
The Effects of Fermented Feed on the Growth Performance, Antioxidant Activity, Immune Function, Intestinal Digestive Enzyme Activity, Morphology, and Microflora of Yellow-Feather Chickens


 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment Material
2.2. Experiment Design and Sample Collection
2.3. Statistical Analysis of Data
3. Results
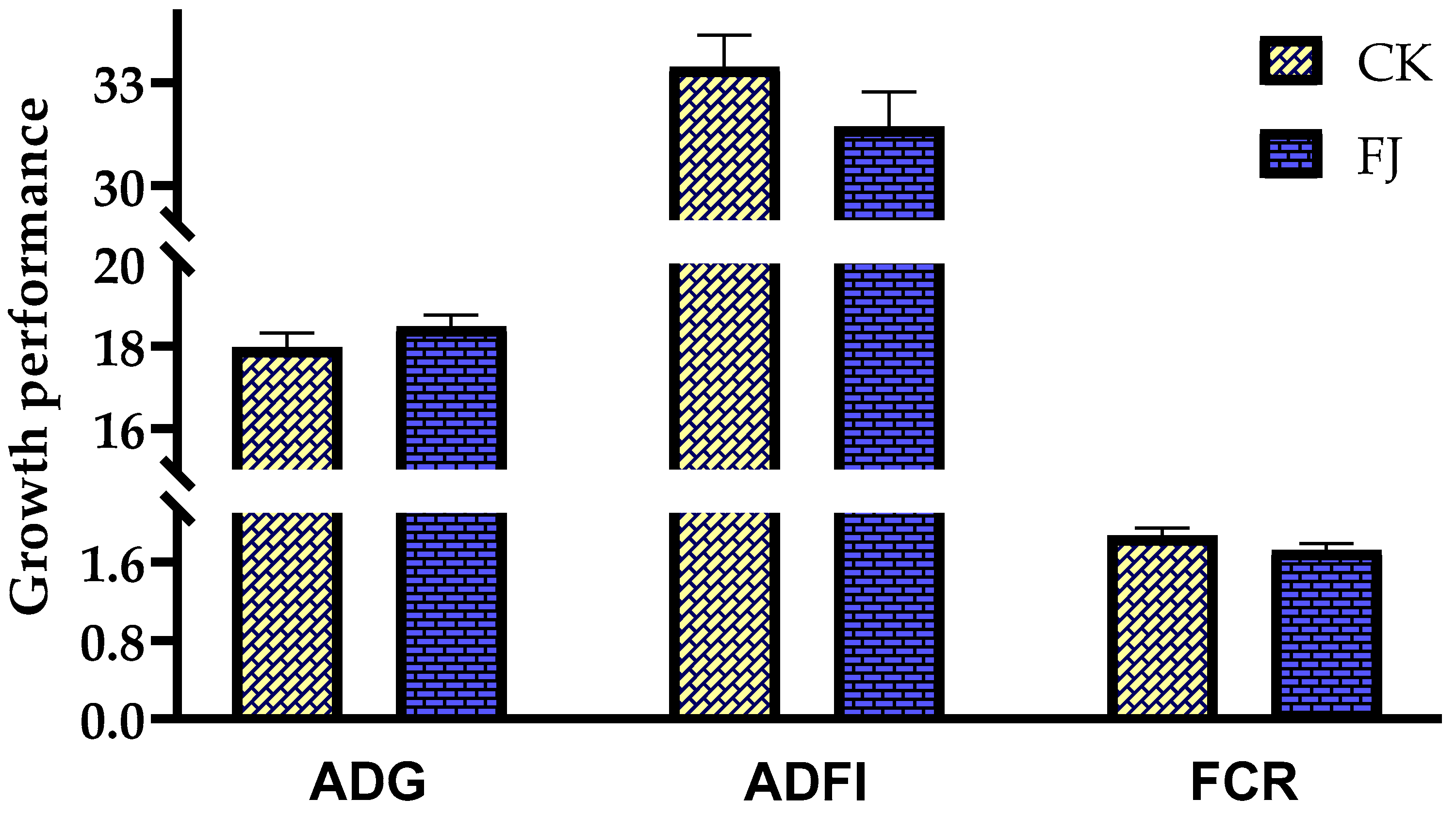
3.1. Effects of Fermented Feed on Growth Performance in Chicks
3.2. Effects of Fermented Feed on Serum Immune Function of Chickens
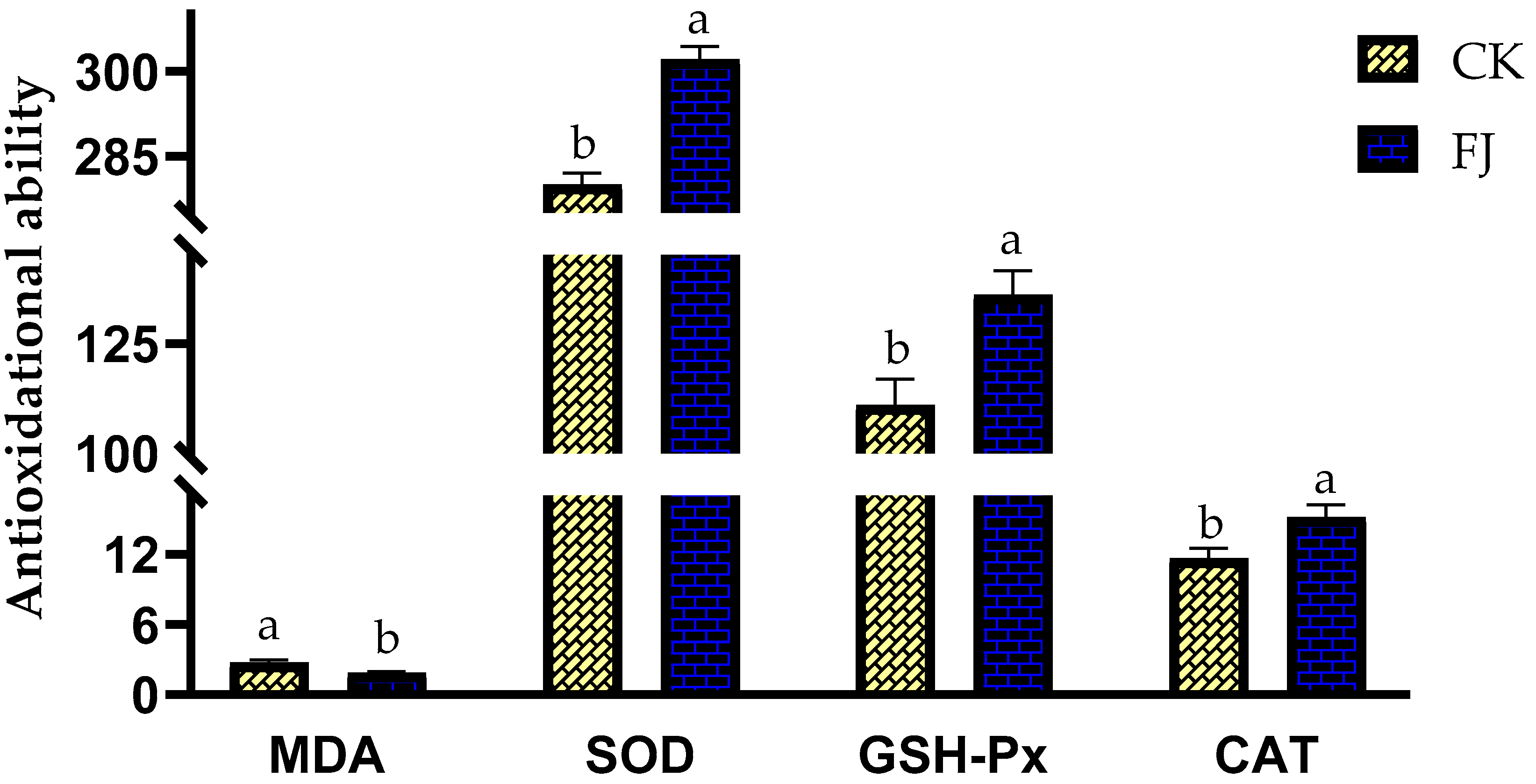
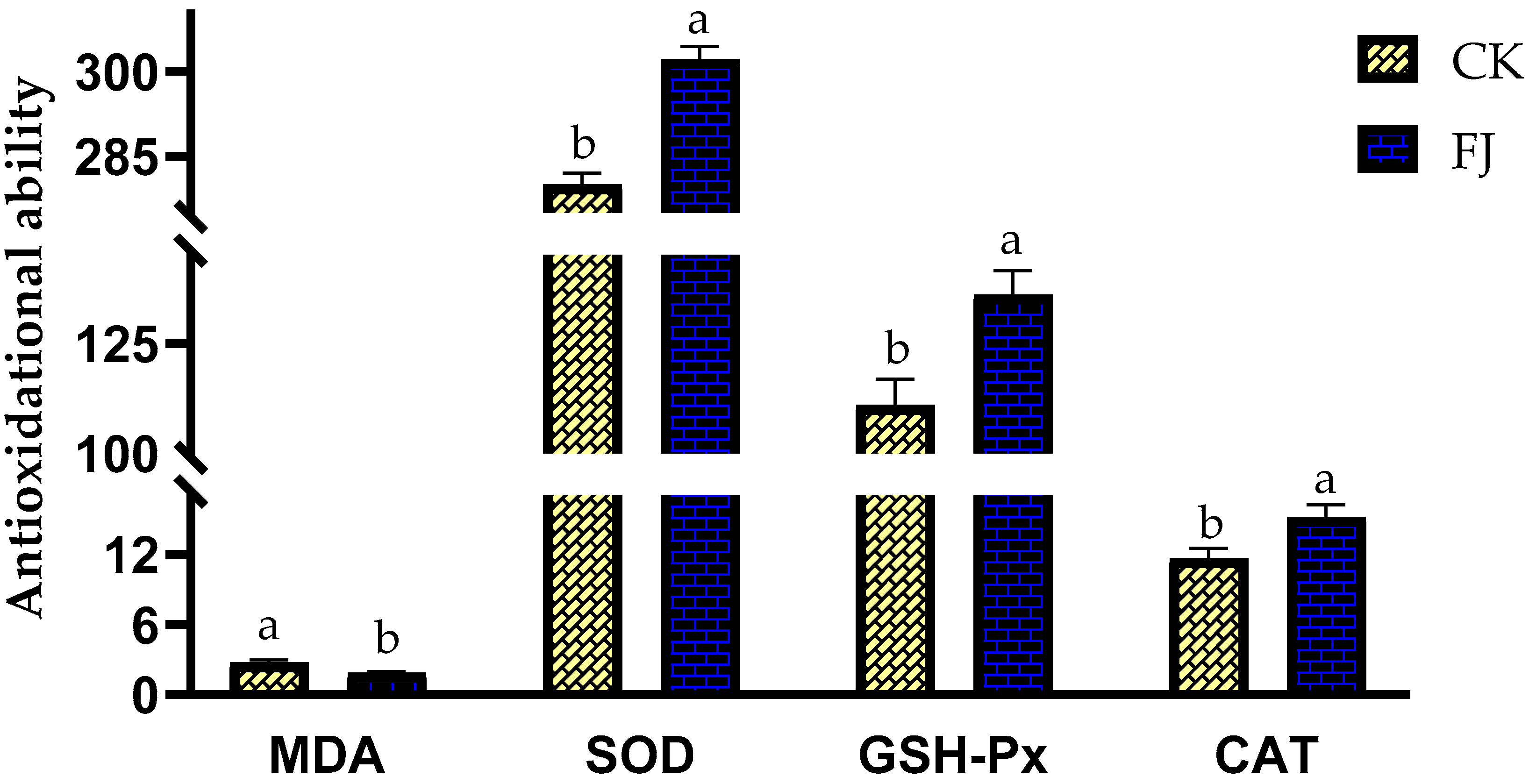
3.3. Effects of Fermented Feed on Antioxidational Ability in the Liver of Chickens
3.4. Effects of Fermented Feed on Digestive Enzyme Activity in Chickens
3.5. Effects of Fermented Feed on Intestinal Tissue Morphology of Chickens
3.6. Effects of Fermented Feed on Microbial Diversity in the Cecum of Chickens
3.6.1. Alpha Diversity Analysis
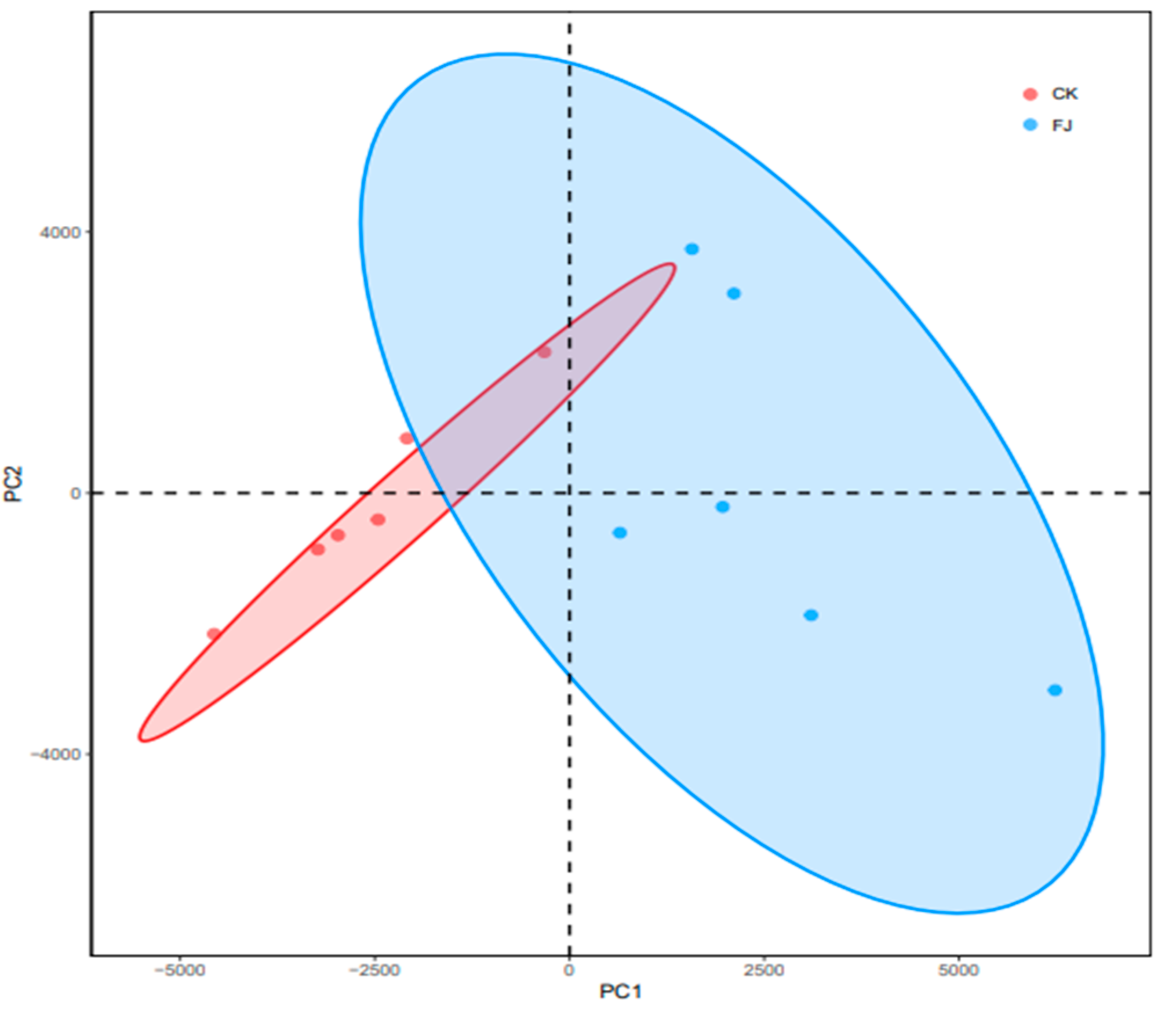
3.6.2. Beta Diversity Comparative Analysis–PCA Analysis of the Chicken Samples between the Groups
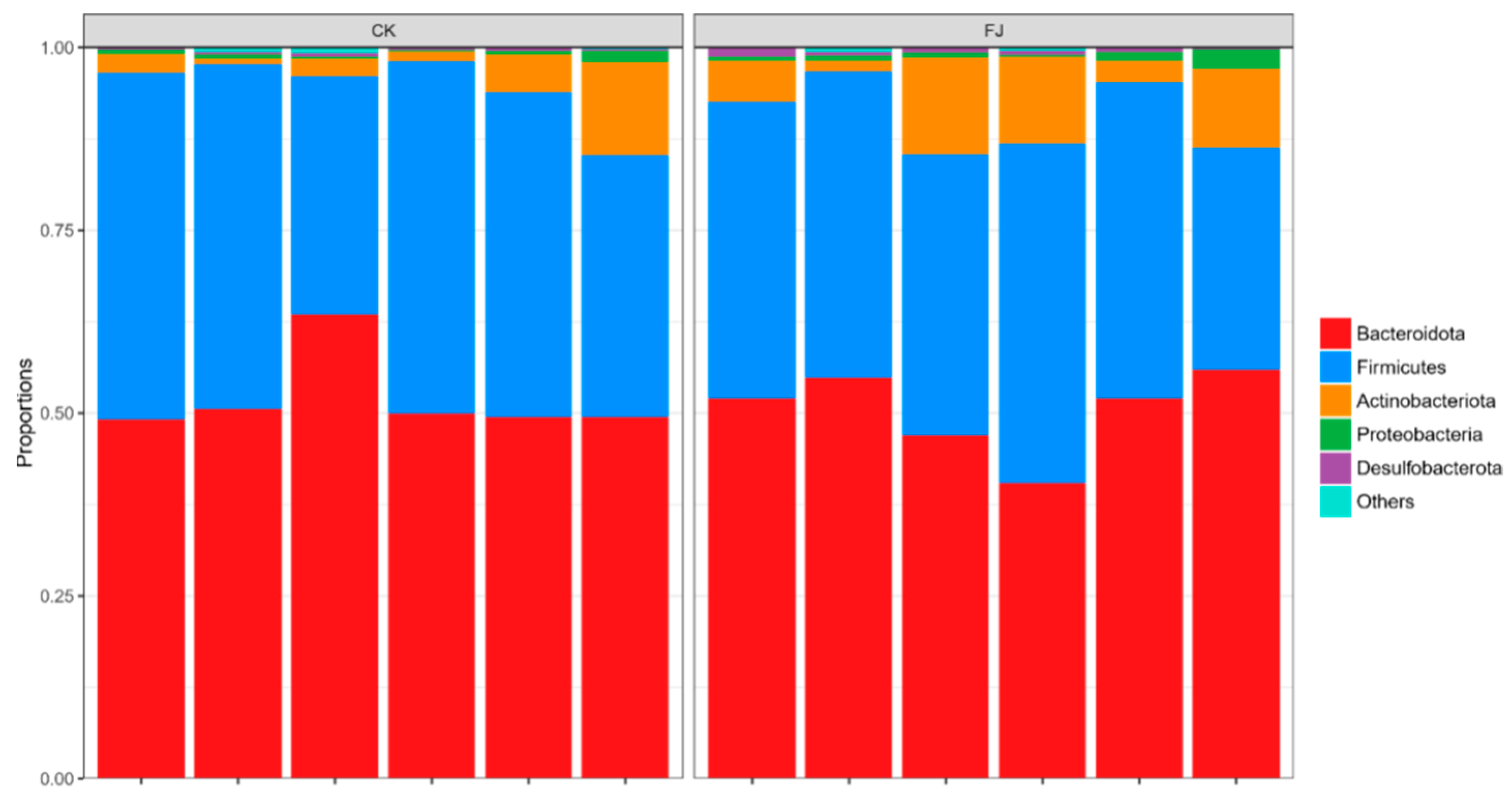
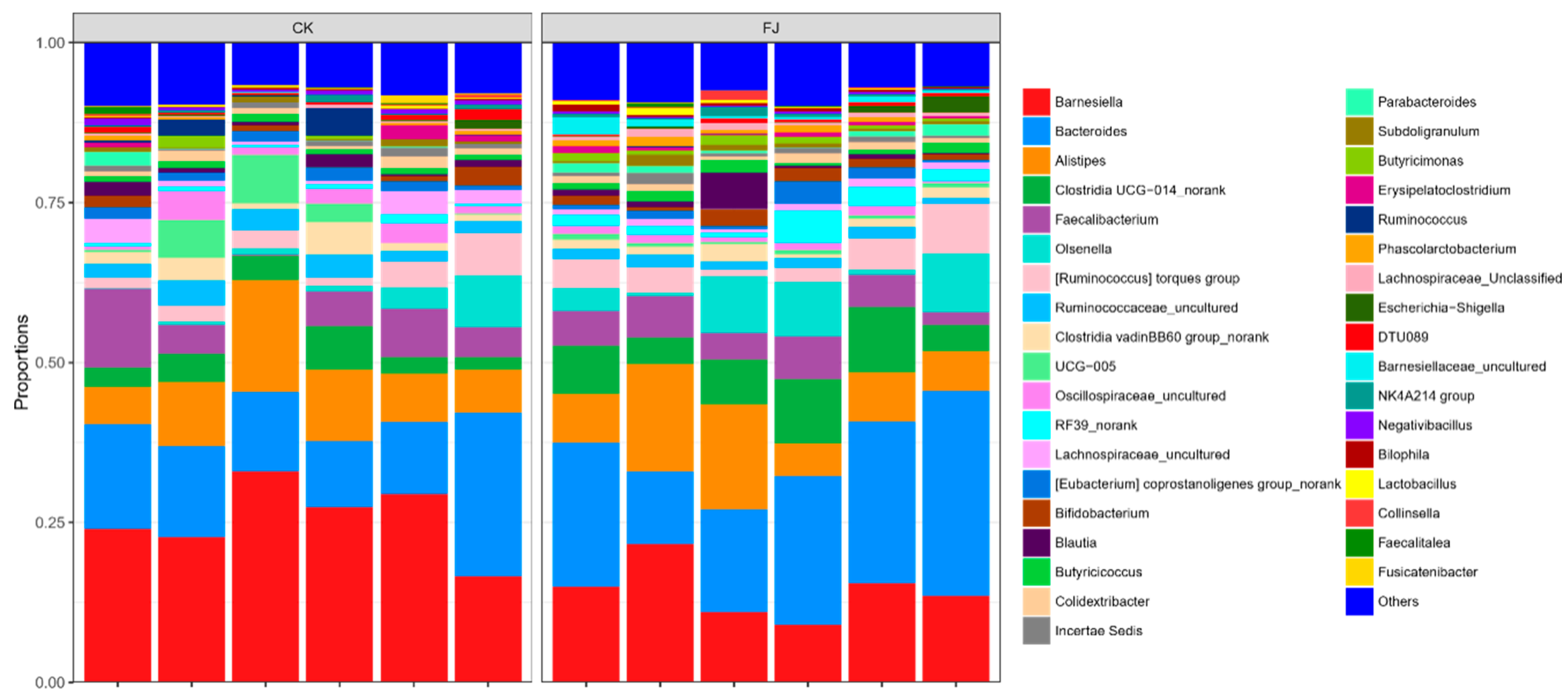
3.6.3. Relative Abundance of Cecal Microflora at Phylum and Genus Levels of Chickens
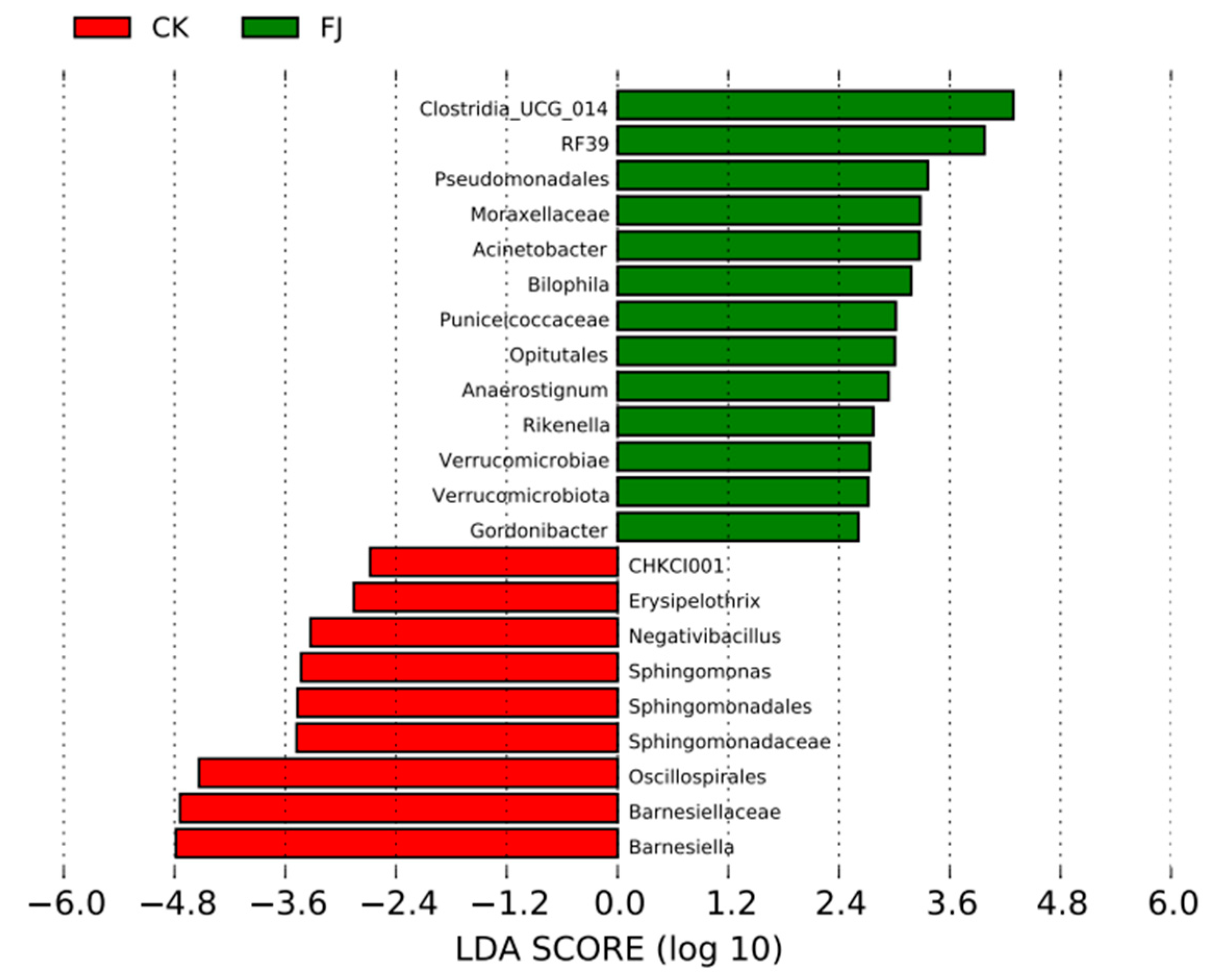
3.6.4. Lefse Difference Analysis of the Chicken Samples between the Groups
4. Discussion
4.1. Effects of Fermented Feed on the Growth Performance of Chickens
4.2. Effects of Fermented Feed on Immune Function in the Serum of Chickens
4.3. Effects of Fermented Feed on Antioxidational Ability of Chickens
4.4. Effects of Fermented Feed on Intestinal Digestive Enzyme Activity of Chickens
4.5. Effects of Fermented Feed on Intestinal Tissue Morphology of Chickens
4.6. Effects of Supplementing Fermented Feed on Microbial Diversity in the Cecum of Chickens
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wickramasuriya, S.S.; Park, I.; Lee, K.; Lee, Y.; Kim, W.H.; Nam, H.; Lillehoj, H.S. Role of physiology, immunity, microbiota, and infectious diseases in the gut health of poultry. Vaccines 2022, 10, 172. [Google Scholar] [CrossRef]
- Biesek, J.; Banaszak, M.; Kądziołka, K.; Wlaźlak, S.; Adamski, M. Growth of broiler chickens, and physical features of the digestive system, and leg bones after aluminosilicates used. Sci. Rep. 2022, 12, 20425. [Google Scholar] [CrossRef] [PubMed]
- Broom, L.J.; Kogut, M.H. The role of the gut microbiome in shaping the immune system of chickens. Vet. Immunol. Immunopathol. 2018, 204, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Shuaib, M.; Paneru, D.; Hafeez, A.; Tahir, M.; Kim, W.K. The chemical composition of soyhulls and their effect on amino acid and nutrient digestibility in laying hens during the peak of production. Animals 2023, 13, 2808. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Bai, Z.; Ma, W.; Guo, M.; Jiang, R.; Liu, J.; Oenema, O.; Velthof, G.L.; Whitmore, A.P.; Crawford, J.; et al. Exploring future food provision scenarios for China. Environ. Sci. Technol. 2019, 53, 1385–1393. [Google Scholar] [CrossRef]
- Yang, L.; Zeng, X.; Qiao, S. Advances in research on solid-state fermented feed and its utilization: The pioneer of private customization for intestinal microorganisms. Anim. Nutr. 2021, 7, 905–916. [Google Scholar] [CrossRef]
- Missotten, J.A.; Michiels, J.; Dierick, N.; Ovyn, A.; Akbarian, A.; De Smet, S. Effect of fermented moist feed on performance, gut bacteria and gut histo-morphology in broilers. Br. Poult. Sci. 2013, 54, 627–634. [Google Scholar] [CrossRef]
- Heres, L.; Engel, B.; van Knapen, F.; de Jong, M.C.; Wagenaar, J.A.; Urlings, H.A. Fermented liquid feed reduces susceptibility of broilers for Salmonella enteritidis. Poult. Sci. 2003, 82, 603–611. [Google Scholar] [CrossRef]
- Heres, L.; Engel, B.; Van Knapen, F.; Wagenaar, J.A.; Urlings, B.A. Effect of fermented feed on the susceptibility for Campylobacter jejuni colonisation in broiler chickens with and without concurrent inoculation of Salmonella enteritidis. Int. J. Food Microbiol. 2003, 87, 75–86. [Google Scholar] [CrossRef]
- Yan, J.; Zhou, B.; Xi, Y.; Huan, H.; Li, M.; Yu, J.; Zhu, H.; Dai, Z.; Ying, S.; Zhou, W.; et al. Fermented feed regulates growth performance and the cecal microbiota community in geese. Poult. Sci. 2019, 98, 4673–4684. [Google Scholar] [CrossRef]
- Wang, C.; Wei, S.; Xu, B.; Hao, L.; Su, W.; Jin, M.; Wang, Y. Bacillus subtilis and Enterococcus faecium co-fermented feed regulates lactating sow’s performance, immune status and gut microbiota. Microb. Biotechnol. 2021, 14, 614–627. [Google Scholar] [CrossRef]
- Zhu, Y.; Tao, Z.; Chen, X.; Xiao, J.; Zhang, Y.; Wang, Z. Effects of broussonetia papyrifera-fermented feed on production performance, egg quality, and caecal microbiota of laying hens during the late laying period. Ital. J. Anim. Sci. 2022, 21, 659–672. [Google Scholar] [CrossRef]
- Yeh, R.H.; Hsieh, C.W.; Chen, K.L. Screening lactic acid bacteria to manufacture two-stage fermented feed and pelleting to investigate the feeding effect on broilers. Poult. Sci. 2018, 97, 236–246. [Google Scholar] [CrossRef]
- Sadiq, F.A.; Yan, B.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Lactic acid bacteria as antifungal and anti-mycotoxigenic agents: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1403–1436. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Guo, B.; Wu, Z.; Wang, W.; Li, C.; Liu, G.; Cai, H. Effects of fermented soybean meal supplementation on the growth performance and cecal microbiota community of broiler chickens. Animals 2020, 10, 1098. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Chen, D.; Cai, H.; Chang, W.; Wang, Z.; Liu, G.; Deng, X.; Chen, Z. Effects of fermenting the plant fraction of a complete feed on the growth performance, nutrient utilization, antioxidant functions, meat quality, and intestinal microbiota of broilers. Animals 2022, 12, 2870. [Google Scholar] [CrossRef] [PubMed]
- Qiu, K.; He, W.; Zhang, H.; Wang, J.; Qi, G.; Guo, N.; Zhang, X.; Wu, S. Bio-fermented malic acid facilitates the production of high-quality chicken via enhancing muscle antioxidant capacity of broilers. Antioxidants 2022, 11, 2309. [Google Scholar] [CrossRef]
- Wu, Z.; Liu, J.; Chen, J.; Pirzado, S.A.; Li, Y.; Cai, H.; Liu, G. Effects of fermentation on standardized ileal digestibility of amino acids and apparent metabolizable energy in rapeseed meal fed to broiler chickens. Animals 2020, 10, 1774. [Google Scholar] [CrossRef]
- Li, P.; Ji, X.; Deng, X.; Hu, S.; Wang, J.; Ding, K.; Liu, N. Effect of rapeseed meal degraded by enzymolysis and fermentation on the growth performance, nutrient digestibility and health status of broilers. Arch. Anim. Nutr. 2022, 76, 221–232. [Google Scholar] [CrossRef]
- Zhang, Q.; Guo, M.; Li, F.; Qin, M.; Yang, Q.; Yu, H.; Xu, J.; Liu, Y.; Tong, T. Evaluation of fermented soybean meal to replace a portion fish meal on growth performance, antioxidant capacity, immunity, and mTOR signaling pathway of coho salmon (Oncorhynchus kisutch). Aquac. Nutr. 2023, 2023, 2558173. [Google Scholar] [CrossRef]
- Collins, N.; Belkaid, Y. Control of immunity via nutritional interventions. Immunity 2022, 55, 210–223. [Google Scholar] [CrossRef] [PubMed]
- Olukomaiya, O.O.; Adiamo, O.Q.; Fernando, W.C.; Mereddy, R.; Li, X.; Sultanbawa, Y. Effect of solid-state fermentation on proximate composition, anti-nutritional factor, microbiological and functional properties of lupin flour. Food Chem. 2020, 315, 126238. [Google Scholar] [CrossRef] [PubMed]
- Vlassa, M.; Filip, M.; Țăranu, I.; Marin, D.; Untea, A.E.; Ropotă, M.; Dragomir, C.; Sărăcilă, M. The yeast fermentation effect on content of bioactive, nutritional and anti-nutritional factors in rapeseed meal. Foods 2022, 11, 2972. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, A.; Sahu, N.P.; Deo, A.D.; Kumar, S. Solid state fermentation of de-oiled rice bran: Effect on in vitro protein digestibility, fatty acid profile and anti-nutritional factors. Food Res. Int. 2019, 119, 1–5. [Google Scholar] [CrossRef]
- Gustaw, K.; Niedźwiedź, I.; Rachwał, K.; Polak-Berecka, M. New insight into bacterial interaction with the matrix of plant-based fermented foods. Foods 2021, 10, 1603. [Google Scholar] [CrossRef]
- Zhu, X.; Tao, L.; Liu, H.; Yang, G. Effects of fermented feed on growth performance, immune organ indices, serum biochemical parameters, cecal odorous compound production, and the microbiota community in broilers. Poult. Sci. 2023, 102, 102629. [Google Scholar] [CrossRef] [PubMed]
- Omar, A.E.; Al-Khalaifah, H.S.; Ismail, T.A.; Abd El-Aziz, R.M.; El-Mandrawy, S.A.M.; Shalaby, S.I.; Ibrahim, D. Performance, serum biochemical and immunological parameters, and digestive enzyme and intestinal barrier-related gene expression of broiler chickens fed fermented fava bean by-products as a substitute for conventional feed. Front. Vet. Sci. 2021, 8, 696841. [Google Scholar] [CrossRef]
- Xie, P.J.; Huang, L.X.; Zhang, C.H.; Zhang, Y.L. Nutrient assessment of olive leaf residues processed by solid-state fermentation as an innovative feedstuff additive. J. Appl. Microbiol. 2016, 121, 28–40. [Google Scholar] [CrossRef]
- Wang, Z.; Shao, D.; Wu, S.; Song, Z.; Shi, S. Heat stress-induced intestinal barrier damage and dimethylglycine alleviates via improving the metabolism function of microbiota gut brain axis. Ecotoxicol. Environ. Saf. 2022, 244, 114053. [Google Scholar] [CrossRef]
- Mazzoni, M.; Zampiga, M.; Clavenzani, P.; Lattanzio, G.; Tagliavia, C.; Sirri, F. Effect of chronic heat stress on gastrointestinal histology and expression of feed intake-regulatory hormones in broiler chickens. Animal 2022, 16, 100600. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Y.; Sun, Q.; Knopf, J.; Herrmann, M.; Lin, L.; Jiang, J.; Shao, C.; Li, P.; He, X.; et al. Immune response in COVID-19: What is next? Cell Death Differ. 2022, 29, 1107–1122. [Google Scholar] [CrossRef]
- Varadé, J.; Magadán, S.; González-Fernández, Á. Human immunology and immunotherapy: Main achievements and challenges. Cell Mol. Immunol. 2021, 18, 805–828. [Google Scholar] [CrossRef] [PubMed]
- Megha, K.B.; Mohanan, P.V. Role of immunoglobulin and antibodies in disease management. Int. J. Biol. Macromol. 2021, 169, 28–38. [Google Scholar] [CrossRef]
- Manik, M.; Singh, R.K. Role of toll-like receptors in modulation of cytokine storm signaling in SARS-CoV-2-induced COVID-19. J. Med. Virol. 2022, 94, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, Y.; Hao, X.; Duan, Y.; Meng, Z.; An, X.; Qi, J. Dietary fermented soybean meal replacement alleviates diarrhea in weaned piglets challenged with enterotoxigenic Escherichia coli K88 by modulating inflammatory cytokine levels and cecal microbiota composition. BMC Vet. Res. 2020, 16, 245. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, D.; Długosz, E.; Zawistowska-Deniziak, A. Functional properties of food origin Lactobacillus in the gastrointestinal ecosystem—In vitro study. Probiotics Antimicrob. Proteins 2019, 11, 820–829. [Google Scholar] [CrossRef]
- Li, S.C.; Lin, H.P.; Chang, J.S.; Shih, C.K. Lactobacillus acidophilus-fermented germinated brown rice suppresses preneoplastic lesions of the colon in rats. Nutrients 2019, 11, 2718. [Google Scholar] [CrossRef]
- Shakya, S.; Danshiitsoodol, N.; Sugimoto, S.; Noda, M.; Sugiyama, M. Anti-oxidant and anti-inflammatory substance generated newly in Paeoniae Radix Alba extract fermented with plant-derived Lactobacillus brevis 174A. Antioxidants 2021, 10, 1071. [Google Scholar] [CrossRef]
- Jomova, K.; Raptova, R.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Reactive oxygen species, toxicity, oxidative stress, and antioxidants: Chronic diseases and aging. Arch. Toxicol. 2023, 97, 2499–2574. [Google Scholar] [CrossRef]
- Miazek, K.; Beton, K.; Śliwińska, A.; Brożek-Płuska, B. The effect of β-carotene, tocopherols and ascorbic acid as anti-oxidant molecules on human and animal in vitro/in vivo studies: A review of research design and analytical techniques used. Biomolecules 2022, 12, 1087. [Google Scholar] [CrossRef]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The chemistry of reactive oxygen species (ROS) revisited: Outlining their role in biological macromolecules (DNA, lipids and proteins) and induced pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef]
- Wang, Z.; He, Z.; Emara, A.M.; Gan, X.; Li, H. Effects of malondialdehyde as a byproduct of lipid oxidation on protein oxidation in rabbit meat. Food Chem. 2019, 288, 405–412. [Google Scholar] [CrossRef]
- Mas-Bargues, C.; Escrivá, C.; Dromant, M.; Borrás, C.; Viña, J. Lipid peroxidation as measured by chromatographic determination of malondialdehyde. Human plasma reference values in health and disease. Arch. Biochem. Biophys. 2021, 709, 108941. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Kidd, M.T. Antioxidant defence systems and oxidative stress in poultry biology: An update. Antioxidants 2019, 8, 235. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Zhang, B.; Li, J.; Zhu, L. Effects of fermented feed on growth performance, immune response, and antioxidant capacity in laying hen chicks and the underlying molecular mechanism involving nuclear factor-κB. Poult. Sci. 2020, 99, 2573–2580. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Sun, Z.Y.; Cao, F.L.; Ahmad, H.; Yang, X.H.; Zhao, L.G.; Wang, T. Effects of dietary supplementation with fermented ginkgo leaves on antioxidant capacity, intestinal morphology and microbial ecology in broiler chicks. Br. Poult. Sci. 2015, 56, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Zhang, J.F.; Wan, X.L.; Huang, Q.; He, J.T.; Zhang, X.H.; Zhao, L.G.; Zhang, L.L.; Wang, T. Effect of fermented Ginkgo biloba leaves on nutrient utilisation, intestinal digestive function and antioxidant capacity in broilers. Br. Poult. Sci. 2019, 60, 47–55. [Google Scholar] [CrossRef]
- Liu, X.; Ju, Y.; Huang, L.; Liu, M.; Bo, J.; Zhou, T.; Zhang, Y.; Liu, C.; Feng, M.; Zhang, S.; et al. Effects of a new fermented soya bean meal on growth performance, serum biochemistry profile, intestinal immune status and digestive enzyme activities in piglets. J. Anim. Physiol. Anim. Nutr. 2022, 106, 1046–1059. [Google Scholar] [CrossRef]
- Gallego-Lobillo, P.; Ferreira-Lazarte, A.; Hernández-Hernández, O.; Villamiel, M. In vitro digestion of polysaccharides: InfoGest protocol and use of small intestinal extract from rat. Food Res. Int. 2021, 140, 110054. [Google Scholar] [CrossRef]
- Patil, U.; Saetang, J.; Zhang, B.; Benjakul, S. Use of tuna visceral pepsin in combination with trypsin as digestion aid: Enhanced protein hydrolysis and bioavailability. Foods 2022, 12, 125. [Google Scholar] [CrossRef]
- Kozan, D.W.; Derrick, J.T.; Ludington, W.B.; Farber, S.A. From worms to humans: Understanding intestinal lipid metabolism via model organisms. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2023, 1868, 159290. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Liu, X.; Xu, Z.R.; Wang, Y.Z.; Liu, J.X. Effects of fermented soybean meal on digestive enzyme activities and intestinal morphology in broilers. Poult. Sci. 2007, 86, 1149–1154. [Google Scholar] [CrossRef]
- Sun, H.; Tang, J.W.; Yao, X.H.; Wu, Y.F.; Wang, X.; Feng, J. Effects of dietary inclusion of fermented cottonseed meal on growth, cecal microbial population, small intestinal morphology, and digestive enzyme activity of broilers. Trop. Anim. Health Prod. 2013, 45, 987–993. [Google Scholar] [CrossRef]
- DeSesso, J.M. Comparative anatomy, pre-and postnatal changes during the development and maturation of the small intestine: Life-stage influences on exposure. Birth Defects Res. 2022, 114, 449–466. [Google Scholar] [CrossRef] [PubMed]
- Dalia, A.M.; Loh, T.C.; Sazili, A.Q.; Samsudin, A.A. Influence of bacterial organic selenium on blood parameters, immune response, selenium retention and intestinal morphology of broiler chickens. BMC Vet. Res. 2020, 16, 365. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, J.; Mahfuz, S.; Long, S.; Wu, D.; Gao, J.; Piao, X. Supplementation of mixed organic acids improves growth performance, meat quality, gut morphology and volatile fatty acids of broiler chicken. Animals 2021, 11, 3020. [Google Scholar] [CrossRef]
- Dai, D.; Qiu, K.; Zhang, H.J.; Wu, S.G.; Han, Y.M.; Wu, Y.Y.; Qi, G.H.; Wang, J. Organic acids as alternatives for antibiotic growth promoters alter the intestinal structure and microbiota and improve the growth performance in broilers. Front. Microbiol. 2021, 11, 618144. [Google Scholar] [CrossRef]
- Farahat, M.; Ibrahim, D.; Kishawy, A.T.Y.; Abdallah, H.M.; Hernandez-Santana, A.; Attia, G. Effect of cereal type and plant extract addition on the growth performance, intestinal morphology, caecal microflora, and gut barriers gene expression of broiler chickens. Animal 2021, 15, 100056. [Google Scholar] [CrossRef]
- Peng, W.; Talpur, M.Z.; Zeng, Y.; Xie, P.; Li, J.; Wang, S.; Wang, L.; Zhu, X.; Gao, P.; Jiang, Q.; et al. Influence of fermented feed additive on gut morphology, immune status, and microbiota in broilers. BMC Vet. Res. 2022, 18, 218. [Google Scholar] [CrossRef]
- Xiang, Y.; Wen, H.; Yu, Y.; Li, M.; Fu, X.; Huang, S. Gut-on-chip: Recreating human intestine in vitro. J. Tissue Eng. 2020, 11, 2041731420965318. [Google Scholar] [CrossRef]
- El-Sayed, A.; Aleya, L.; Kamel, M. The link among microbiota, epigenetics, and disease development. Environ. Sci. Pollut. Res. 2021, 28, 28926–28964. [Google Scholar] [CrossRef]
- Ding, S.; Yan, W.; Ma, Y.; Fang, J. The impact of probiotics on gut health via alternation of immune status of monogastric animals. Anim. Nutr. 2021, 7, 24–30. [Google Scholar] [CrossRef]
- Lv, J.; Guo, L.; Chen, B.; Hao, K.; Ma, H.; Liu, Y.; Min, Y. Effects of different probiotic fermented feeds on production performance and intestinal health of laying hens. Poult. Sci. 2022, 101, 101570. [Google Scholar] [CrossRef]
- Segura-Wang, M.; Grabner, N.; Koestelbauer, A.; Klose, V.; Ghanbari, M. Genome-resolved metagenomics of the chicken gut microbiome. Front. Microbiol. 2021, 12, 726923. [Google Scholar] [CrossRef]
- Xiao, S.S.; Mi, J.D.; Mei, L.; Liang, J.; Feng, K.X.; Wu, Y.B.; Liao, X.D.; Wang, Y. Microbial diversity and community variation in the intestines of layer chickens. Animals 2021, 11, 840. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, S.; Nie, Q.; He, H.; Tan, H.; Geng, F.; Ji, H.; Hu, J.; Nie, S. Gut firmicutes: Relationship with dietary fiber and role in host homeostasis. Crit. Rev. Food Sci. Nutr. 2022, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Chen, N.; Liu, T.; Feng, C. Effects of adding different carbon sources on the microbial behavior of sulfate-reducing bacteria in sulfate-containing wastewater. J. Clean. Prod. 2023, 392, 136332. [Google Scholar] [CrossRef]
- Zafar, H.; Saier, M.H., Jr. Gut Bacteroides species in health and disease. Gut Microbes 2021, 13, 1848158. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P. Glycan utilisation system in Bacteroides and Bifidobacteria and their roles in gut stability and health. Appl. Microbiol. Biotechnol. 2019, 103, 7287–7315. [Google Scholar] [CrossRef]
- Guo, X.; Okpara, E.S.; Hu, W.; Yan, C.; Wang, Y.; Liang, Q.; Chiang, J.Y.L.; Han, S. Interactive relationships between intestinal flora and bile acids. Int. J. Mol. Sci. 2022, 23, 8343. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Basal Diets | Contents | Fermented Feed | Contents |
|---|---|---|---|
| Ingredients | Ingredients | ||
| Corn, % | 21.30 | Corn,% | 20.59 |
| Wheat bran, % | 43.64 | Beancurd residue, % | 30.12 |
| Soybean meal, % | 19.26 | Beer grains, % | 20.26 |
| Extruded soybeans, % | 3.00 | Soybean meal, % | 11.79 |
| Sunflower seed meal, % | 2.00 | Extruded soybeans, % | 3.50 |
| Peanut meal, % | 3.00 | Sunflower seed meal, % | 2.56 |
| Corn gluten meal, % | 2.56 | Corn gluten meal, % | 3.00 |
| Lard oil, % | 1.07 | sunflower seed oil,% | 4.01 |
| Calcium hydrogen phosphate, % | 1.09 | Calcium hydrogen phosphate, % | 1.09 |
| Limestone, % | 1.34 | Limestone, % | 1.34 |
| Baking soda, % | 0.20 | Baking soda, % | 0.20 |
| Salt, % | 0.25 | Salt, % | 0.25 |
| Methionine (95%), % | 0.23 | Methionine (95%), % | 0.23 |
| Threonine (95%), % | 0.15 | Threonine (95%), % | 0.15 |
| Lysine sulphate, % | 0.69 | Lysine sulphate, % | 0.69 |
| Choline, % | 0.08 | Choline, % | 0.08 |
| Premix 2, % | 0.14 | Premix 2, % | 0.14 |
| Total | 100.00 | Total | 100.00 |
| Nutrient levels, on air-dry basis | Nutrient levels, on air-dry basis | ||
| Metabolic energy 3, ME, MJ/kg | 12.10 | Metabolic energy 3, ME, MJ/kg | 12.11 |
| Crude protein 4, CP, % | 21.00 | Crude protein 4, CP, % | 20.93 |
| Crude fiber 4, CF, % | 2.96 | Crude fiber 4, CF, % | 2.98 |
| Calcium 4, Ca, % | 0.88 | Calcium 4, Ca, % | 0.89 |
| Phosphorus 4, P, % | 0.60 | Phosphorus 4, P, % | 0.59 |
| Lysine 4, Lys, % | 1.15 | Lysine 4, Lys, % | 1.14 |
| Methionine 4, Met, % | 0.52 | Methionine 4, Met, % | 0.53 |
| Items | Groups | p-Value | |
|---|---|---|---|
| CK | FJ | ||
| IgA, g/L | 1.91 ± 0.04 b | 2.69 ± 0.05 a | <0.001 |
| IgG, g/L | 4.76 ± 0.10 b | 5.32 ± 0.10 a | 0.002 |
| IgM, g/L | 1.47 ± 0.07 b | 1.91 ± 0.08 a | 0.001 |
| IL-1β, pg/mL | 35.07 ± 0.49 a | 19.18 ± 0.43 b | <0.001 |
| IL-6, pg/mL | 218 ± 6.21 a | 120 ± 4.34 b | <0.001 |
| Items | Groups | p-Value | |
|---|---|---|---|
| CK | FJ | ||
| Duodenum | |||
| Amylase, U/mg·protein | 5.68 ± 1.2 | 4.77 ± 1.04 | 0.580 |
| Trypsin, U/mg·protein | 1536 ± 135.03 b | 2967 ± 345.6 a | 0.003 |
| Lipase, U/g·protein | 7.66 ± 1.38 | 8.55 ± 1.59 | 0.620 |
| Jejunum | |||
| Amylase, U/mg·protein | 3.09 ± 0.88 | 4.39 ± 0.82 | 0.305 |
| Trypsin, U/mg·protein | 25977 ± 336.71 | 26507 ± 324.52 | 0.121 |
| Lipase, U/g·protein | 6.29 ± 1.25 | 25.82 ± 9.92 | 0.110 |
| Ileum | |||
| Amylase, U/mg·protein | 2.63 ± 0.43 | 4.6 ± 1.21 | 0.190 |
| Trypsin, U/mg·protein | 25077 ± 175.96 | 25957 ± 347.06 | 0.238 |
| Lipase, U/g·protein | 21.85 ± 7.12 | 22.42 ± 7.69 | 0.568 |
| Items | Groups | p-Value | |
|---|---|---|---|
| CK | FJ | ||
| Duodenum | |||
| Villus height, μm | 954 ± 30.51 | 1026 ± 75.44 | 0.382 |
| Crypt depth, μm | 169 ± 15.58 | 141 ± 10.35 | 0.141 |
| Villus height/Crypt depth, μm/μm | 6.52 ± 0.75 | 8.07 ± 0.91 | 0.197 |
| Jejunum | |||
| Villus height, μm | 794 ± 32.89 | 822 ± 43.64 | 0.620 |
| Crypt depth, μm | 121 ± 10.86 | 1162 ± 8.57 | 0.279 |
| Villus height/Crypt depth, μm/μm | 6.92 ± 0.7 | 7.08 ± 0.7 | 0.709 |
| Ileum | |||
| Villus height, μm | 769 ± 18.57 a | 527 ± 31.56 b | 0.000 |
| Crypt depth, μm | 162 ± 13.64 | 132 ± 10.08 | 0.086 |
| Villus height/Crypt depth, μm/μm | 4.78 ± 0.40 | 4.23 ± 0.35 | 0.312 |
| Items | Groups | p-Value | |
|---|---|---|---|
| CK | FJ | ||
| Chao1 index | 997 ± 32.71 | 10387 ± 39.96 | 0.437 |
| Richness index | 8397 ± 34.33 | 853 ± 29.86 | 0.767 |
| Shannon index | 5.94 ± 0.05 b | 6.32 ± 0.1 a | 0.040 |
| Simpson index | 0.08 ± 0.01 | 0.05 ± 0.01 | 0.098 |
| ACE index | 1002 ± 34.15 | 1045 ± 34.74 | 0.400 |
| Evenness index | 0.61 ± 0.004 | 0.64 ± 0.015 | 0.119 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, F.; Wu, H.; Xie, J.; Zeng, T.; Hao, L.; Xu, W.; Lu, L. The Effects of Fermented Feed on the Growth Performance, Antioxidant Activity, Immune Function, Intestinal Digestive Enzyme Activity, Morphology, and Microflora of Yellow-Feather Chickens. Animals 2023, 13, 3545. https://doi.org/10.3390/ani13223545
Xu F, Wu H, Xie J, Zeng T, Hao L, Xu W, Lu L. The Effects of Fermented Feed on the Growth Performance, Antioxidant Activity, Immune Function, Intestinal Digestive Enzyme Activity, Morphology, and Microflora of Yellow-Feather Chickens. Animals. 2023; 13(22):3545. https://doi.org/10.3390/ani13223545
Chicago/Turabian StyleXu, Fei, Hongzhi Wu, Jiajun Xie, Tao Zeng, Lijian Hao, Wenwu Xu, and Lizhi Lu. 2023. "The Effects of Fermented Feed on the Growth Performance, Antioxidant Activity, Immune Function, Intestinal Digestive Enzyme Activity, Morphology, and Microflora of Yellow-Feather Chickens" Animals 13, no. 22: 3545. https://doi.org/10.3390/ani13223545
APA StyleXu, F., Wu, H., Xie, J., Zeng, T., Hao, L., Xu, W., & Lu, L. (2023). The Effects of Fermented Feed on the Growth Performance, Antioxidant Activity, Immune Function, Intestinal Digestive Enzyme Activity, Morphology, and Microflora of Yellow-Feather Chickens. Animals, 13(22), 3545. https://doi.org/10.3390/ani13223545