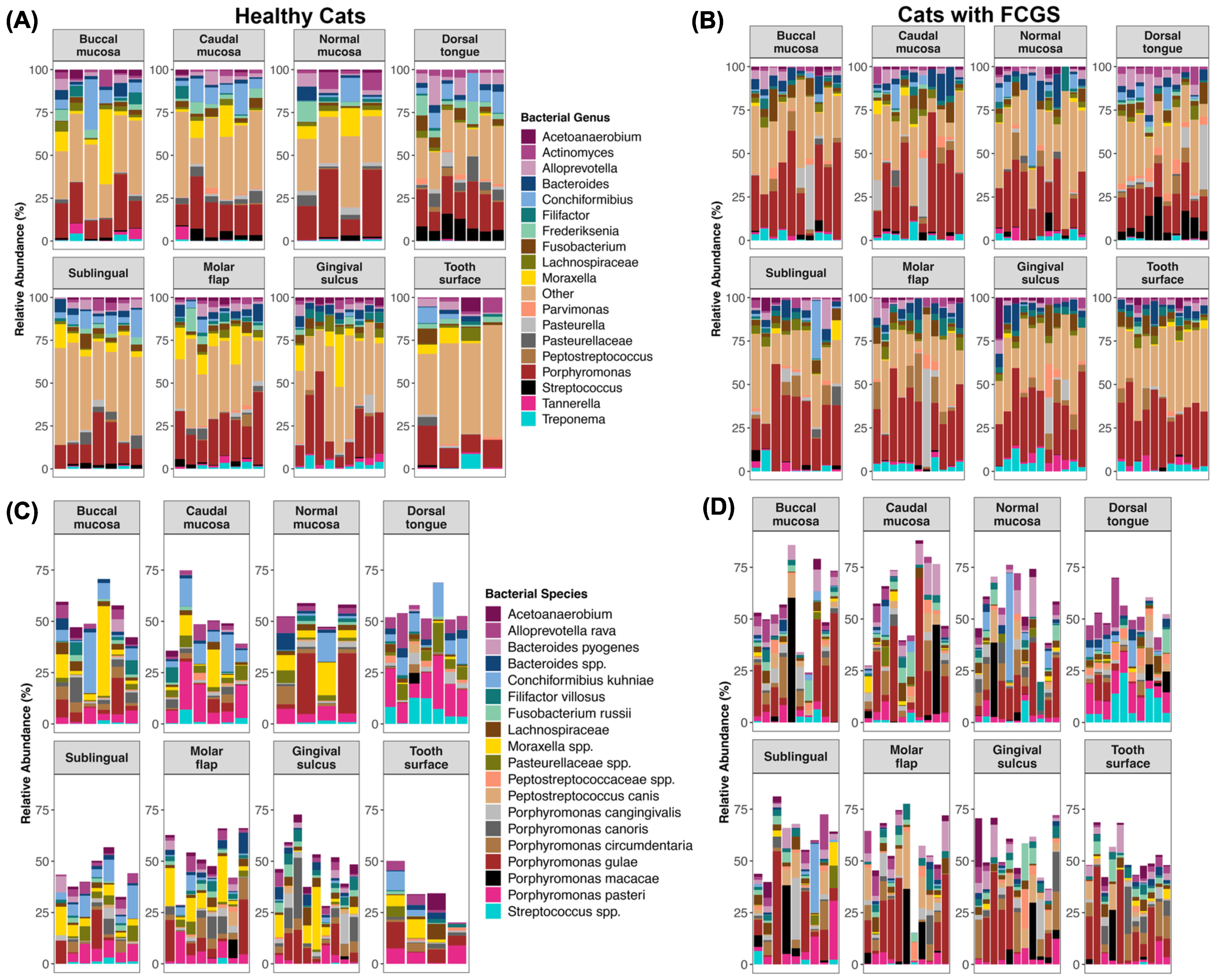
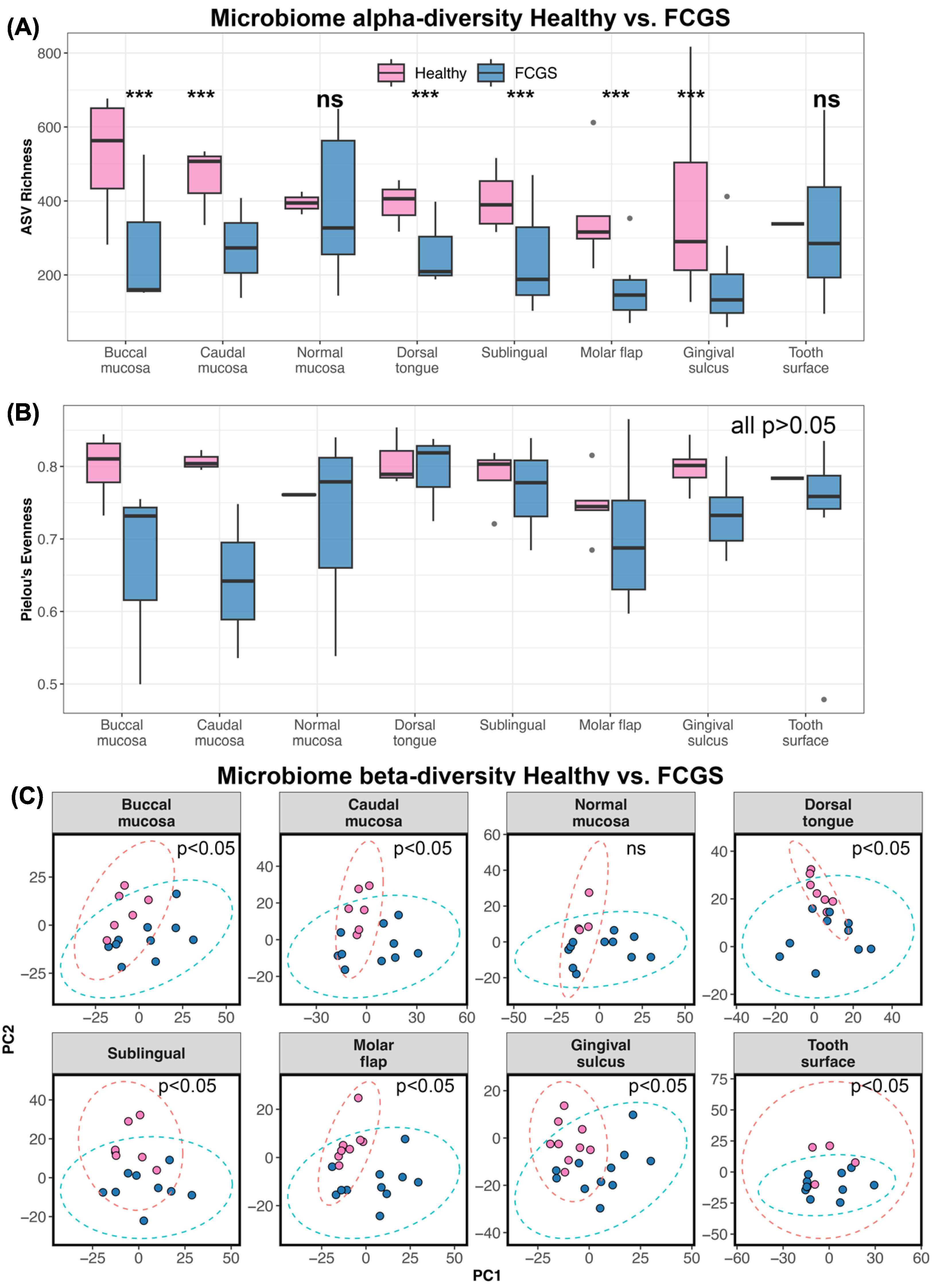
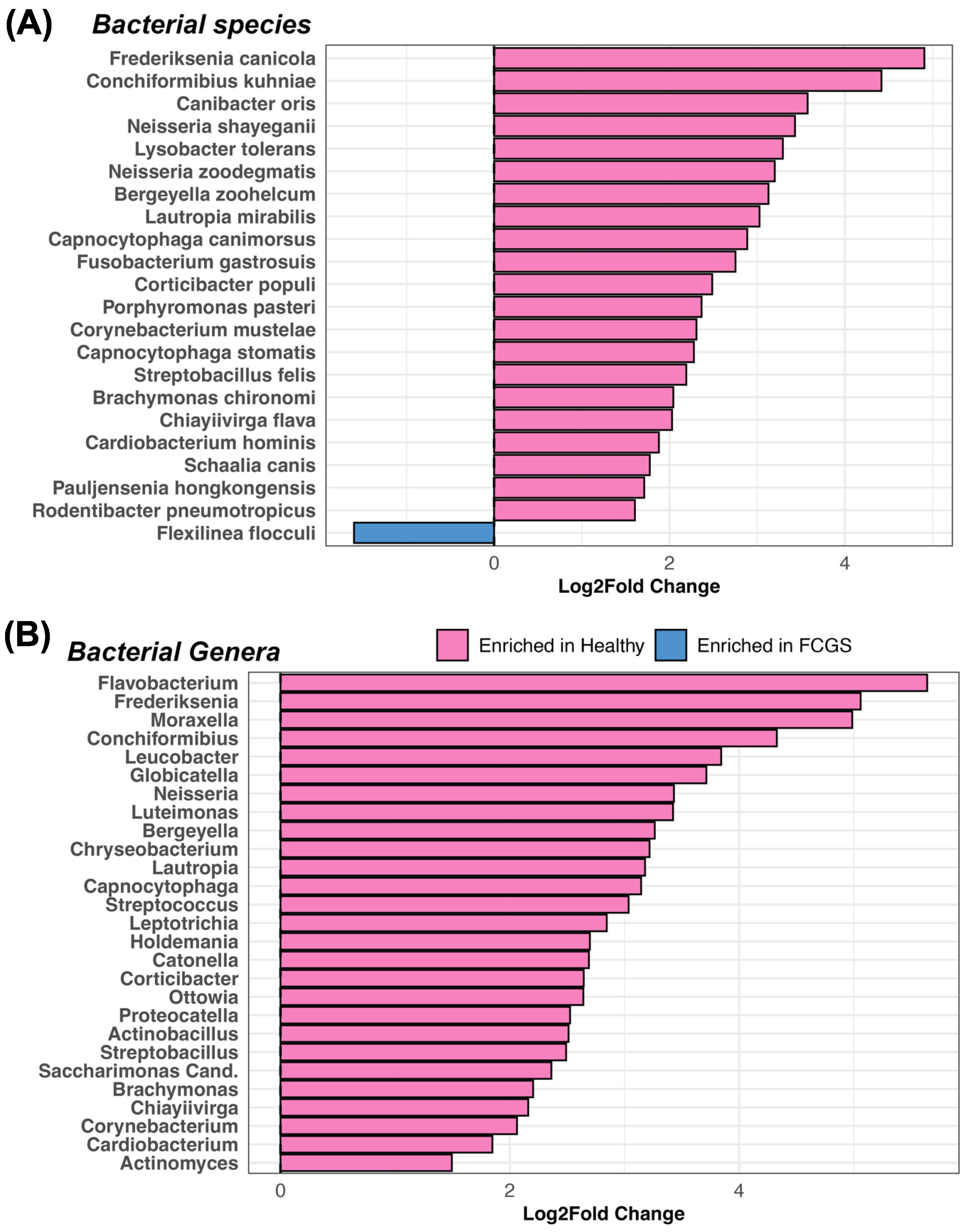
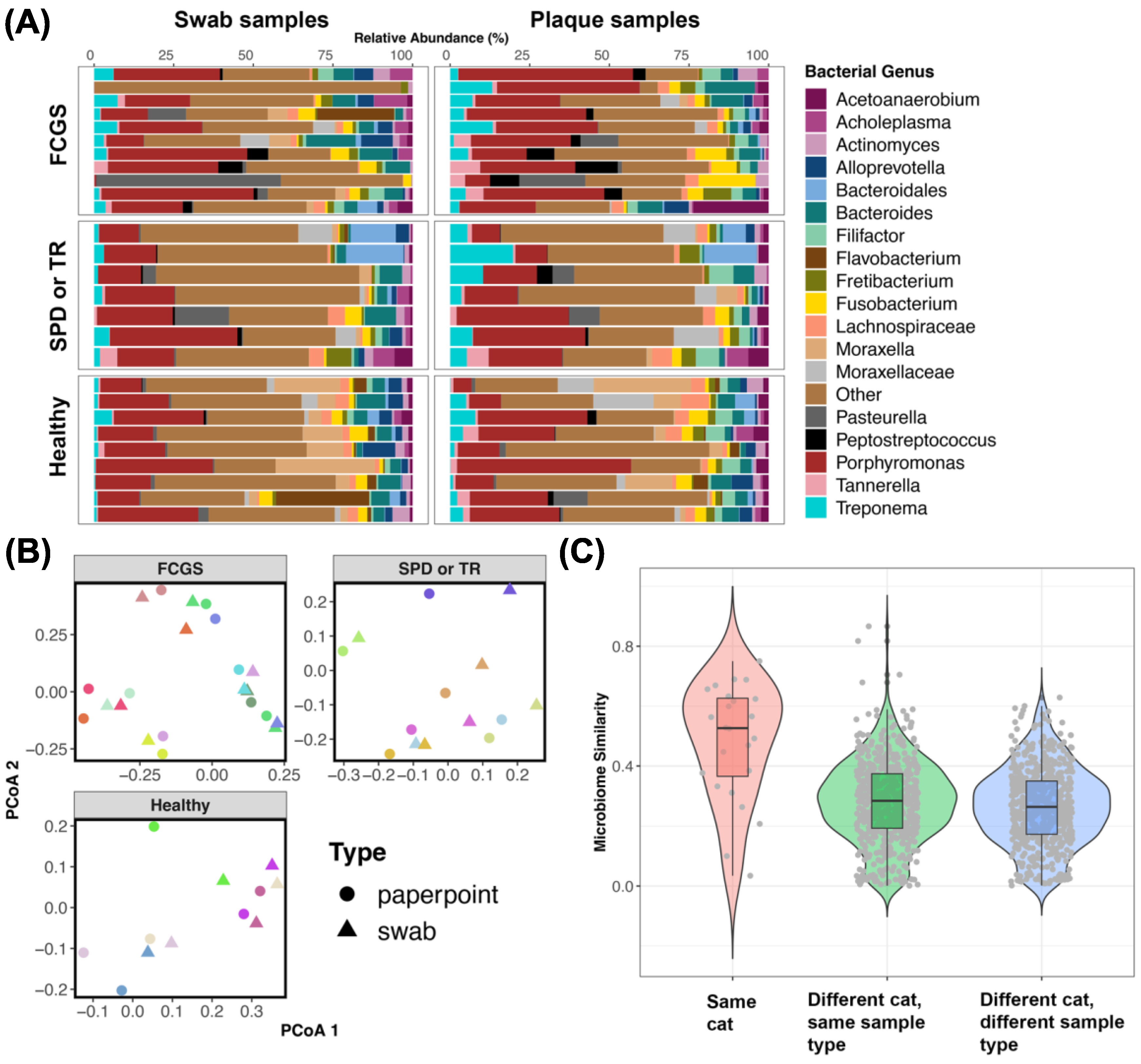
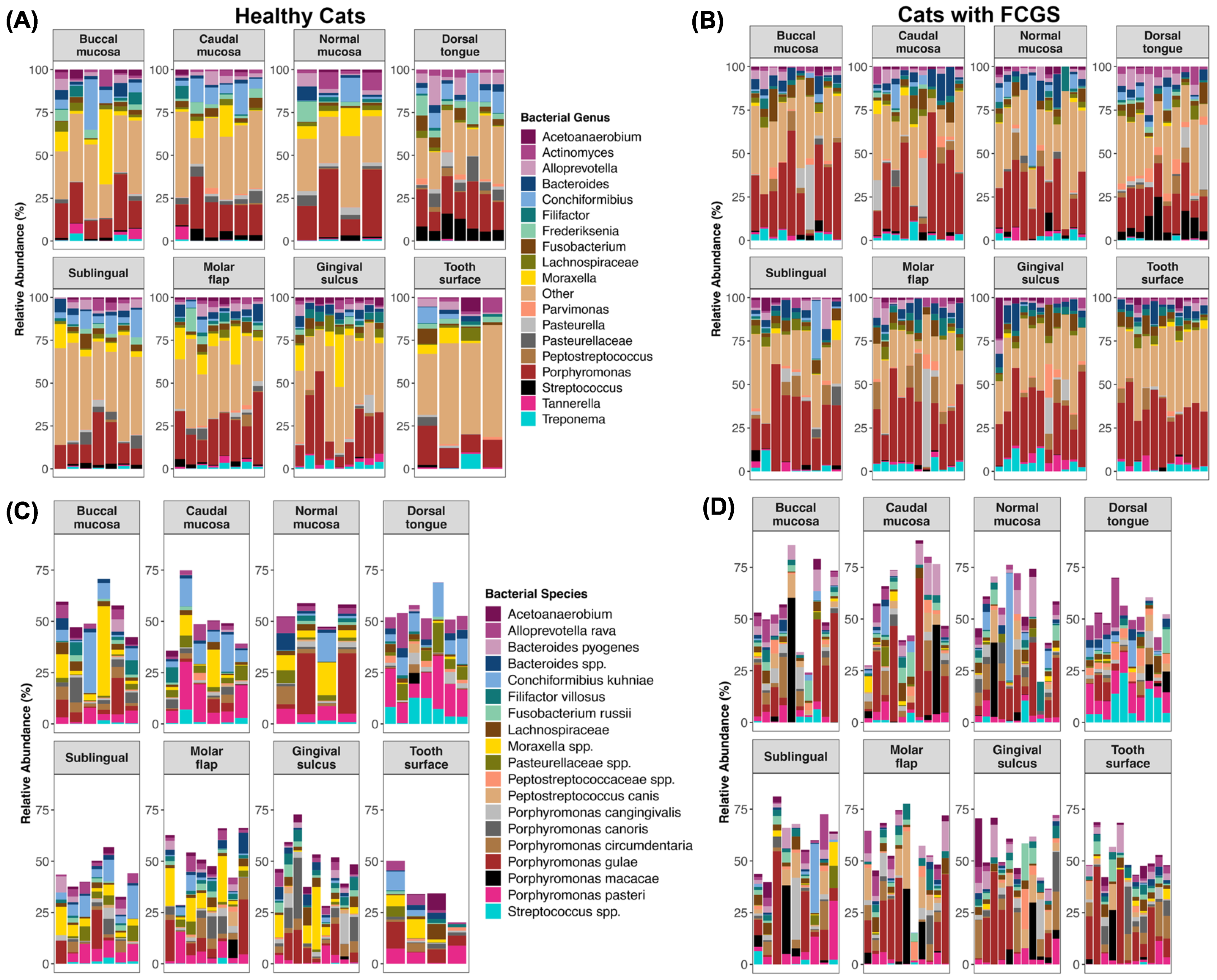
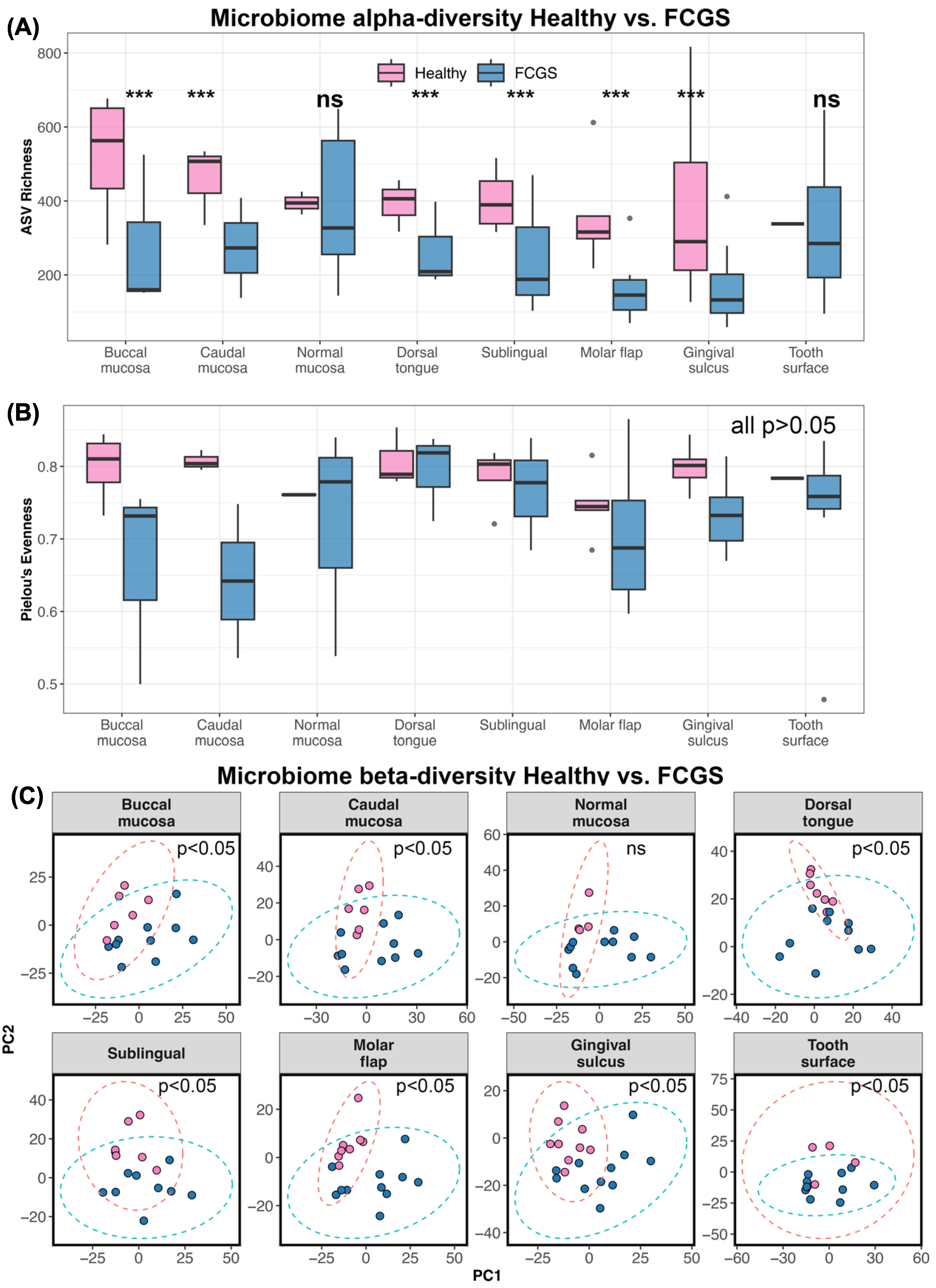
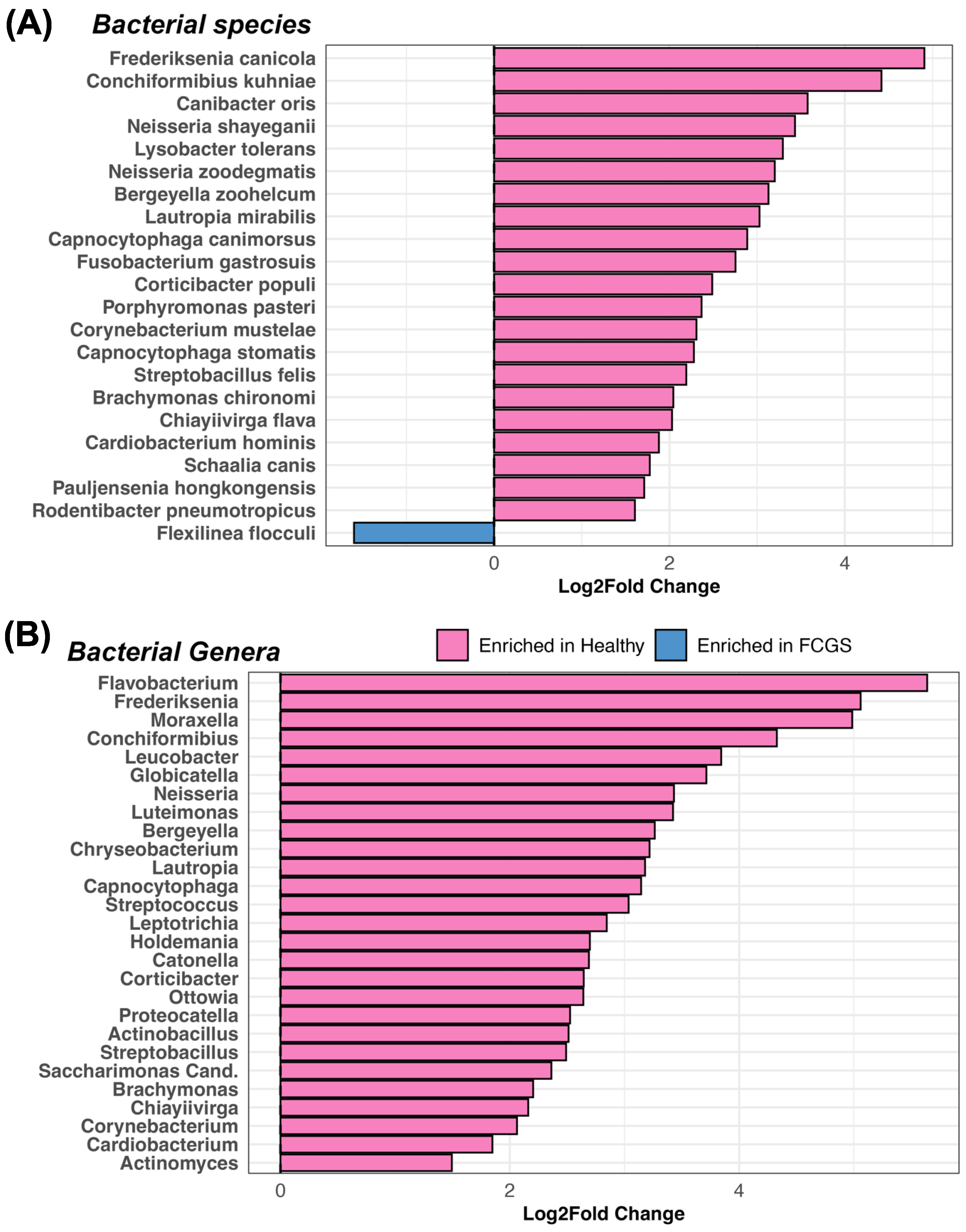
3.2. Is Microbiome Dysbiosis Observed in Cats with FCGS?
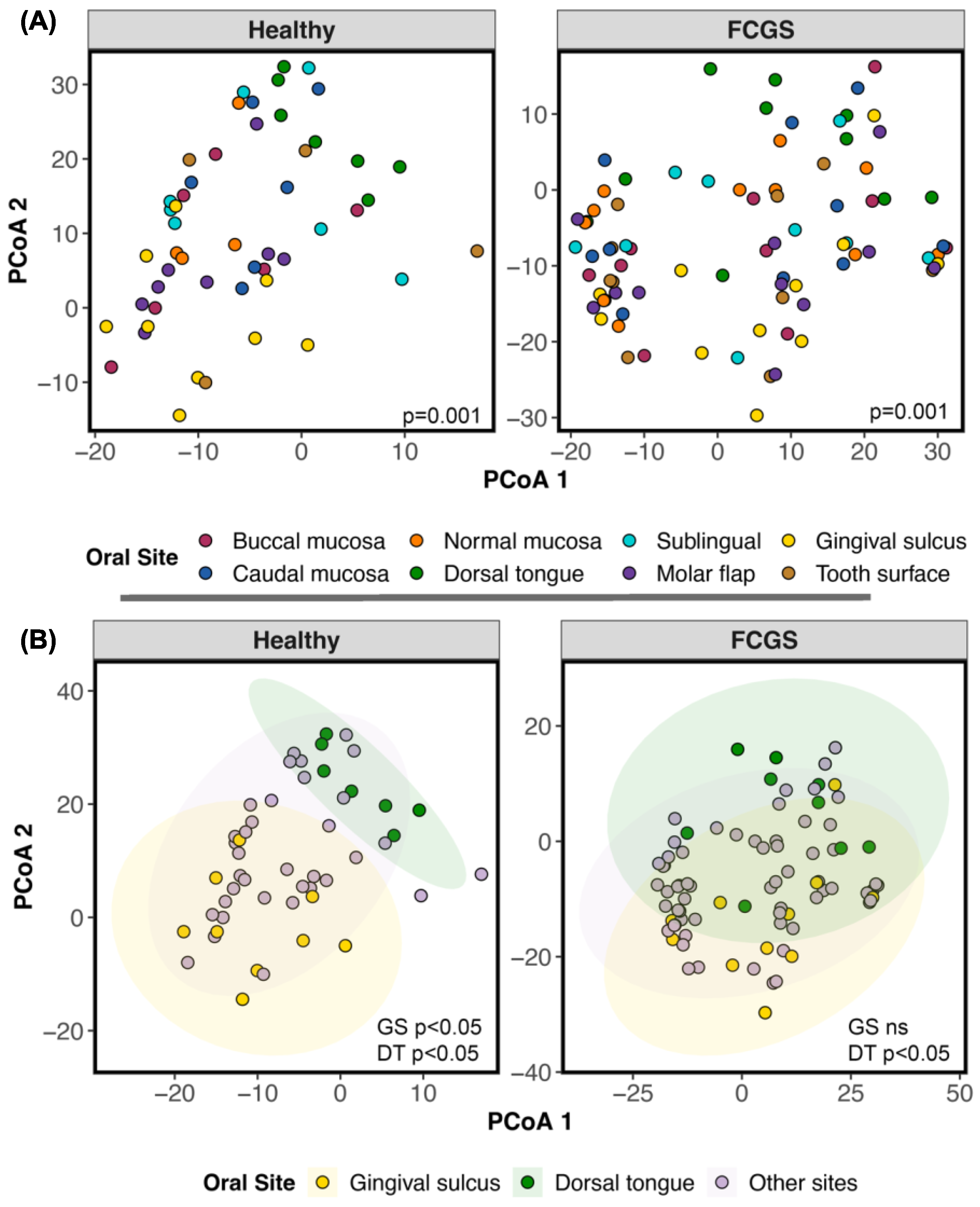
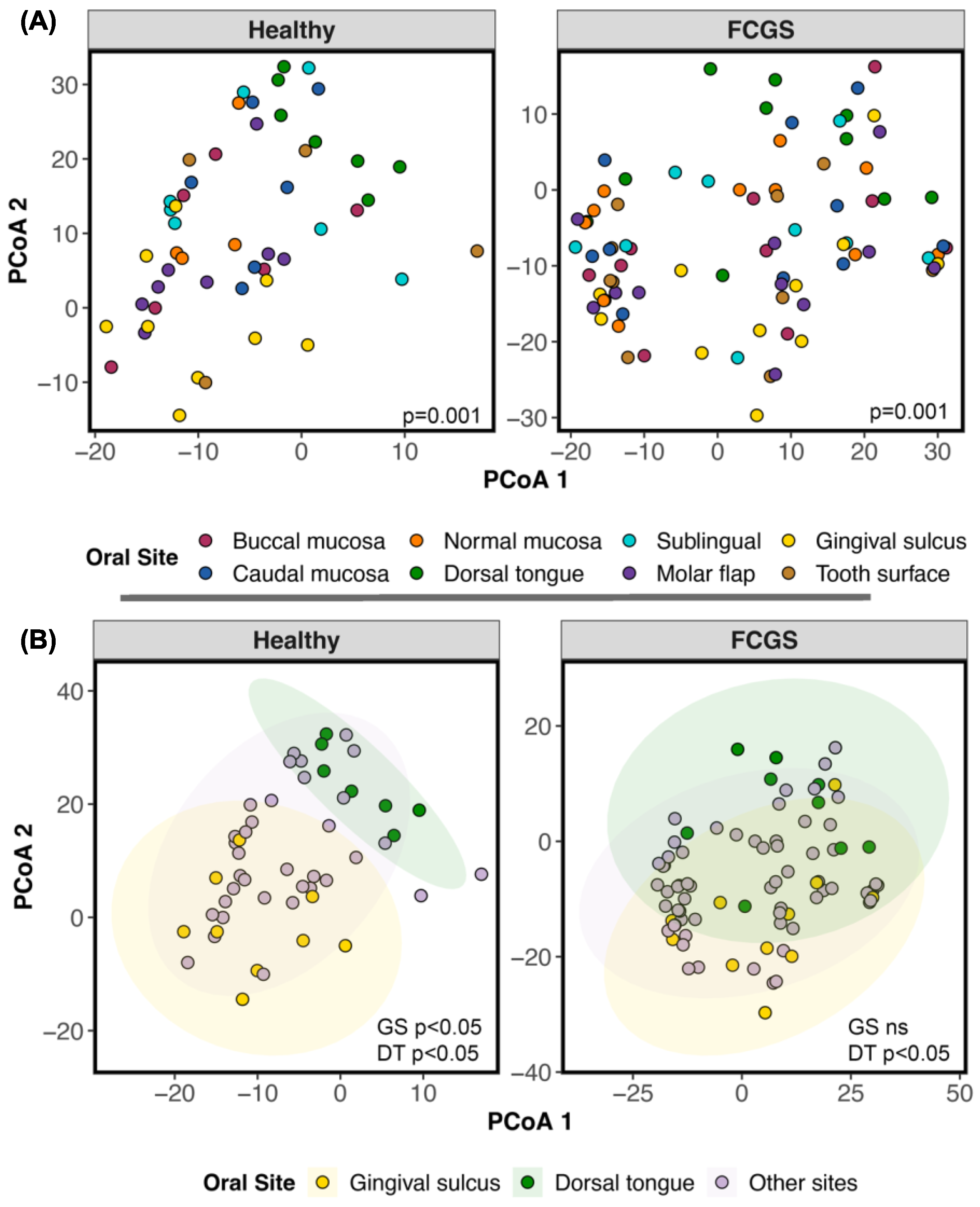
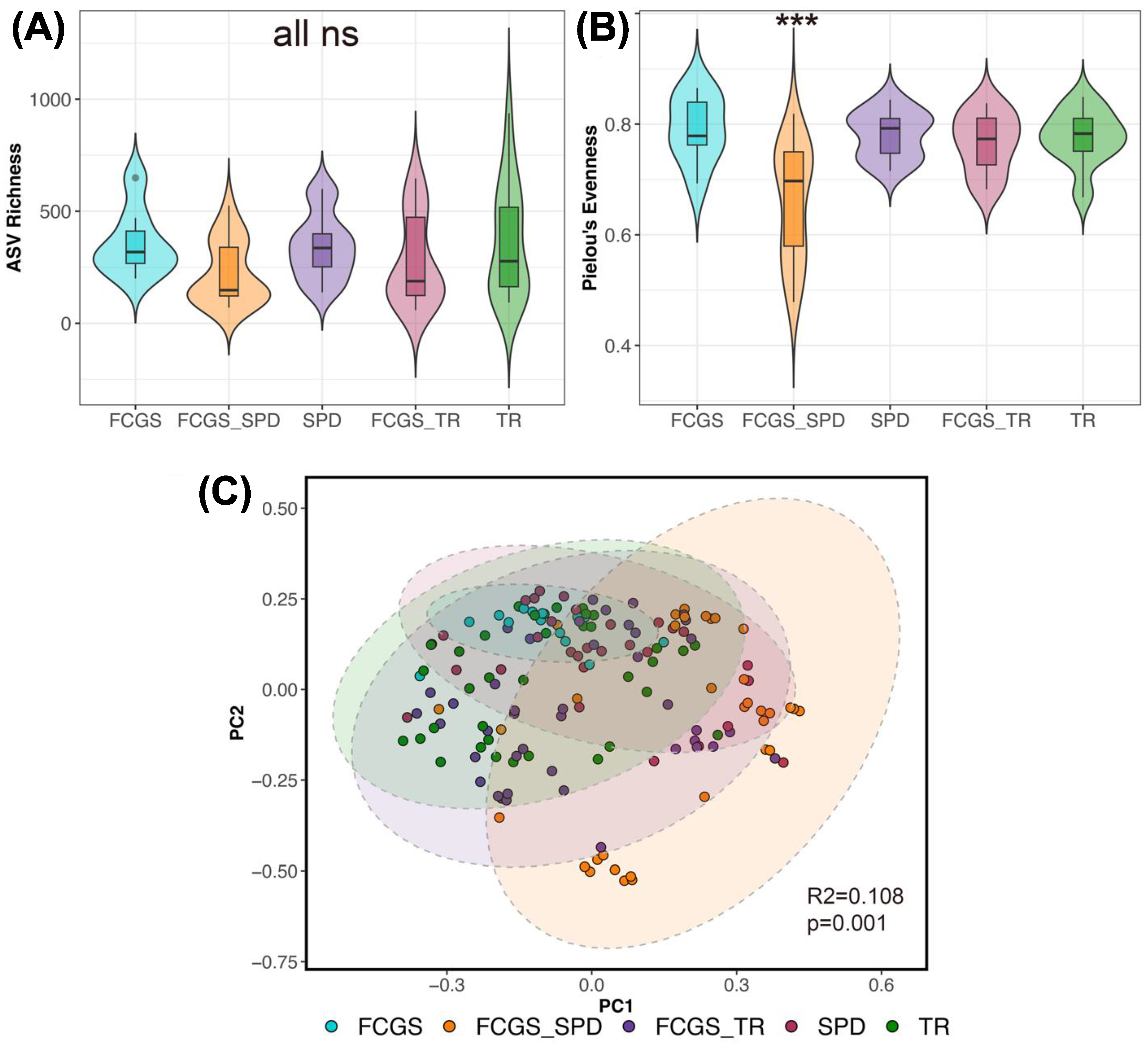
3.3. Do the Microbiomes of Cats with Different Oral Conditions Diverge from Each Other?
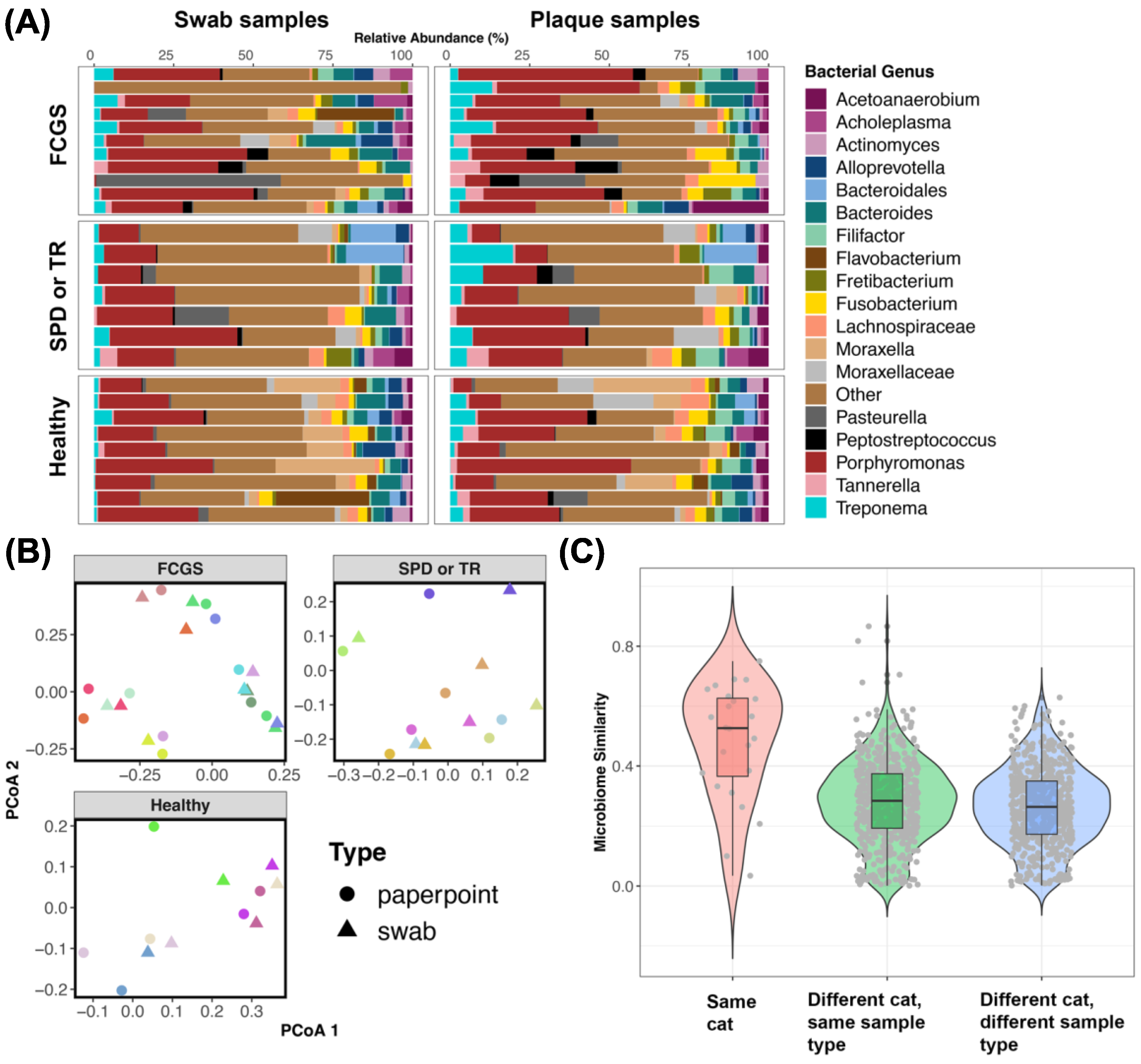
3.4. Do the Microbiomes of Swabs from the Oral Cavity Approximate Plaque Microbiomes Obtained Using Endodontic Paper Points?
4. Discussion
4.1. Bacteria That Are Abundant in the Oral Cavities of Cats with FCGS
4.2. Microbiome Diversity in Cats with FCGS Compared with Healthy Cats
4.3. Potential Bacterial Drivers of Microbiome Dysbiosis in Cats with FCGS
4.4. Differences between the Microbiomes of Cats with Gingivostomatitis, Tooth Resorption, and/or Periodontitis
4.5. Swabs of the Oral Cavity Are a Reliable Method of Sampling the Feline Microbiome
4.6. Limitations
4.7. Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, D.B.; Verstraete, F.J.M.; Arzi, B. An Update on Feline Chronic Gingivostomatitis. Vet. Clin. N. Am. Small Anim. Pract. 2020, 50, 973–982. [Google Scholar] [CrossRef]
- Lyon, K.F. Gingivostomatitis. Vet. Clin. N. Am. Small Anim. Pract. 2005, 35, 891–911. [Google Scholar] [CrossRef] [PubMed]
- Healey, K.A.E.; Dawson, S.; Burrow, R.; Cripps, P.; Gaskell, C.J.; Hart, C.A.; Pinchbeck, G.L.; Radford, A.D.; Gaskell, R.M. Prevalence of Feline Chronic Gingivo-Stomatitis in First Opinion Veterinary Practice. J. Feline Med. Surg. 2007, 9, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Bellows, J. Feline Oropharyngeal Inflammation. NAVC Clin. Brief 2010, 33–37. [Google Scholar]
- Vapniarsky, N.; Simpson, D.L.; Arzi, B.; Taechangam, N.; Walker, N.J.; Garrity, C.; Bulkeley, E.; Borjesson, D.L. Histological, Immunological, and Genetic Analysis of Feline Chronic Gingivostomatitis. Front. Vet. Sci. 2020, 7, 310. [Google Scholar] [CrossRef]
- Fried, W.A.; Soltero-Rivera, M.; Ramesh, A.; Lommer, M.J.; Arzi, B.; DeRisi, J.L.; Horst, J.A. Use of Unbiased Metagenomic and Transcriptomic Analyses to Investigate the Association between Feline Calicivirus and Feline Chronic Gingivostomatitis in Domestic Cats. Am. J. Vet. Res. 2021, 82, 381–394. [Google Scholar] [CrossRef]
- Knowles, J.O.; Gaskell, R.M.; Gaskell, C.J.; Harvey, C.E.; Lutz, H. Prevalence of Feline Calicivirus, Feline Leukaemia Virus and Antibodies to FIV in Cats with Chronic Stomatitis. Vet. Rec. 1989, 124, 336–338. [Google Scholar] [CrossRef]
- Belgard, S.; Truyen, U.; Thibault, J.-C.; Sauter-Louis, C.; Hartmann, K. Relevance of Feline Calicivirus, Feline Immunodeficiency Virus, Feline Leukemia Virus, Feline Herpesvirus and Bartonella Henselae in Cats with Chronic Gingivostomatitis. Berl. Munch. Tierarztl. Wochenschr. 2010, 123, 369–376. [Google Scholar]
- Villatoro, A.J.; Martín-Astorga, M.D.C.; Alcoholado, C.; Kazantseva, L.; Cárdenas, C.; Fariñas, F.; Becerra, J.; Visser, R. Secretory Profile of Adipose-Tissue-Derived Mesenchymal Stem Cells from Cats with Calicivirus-Positive Severe Chronic Gingivostomatitis. Viruses 2022, 14, 1146. [Google Scholar] [CrossRef]
- Peralta, S.; Grenier, J.K.; Webb, S.M.; Miller, A.D.; Miranda, I.C.; Parker, J.S.L. Transcriptomic Signatures of Feline Chronic Gingivostomatitis Are Influenced by Upregulated IL6. Res. Sq. 2023. [Google Scholar] [CrossRef]
- Rodrigues, M.X.; Bicalho, R.C.; Fiani, N.; Lima, S.F.; Peralta, S. The Subgingival Microbial Community of Feline Periodontitis and Gingivostomatitis: Characterization and Comparison between Diseased and Healthy Cats. Sci. Rep. 2019, 9, 12340. [Google Scholar] [CrossRef] [PubMed]
- Krumbeck, J.A.; Reiter, A.M.; Pohl, J.C.; Tang, S.; Kim, Y.J.; Linde, A.; Prem, A.; Melgarejo, T. Characterization of Oral Microbiota in Cats: Novel Insights on the Potential Role of Fungi in Feline Chronic Gingivostomatitis. Pathogens 2021, 10, 904. [Google Scholar] [CrossRef] [PubMed]
- Older, C.E.; Gomes, M.d.O.S.; Hoffmann, A.R.; Policano, M.D.; Reis, C.A.C.D.; Carregaro, A.B.; Ambrósio, C.E.; Carregaro, V.M.L. Influence of the FIV Status and Chronic Gingivitis on Feline Oral Microbiota. Pathogens 2020, 9, 383. [Google Scholar] [CrossRef] [PubMed]
- Riviere, G.R.; Thompson, A.J.; Brannan, R.D.; McCoy, D.E.; Simonson, L.G. Detection of Pathogen-Related Oral Spirochetes, Treponema Denticola, and Treponema Socranskii in Dental Plaque from Dogs. J. Vet. Dent. 1996, 13, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, Y.; Ishihara, K. Characterization of the Treponema Denticola Virulence Factor Dentilisin. Methods Mol. Biol. 2021, 2210, 173–184. [Google Scholar] [PubMed]
- Niemiec, B.A.; Gawor, J.; Tang, S.; Prem, A.; Krumbeck, J.A. The Bacteriome of the Oral Cavity in Healthy Dogs and Dogs with Periodontal Disease. Am. J. Vet. Res. 2021, 83, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.G.; Paster, B.J.; Kokaras, A.; Chen, T. Characterization of the Oral Microbiome in Canine Chronic Ulcerative Stomatitis. J. Immun. Res. 2021, 7, 1037. [Google Scholar]
- Bellows, J.; Berg, M.L.; Dennis, S.; Harvey, R.; Lobprise, H.B.; Snyder, C.J.; Stone, A.E.S.; Van de Wetering, A.G. 2019 AAHA Dental Care Guidelines for Dogs and Cats. J. Am. Anim. Hosp. Assoc. 2019, 55, 49–69. [Google Scholar] [CrossRef]
- Istace, K. Comprehensive Oral Health Assessment and Treatment (COHAT); Dental Instrument Use and Maintenance. In An Introduction to Pet Dental Care: For Veterinary Nurses and Technicians; CABI: Wallingford, UK, 2021; pp. 14–44. ISBN 9781789248869. [Google Scholar]
- Soltero-Rivera, M.; Vapniarsky, N.; Rivas, I.L.; Arzi, B. Clinical, Radiographic and Histopathologic Features of Early-Onset Gingivitis and Periodontitis in Cats (1997–2022). J. Feline Med. Surg. 2023, 25, 1098612X221148577. [Google Scholar] [CrossRef]
- Farcas, N.; Lommer, M.J.; Kass, P.H.; Verstraete, F.J.M. Dental Radiographic Findings in Cats with Chronic Gingivostomatitis (2002–2012). J. Am. Vet. Med. Assoc. 2014, 244, 339–345. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A Versatile Open Source Tool for Metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Villanueva, R.A.M.; Chen, Z.J. ggplot2: Elegant Graphics for Data Analysis (2nd Ed.). Measurement 2019, 17, 160–167. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, X.; Zhou, H. GUniFrac: Generalized UniFrac Distances, Distance-Based Multivariate Methods and Feature-Based Univariate Methods for Microbiome Data Analysis. R package. Available online: https://cran.r-project.org/web/packages/GUniFrac/index.html (accessed on 14 September 2023).
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Lahti, L.; Shetty, S.; Turaga, N.; Wokaty, J.; Leung, E.; Gilmore, R.; Salojärvi, J.; Blake, T.; van der Plas, D.; Obenchain, V.; et al. Introduction to the Microbiome R Package. R package. 2018. Available online: https://microbiome.github.io/tutorials (accessed on 3 August 2023).
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package 2022. R package. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 3 August 2023).
- Martinez Arbizu, P. pairwiseAdonis: Pairwise Multilevel Comparison Using Adonis 2017. R package. Available online: https://github.com/pmartinezarbizu/pairwiseAdonis/blob/master/pairwiseAdonis/R/pairwise.adonis.R (accessed on 3 August 2023).
- Zhou, H.; He, K.; Chen, J.; Zhang, X. LinDA: Linear Models for Differential Abundance Analysis of Microbiome Compositional Data. Genome Biol. 2022, 23, 95. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R.V. Emmeans: Estimated Marginal Means, Aka Least-Squares Means 2023. Available online: https://github.com/rvlenth/emmeans (accessed on 17 October 2023).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Gaskell, R.M.; Tj, G.J. Intractable Feline Stomatitis. Vet. Annu. 1997, 17, 195–199. [Google Scholar]
- Pérez-Salcedo, L.; Herrera, D.; Esteban-Saltiveri, D.; León, R.; Jeusette, I.; Torre, C.; O’Connor, A.; González, I.; González, I. Isolation and Identification of Porphyromonas Spp. and Other Putative Pathogens from Cats with Periodontal Disease. J. Vet. Dent. 2013, 30, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Lenzo, J.C.; O’Brien-Simpson, N.M.; Orth, R.K.; Mitchell, H.L.; Dashper, S.G.; Reynolds, E.C. Porphyromonas Gulae Has Virulence and Immunological Characteristics Similar to Those of the Human Periodontal Pathogen Porphyromonas Gingivalis. Infect. Immun. 2016, 84, 2575–2585. [Google Scholar] [CrossRef] [PubMed]
- Love, D.N.; Johnson, J.L.; Moore, L.V. Bacteroides Species from the Oral Cavity and Oral-Associated Diseases of Cats. Vet. Microbiol. 1989, 19, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Abrahamian, F.M.; Goldstein, E.J.C. Microbiology of Animal Bite Wound Infections. Clin. Microbiol. Rev. 2011, 24, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A. Persistence of Tannerella Forsythia and Fusobacterium Nucleatum in Dental Plaque: A Strategic Alliance. Curr. Oral Health Rep. 2020, 7, 22–28. [Google Scholar] [CrossRef]
- Özavci, V.; Erbas, G.; Parin, U.; Yüksel, H.T.; Kirkan, Ş. Molecular Detection of Feline and Canine Periodontal Pathogens. Vet. Anim. Sci. 2019, 8, 100069. [Google Scholar] [CrossRef]
- Kwon, D.; Bae, K.; Kim, H.; Kim, S.-H.; Lee, D.; Lee, J.-H. Treponema Denticola as a Prognostic Biomarker for Periodontitis in Dogs. PLoS ONE 2022, 17, e0262859. [Google Scholar] [CrossRef]
- Loesche, W.J.; Grossman, N.S. Periodontal Disease as a Specific, Albeit Chronic, Infection: Diagnosis and Treatment. Clin. Microbiol. Rev. 2001, 14, 727–752. [Google Scholar] [CrossRef]
- Sela, M.N. Role of Treponema Denticola in Periodontal Diseases. Crit. Rev. Oral Biol. Med. 2001, 12, 399–413. [Google Scholar] [CrossRef]
- Dashper, S.G.; Seers, C.A.; Tan, K.H.; Reynolds, E.C. Virulence Factors of the Oral Spirochete Treponema Denticola. J. Dent. Res. 2011, 90, 691–703. [Google Scholar] [CrossRef]
- Dolieslager, S.M.J.; Riggio, M.P.; Lennon, A.; Lappin, D.F.; Johnston, N.; Taylor, D.; Bennett, D. Identification of Bacteria Associated with Feline Chronic Gingivostomatitis Using Culture-Dependent and Culture-Independent Methods. Vet. Microbiol. 2011, 148, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Lappin, D.F.; Nile, C.J.; Spears, J.; Bennett, D.; Brandt, B.W.; Riggio, M.P. Microbiome Analysis of Feline Odontoclastic Resorptive Lesion (FORL) and Feline Oral Health. J. Med. Microbiol. 2021, 70. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.; Jang, S. Oral Microbiome Research on Oral Lichen Planus: Current Findings and Perspectives. Biology 2022, 11, 723. [Google Scholar] [CrossRef] [PubMed]
- Santibáñez, R.; Rodríguez-Salas, C.; Flores-Yáñez, C.; Garrido, D.; Thomson, P. Assessment of Changes in the Oral Microbiome That Occur in Dogs with Periodontal Disease. Vet. Sci. China 2021, 8, 291. [Google Scholar] [CrossRef]
- Harris, S.; Croft, J.; O’Flynn, C.; Deusch, O.; Colyer, A.; Allsopp, J.; Milella, L.; Davis, I.J. A Pyrosequencing Investigation of Differences in the Feline Subgingival Microbiota in Health, Gingivitis and Mild Periodontitis. PLoS ONE 2015, 10, e0136986. [Google Scholar] [CrossRef]
- Borsanelli, A.C.; Lappin, D.F.; Viora, L.; Bennett, D.; Dutra, I.S.; Brandt, B.W.; Riggio, M.P. Microbiomes Associated with Bovine Periodontitis and Oral Health. Vet. Microbiol. 2018, 218, 1–6. [Google Scholar] [CrossRef]
- Rodrigues, M.X.; Fiani, N.; Bicalho, R.C.; Peralta, S. Preliminary Functional Analysis of the Subgingival Microbiota of Cats with Periodontitis and Feline Chronic Gingivostomatitis. Sci. Rep. 2021, 11, 6896. [Google Scholar] [CrossRef]
- Holt, S.C. Capnocytophaga. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 1–13. [Google Scholar]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef]
- Sakamoto, M.; Li, D.; Shibata, Y.; Takeshita, T.; Yamashita, Y.; Ohkuma, M. Porphyromonas Pasteri Sp. Nov., Isolated from Human Saliva. Int. J. Syst. Evol. Microbiol. 2015, 65, 2511–2515. [Google Scholar] [CrossRef]
- Yasunaga, H.; Takeshita, T.; Shibata, Y.; Furuta, M.; Shimazaki, Y.; Akifusa, S.; Ninomiya, T.; Kiyohara, Y.; Takahashi, I.; Yamashita, Y. Exploration of Bacterial Species Associated with the Salivary Microbiome of Individuals with a Low Susceptibility to Dental Caries. Clin. Oral Investig. 2017, 21, 2399–2406. [Google Scholar] [CrossRef]
- Holcombe, L.J.; Patel, N.; Colyer, A.; Deusch, O.; O’Flynn, C.; Harris, S. Early Canine Plaque Biofilms: Characterization of Key Bacterial Interactions Involved in Initial Colonization of Enamel. PLoS ONE 2014, 9, e113744. [Google Scholar] [CrossRef] [PubMed]
- Dahle, U.R.; Tronstad, L.; Olsen, I. Spirochaetes in Oral Infections. Endod. Dent. Traumatol. 1993, 9, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Toyonaga, M.; Ohashi, A.; Matsuura, N.; Tourlousse, D.M.; Meng, X.-Y.; Tamaki, H.; Hanada, S.; Cruz, R.; Yamaguchi, T.; et al. Isolation and Characterization of Flexilinea Flocculi Gen. Nov., Sp. Nov., a Filamentous, Anaerobic Bacterium Belonging to the Class Anaerolineae in the Phylum Chloroflexi. Int. J. Syst. Evol. Microbiol. 2016, 66, 988–996. [Google Scholar] [CrossRef] [PubMed]
- Borsanelli, A.C.; Athayde, F.R.F.; Riggio, M.P.; Brandt, B.W.; Rocha, F.I.; Jesus, E.C.; Gaetti-Jardim, E., Jr.; Schweitzer, C.M.; Dutra, I.S. Dysbiosis and Predicted Function of Dental and Ruminal Microbiome Associated with Bovine Periodontitis. Front. Microbiol. 2022, 13, 936021. [Google Scholar] [CrossRef]
- Anderson, J.G. Periodontal and radiographic findings in 100 cases of feline gingivostomatitis. In Proceedings of the 20th Veterinary Dental Forum, Portland, OR, USA, 22 September 2006. [Google Scholar]
- Hajishengallis, G.; Abe, T.; Maekawa, T.; Hajishengallis, E.; Lambris, J.D. Role of Complement in Host-Microbe Homeostasis of the Periodontium. Semin. Immunol. 2013, 25, 65–72. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy (n = 9 but 3 NAs) | SPD (n = 5) | TR (n = 6, but 1 NA) | FCGS (n = 12, but 1 NA) | All (n = 32 but 5 NAs) | |
|---|---|---|---|---|---|
| Age (years) | 4.6 | 5 | 9 | 5 | 6.4 |
| Sex (% male) | 43 | 60 | 40 | 45 | 46 |
| Body weight (kg) | 4.4 | 5.5 | 5.5 | 4.6 | 4.75 |
| Swabs of Oral Cavity | Plaque Samples of the Gingival Sulcus | |||
|---|---|---|---|---|
| Predictor | Bray–Curtis (n = 25) | Aitchison (n = 25) | Bray–Curtis (n = 25) | Aitchison (n = 25) |
| Age (years) | R2 = 0.042, p = 0.31 | R2 = 0.03, p = 0.83 | R2 = 0.04, p = 0.29 | R2 = 0.036, p = 0.63 |
| Sex (M/F) | R2 = 0.15, p = 0.01 | R2 = 0.136, p = 0.012 | R2 = 0.035, p = 0.67 | R2 = 0.041, p = 0.46 |
| Body weight (kg) | R2 = 0.049, p = 0.15 | R2 = 0.04, p = 0.407 | R2 = 0.43, p = 0.39 | R2 = 0.042, p = 0.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anderson, J.G.; Rojas, C.A.; Scarsella, E.; Entrolezo, Z.; Jospin, G.; Hoffman, S.L.; Force, J.; MacLellan, R.H.; Peak, M.; Shope, B.H.; et al. The Oral Microbiome across Oral Sites in Cats with Chronic Gingivostomatitis, Periodontal Disease, and Tooth Resorption Compared with Healthy Cats. Animals 2023, 13, 3544. https://doi.org/10.3390/ani13223544
Anderson JG, Rojas CA, Scarsella E, Entrolezo Z, Jospin G, Hoffman SL, Force J, MacLellan RH, Peak M, Shope BH, et al. The Oral Microbiome across Oral Sites in Cats with Chronic Gingivostomatitis, Periodontal Disease, and Tooth Resorption Compared with Healthy Cats. Animals. 2023; 13(22):3544. https://doi.org/10.3390/ani13223544
Chicago/Turabian StyleAnderson, Jamie G., Connie A. Rojas, Elisa Scarsella, Zhandra Entrolezo, Guillaume Jospin, Sharon L. Hoffman, Judy Force, Roxane H. MacLellan, Mike Peak, Bonnie H. Shope, and et al. 2023. "The Oral Microbiome across Oral Sites in Cats with Chronic Gingivostomatitis, Periodontal Disease, and Tooth Resorption Compared with Healthy Cats" Animals 13, no. 22: 3544. https://doi.org/10.3390/ani13223544
APA StyleAnderson, J. G., Rojas, C. A., Scarsella, E., Entrolezo, Z., Jospin, G., Hoffman, S. L., Force, J., MacLellan, R. H., Peak, M., Shope, B. H., Tsugawa, A. J., & Ganz, H. H. (2023). The Oral Microbiome across Oral Sites in Cats with Chronic Gingivostomatitis, Periodontal Disease, and Tooth Resorption Compared with Healthy Cats. Animals, 13(22), 3544. https://doi.org/10.3390/ani13223544