
Bridging Evolutionary History and Conservation of New World Vultures

 ,
,  ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Methods
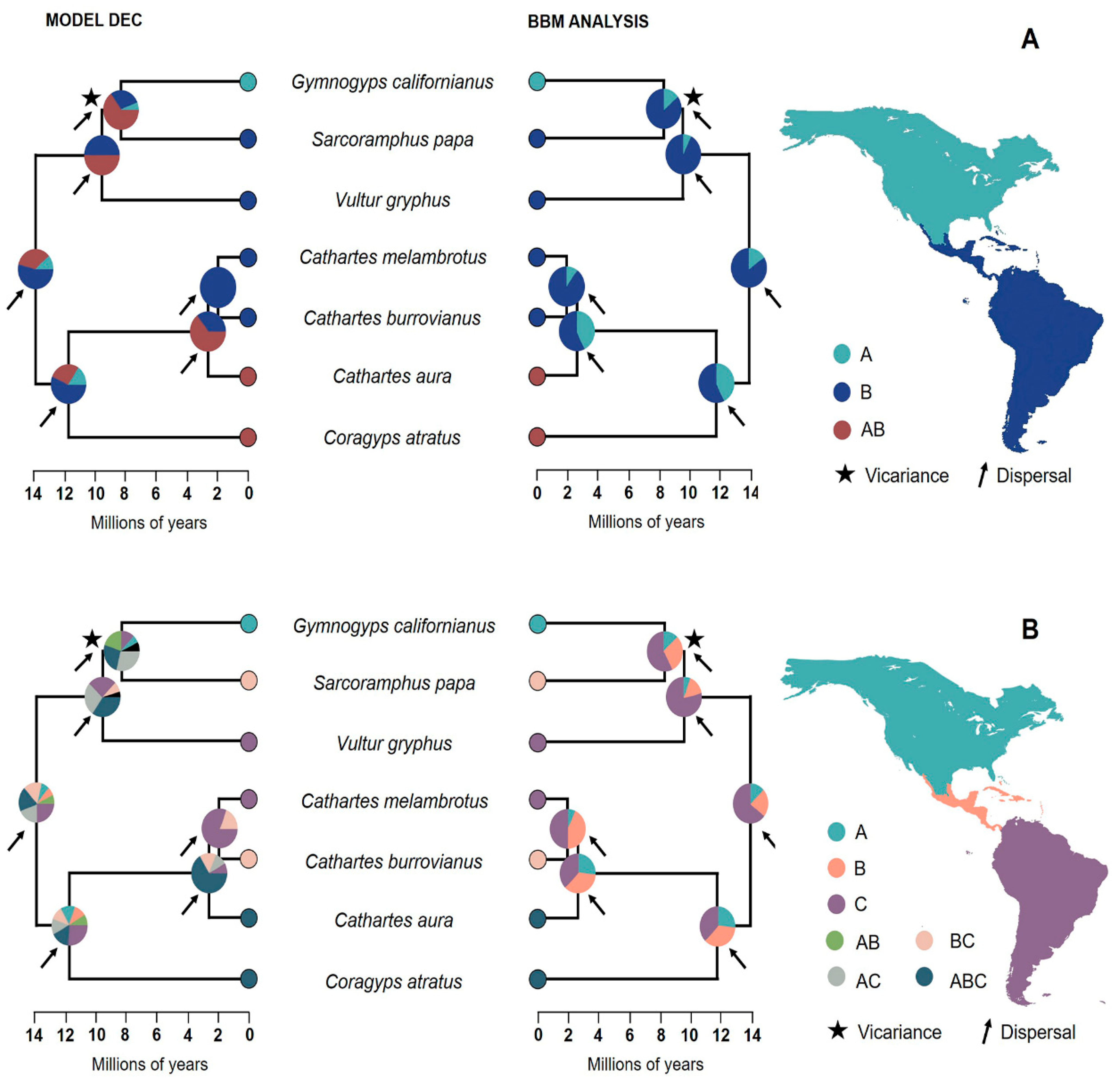
2.1. Biogeography
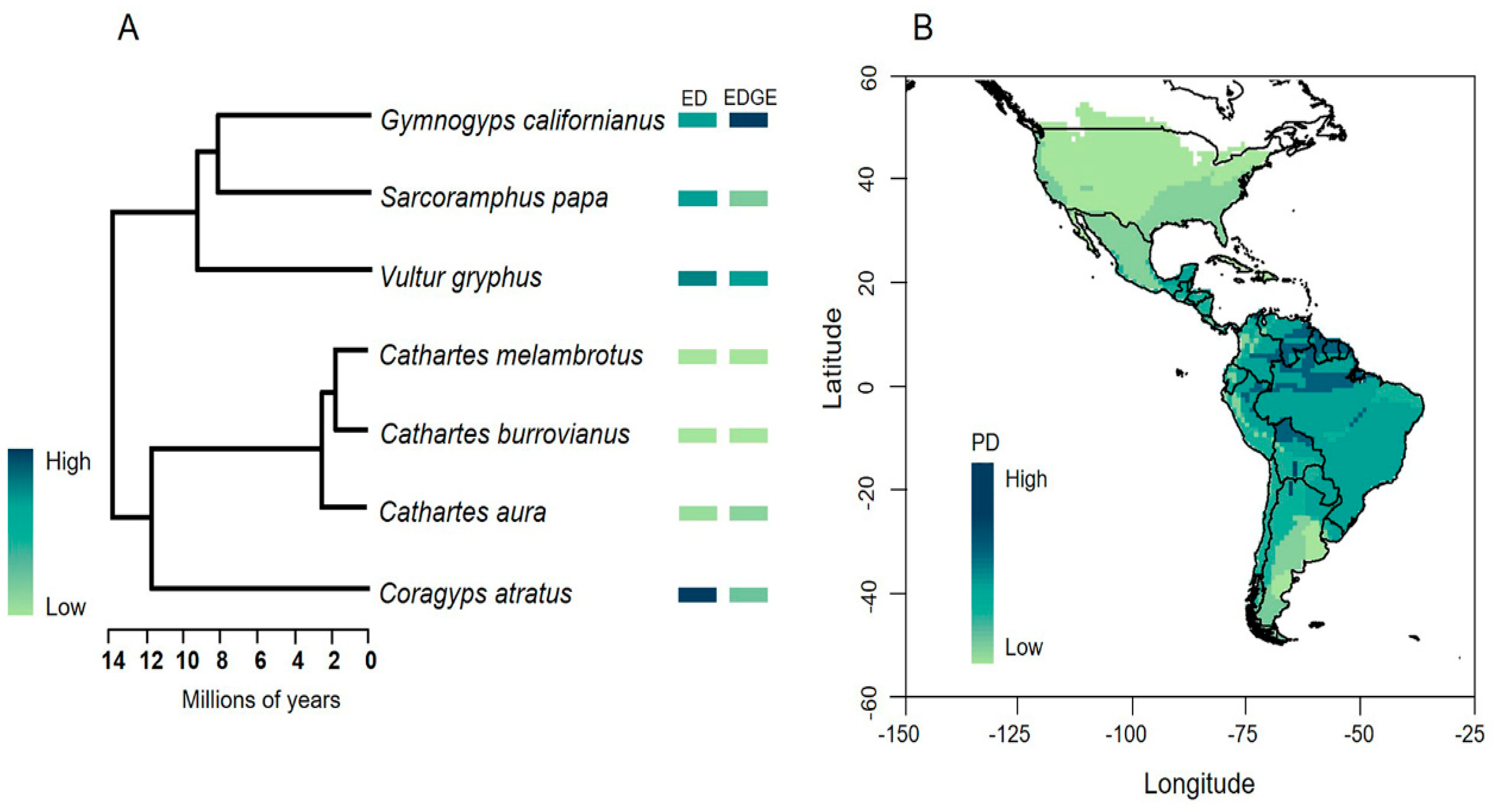
2.2. Phylogenetic Diversity, Evolutionary Distinctiveness and EDGE Species
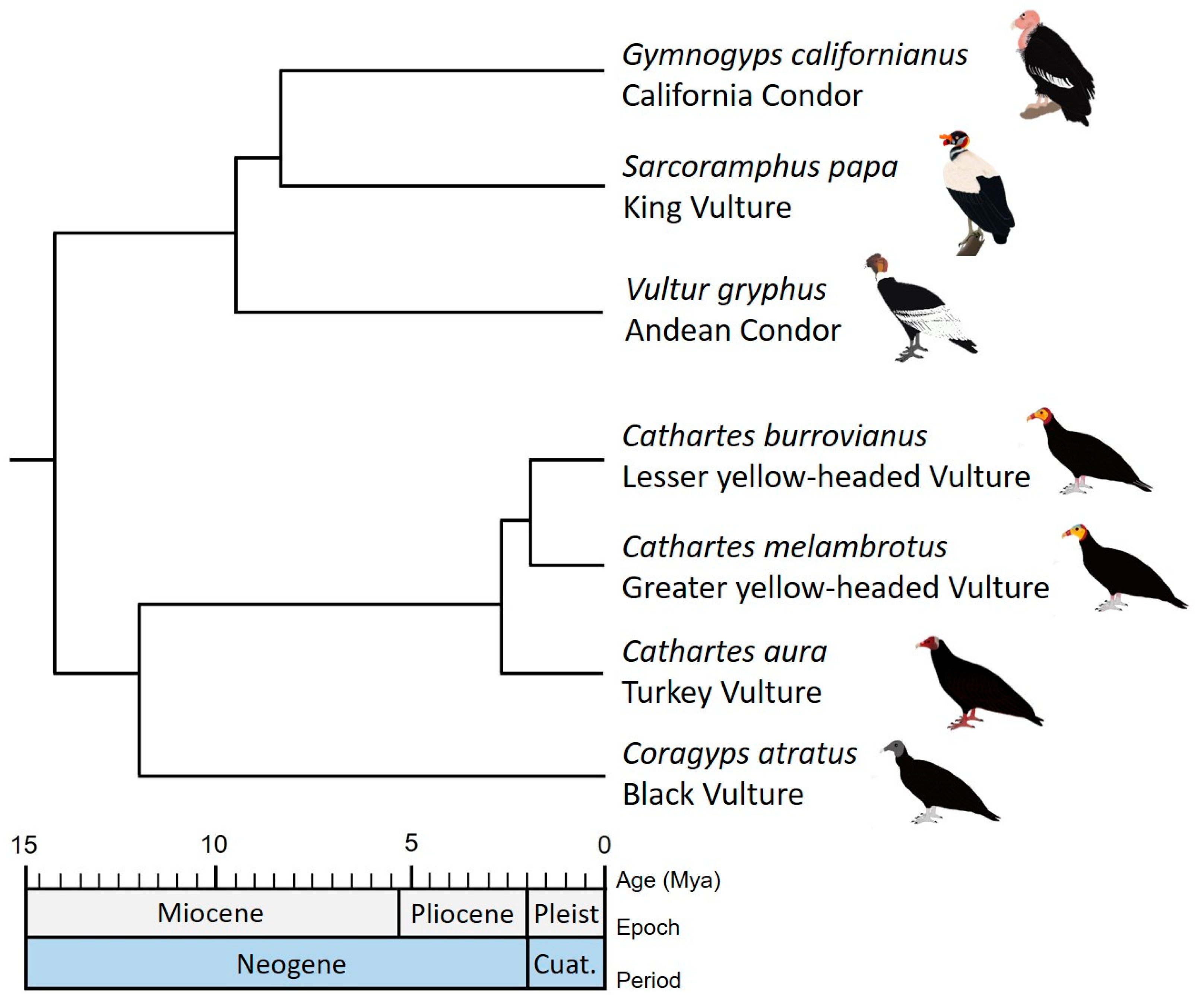
3. Results
3.1. Biogeography
3.2. Phylogenetic Diversity, ED and EDGE Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campbell, M.O. Vultures: Their Evolution, Ecology and Conservation; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2015. [Google Scholar]
- Stucchi, M.; Emslie, S.D.; Varas-Malca, R.M.; Urbina-Schmitt, M. A new late Miocene condor (Aves, Cathartidae) from Peru and the origin of South American condors. J. Vertebr. Paleontol. 2015, 35, e972507. [Google Scholar] [CrossRef]
- Remsen, J.V., Jr.; Areta, J.I.; Bonaccorso, E.; Claramunt, S.; Del-Rio, G.; Jaramillo, A.; Lane, D.F.; Robbins, M.B.; Stiles, F.G.; Zimmer, K.J. A Classification of the Bird Species of South America; Museum of Natural Science, Louisiana State University: Baton Rouge, USA, 2020; Available online: http://www.museum.lsu.edu/~Remsen/SACCBaseline.htm (accessed on 10 October 2023).
- Avise, J.C.; Nelson, W.S.; Sibley, C.G. DNA sequence support for a close phylogenetic relationship between some storks and New World vultures. Proc. Natl. Acad. Sci. USA 1994, 91, 5173–5177. [Google Scholar] [CrossRef] [PubMed]
- Sibley, C.G.; Ahlquist, J.E.; Monroe Jr, B.L. A classification of the living birds of the world based on DNA-DNA hybridization studies. Auk 1988, 105, 409–423. [Google Scholar] [CrossRef]
- Sibley, C.G.; Ahlquist, J.E. Phylogeny and Classification of Birds; Yale University Press: New Haven, CT, USA, 1990. [Google Scholar]
- Ericson, P.G. Evolution of terrestrial birds in three continents: Biogeography and parallel radiations. J. Biogeogr. 2012, 39, 813–824. [Google Scholar] [CrossRef]
- Prum, R.O.; Berv, J.S.; Dornburg, A.; Field, D.J.; Townsend, J.P.; Lemmon, E.M.; Lemmon, A.R. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature 2015, 526, 569–573. [Google Scholar] [CrossRef]
- Jetz, W.; Thomas, G.H.; Joy, J.B.; Redding, D.W.; Hartmann, K.; Mooers, A.O. The global diversity of birds in space and time. Nature 2012, 491, 444–448. [Google Scholar] [CrossRef]
- Seibold, I.; Helbig, A.J. Evolutionary history of New and Old World vultures inferred from nucleotide sequences of the mitochondrial cytochrome b gene. Philos. Trans. R. Soc. London Ser. B Biol. Sci. 1995, 350, 163–178. [Google Scholar]
- Wink, M.; Sauer-Gürth, H. Phylogenetic relationships in diurnal raptors based on nucleotide sequences of mitochondrial and nuclear marker genes. Raptors Worldw. 2004, 483–498. [Google Scholar]
- Johnson, J.A.; Brown, J.; Fuchs, J.; Mindell, D.P. Multi-locus phylogenetic inference among New World Vultures (Aves: Cathartidae). Mol. Phylogenet. Evol. 2016, 105, 193–199. [Google Scholar] [CrossRef]
- Ferguson-Lees, J.; Christie, D. Raptors of the World; Houghton Mifflin Company: Boston, MA, USA, 2001. [Google Scholar]
- Sánchez-Soto, S.; Gómez-Martínez, U.N. Segundo registro de Sarcoramphus papa (Cathartidae) en el estado de Tabasco, México. Boletín SAO 2014, 23, 6–10. [Google Scholar]
- Lambertucci, S.A. Biología y conservación del Cóndor Andino (Vultur gryphus) en Argentina. El Hornero 2007, 22, 149–158. [Google Scholar] [CrossRef]
- de la Cruz, E.; Peters, E. La reintroducción del cóndor de California en la Sierra de San Pedro Mártir, Baja California. Gac. Ecológica 2007, 82, 55–67. [Google Scholar]
- Spina, M.A. Levantamento Parasitológico em Coragyps atratus Bechstein, 1793 (Cathartiformes, Cathartidae) de Vida Livre no Parque Estadual das Fontes do Ipiranga e Análise de Potencial Risco Biológico Para a Saúde Animal e Ambiental. Master’s Thesis, Universidade Federal de Sao Carlos, São Paulo, Brasil, 2019. [Google Scholar]
- Ogada, D.L.; Keesing, F.; Virani, M.Z. Dropping dead: Causes and consequences of vulture population declines worldwide. Ann. N.Y. Acad. Sci. 2012, 1249, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Buechley, E.R.; Şekercioğlu, C.H. The avian scavenger crisis: Looming extinctions, trophic cascades, and loss of critical ecosystem functions. Biol. Conserv. 2016, 198, 220–228. [Google Scholar] [CrossRef]
- Plaza, P.I.; Lambertucci, S.A. What do we know about lead contamination in wild vultures and condors? A review of decades of research. Sci. Total Environ. 2019, 654, 409–417. [Google Scholar] [CrossRef]
- Restrepo-Cardona, J.S.; Parrado-Vargas, M.A.; Vargas, F.H.; Kohn, S.; Sáenz-Jiménez, F.; Potaufeu, Y.; Narváez, F. Anthropogenic threats to the Vulnerable Andean Condor in northern South America. PLoS ONE 2022, 17, e0278331. [Google Scholar] [CrossRef]
- Campbell, M.O. Biological Conservation in the 21st Century: A Conservation Biology of Large Wildlife; Nova Science Publisher: New York, NY, USA, 2017. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Version 2020-2. 2020. Available online: https://www.iucnredlist.org/ (accessed on 10 October 2023).
- BirdLife International. Cathartes aura. In IUCN Red List of Threatened Species; Version 2012.1; BirdLife International: Cambridge, UK, 2023; Available online: https://www.iucnredlist.org/species/22697627/131941613 (accessed on 10 October 2023).
- BirdLife International. Cathartes melambrotus. In IUCN Red List of Threatened Species; Version 2012.1; BirdLife International: Cambridge, UK, 2023; Available online: https://www.iucnredlist.org/species/22697633/93626176 (accessed on 10 October 2023).
- BirdLife International. Cathartes burrovianus. In IUCN Red List of Threatened Species; Version 2012.1; BirdLife International: Cambridge, UK, 2023; Available online: https://www.iucnredlist.org/species/22697630/163511443 (accessed on 10 October 2023).
- BirdLife International. Coragyps atratus. In IUCN Red List of Threatened Species; Version 2012.1; BirdLife International: Cambridge, UK, 2023; Available online: https://www.iucnredlist.org/species/22697624/93624950 (accessed on 10 October 2023).
- BirdLife International. Sarcoramphus papa. In IUCN Red List of Threatened Species; Version 2012.1; BirdLife International: Cambridge, UK, 2023; Available online: https://www.iucnredlist.org/species/22697645/93627003 (accessed on 10 October 2023).
- Monsalvo, J.A.; Silva, M.A.; Heming, N.M.; Marini, M.A. Geographical variation and current knowledge on breeding traits of vultures in the neotropics. Ornithol. Res. 2020, 28, 13–37. [Google Scholar] [CrossRef]
- Santangeli, A.; Buechley, E.R.; Mammola, S.; Lambertucci, S.A. Priorities for research and action to prevent a New World vulture crisis. Biol. Conserv. 2022, 270, 109563. [Google Scholar] [CrossRef]
- Wilson, E.E.; Wolkovich, E.M. Scavenging: How carnivores and carrion structure communities. Trends Ecol. Evol. 2011, 26, 129–135. [Google Scholar] [CrossRef]
- Benbow, M.E.; Tomberlin, J.K.; Tarone, A.M. Carrion Ecology, Evolution, and Their Applications, 1st ed.; CRC press (Taylor & Francis Group): Boca Raton, FL, USA, 2015. [Google Scholar]
- Plaza, P.I.; Blanco, B.; Lambertucci, S.A. Implications of bacterial, viral and mycotic microorganisms in vultures for wildlife conservation, ecosystem services and public health. Ibis 2020, 162, 1109–1124. [Google Scholar] [CrossRef]
- Markandya, A.; Taylor, T.; Longo, A.; Murty, M.N.; Murty, S.; Dhavala, K. Counting the cost of vulture decline-an appraisal of the human health and other benefits of vultures in India. Ecol. Econ. 2008, 67, 194–204. [Google Scholar] [CrossRef]
- Michel, N.L.; Whelan, C.J.; Verutes, G.M. Ecosystem services provided by Neotropical birds. Condor 2020, 122, duaa022. [Google Scholar] [CrossRef]
- McClure, C.J.; Westrip, J.R.; Johnson, J.A.; Schulwitz, S.E.; Virani, M.Z.; Davies, R.; Symes, A.; Wheatley, H.; Thorstrom, R.; Amar, A.; et al. State of the world’s raptors: Distributions, threats, and conservation recommendations. Biol. Conserv. 2018, 227, 390–402. [Google Scholar] [CrossRef]
- Soltis, P.S.; Soltis, D.E. Molecular systematics: Assembling and using the Tree of Life. Taxon 2001, 50, 663–677. [Google Scholar] [CrossRef]
- Bininda-Emonds, O.R. Phylogenetic Supertrees: Combining Information to Reveal the Tree of Life; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; Volume 4. [Google Scholar]
- Delsuc, F.; Brinkmann, H.; Philippe, H. Phylogenomics and the reconstruction of the tree of life. Nat. Rev. Genet. 2005, 6, 361–375. [Google Scholar] [CrossRef]
- Ciccarelli, F.D.; Doerks, T.; Von Mering, C.; Creevey, C.J.; Snel, B.; Bork, P. Toward automatic reconstruction of a highly resolved tree of life. Science 2006, 311, 1283–1287. [Google Scholar] [CrossRef]
- Maddison, D.R.; Schulz, K.S.; Maddison, W.P. The tree of life web project. Zootaxa 2007, 1668, 19–40. [Google Scholar] [CrossRef]
- Isaac, N.J.B.; Turvey, S.T.; Collen, B.; Waterman, C.; Baillie, J.E.M. Mammals on the EDGE: Conservation priorities based on threat and phylogeny. PLoS ONE 2007, 2, e296. [Google Scholar] [CrossRef]
- Gumbs, R.; Gray, C.L.; Wearn, O.R.; Owen, N.R. Tetrapods on the EDGE: Overcoming data limitations to identify phylogenetic conservation priorities. PLoS ONE 2018, 13, e0194680. [Google Scholar] [CrossRef]
- McClure, C.J.; Berkunsky, I.; Buechley, E.R.; Dunn, L.; Johnson, J.; McCabe, J.; Oppel, S.; Rolek, B.W.; Sutton, L.J.; Gumbs, R. Conserving the evolutionary history of birds. Conserv. Biol. 2023. [Google Scholar] [CrossRef]
- Collen, B.; Turvey, S.T.; Waterman, C.; Meredith, H.M.; Kuhn, T.S.; Baillie, J.E.; Isaac, N.J. Investing in evolutionary history: Implementing a phylogenetic approach for mammal conservation. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2611–2622. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, D.S.; Cadotte, M.W.; MacDonald, A.A.M.; Marushia, R.G.; Mirotchnick, N. Phylogenetic diversity and the functioning of ecosystems. Ecol. Lett. 2012, 15, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Lean, C.; Maclaurin, J. The value of phylogenetic diversity. In Biodiversity Conservation and Phylogenetic Systematic; Pellens, R., Grandcolas, P., Eds.; Springer Nature: Cham, Switzerland, 2016. [Google Scholar]
- Yu, Y.; Harris, J.; Blair, C.; He, X. RASP (Reconstruct Ancestral State in Phylogenies): A tool for historical biogeography. Mol. Phylogenet. Evol. 2015, 87, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Matzke, N.J. Probabilistic historical biogeography: New models for founder-event speciation, imperfect detection, and fossils allow improved accuracy and model-testing. Front. Biogeogr. 2013, 5, 242–248. [Google Scholar]
- Pulgarín-R, P.C.; Smith, B.T.; Bryson, R.W.; Spellman, G.M.; Klicka, J. Multilocus phylogeny and biogeography of the New World Pheucticus grosbeaks (Aves: Cardinalidae). Mol. Phylogenet. Evol. 2013, 69, 1222–1227. [Google Scholar] [CrossRef]
- Fuchs, J.; Johnson, J.A.; Mindell, D.P. Rapid diversification of falcons (Aves: Falconidae) due to expansion of open habitats in the Late Miocene. Mol. Phylogenet. Evol. 2015, 82, 166–182. [Google Scholar] [CrossRef]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 463–1464. [Google Scholar] [CrossRef]
- Thuiller, W.; Maiorano, L.; Mazel, F.; Guilhaumon, F.; Ficetola, G.F.; Lavergne, S.; Renaud, J.; Roquet, C.; Mouillot, D. Conserving the functional and phylogenetic trees of life of European tetrapods. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140005. [Google Scholar] [CrossRef]
- Daru, B.H.; Karunarathne, P.; Schliep, K. Phyloregion: R package for biogeographic regionalization and spatial conservation. Methods Ecol. Evol. 2021, 11, 1483–1491. [Google Scholar] [CrossRef]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Mayr, E. History of the North American bird fauna. Wilson Bull. 1946, 58, 3–41. [Google Scholar]
- Wetmore, A. Fossil birds from the Oligocene of Colorado. Colo. Mus. Naturla Hist. 1927, 7, 1–13. [Google Scholar]
- Cracraft, J.; Rich, P.V. The systematics and evolution of the Cathartidae in the Old World Tertiary. Condor 1972, 74, 272–283. [Google Scholar] [CrossRef][Green Version]
- Alvarenga, H.F. Notas sobre os Cathartidae (Aves) e descrição de um novo gênero do Cenozóico brasileiro. Anais Academia Brasileira Ciências 1985, 57, 349–357. [Google Scholar]
- Emslie, S.D. The fossil history and phylogenetic relationships of condors (Ciconiiformes: Vulturidae) in the New World. J. Vertebr. Paleontol. 1988, 8, 212–228. [Google Scholar] [CrossRef]
- Tonni, E.P.; Noriega, J.I. Los cóndores (Ciconiiformes, Vulturidae) de la Región Pampeana de la Argentina durante el Cenozoico Tardío: Distribución, interacciones y extinciones. Ameghiniana 1998, 35, 141–150. [Google Scholar]
- Blandin, P.; Purser, B. Evolution and diversification of neotropical butterflies: Insights from the biogeography and phylogeny of the genus Morpho Fabricius, 1807 (Nymphalidae: Morphinae), with a review of the geodynamics of South America. Trop. Lepid. Res. 2013, 23, 62–85. [Google Scholar]
- Suárez, W. Contribución al conocimiento del estatus genérico del cóndor extinto (Ciconiiformes: Vulturidae) del Cuaternario cubano. Ornitol. Neotrop. 2000, 11, 109–122. [Google Scholar]
- Stucchi, M.; Emslie, S.D. A new condor (Ciconiiformes, Vulturidae) from the late Miocene/early Pliocene Pisco formation, Peru. Condor 2005, 107, 107–113. [Google Scholar] [CrossRef]
- Galetti, M.; Moleón, M.; Jordano, P.; Pires, M.M.; Guimarães, P.R.; Pape, T.; Nichols, E.; Hansen, D.; Olesen, J.M.; Munk, M.; et al. Ecological and evolutionary legacy of megafauna extinctions. Biol. Rev. 2018, 93, 845–862. [Google Scholar] [CrossRef]
- Perrig, P.L.; Fountain, E.D.; Lambertucci, S.A.; Pauli, J.N. Demography of avian scavengers after Pleistocene megafaunal extinction. Sci. Rep. 2019, 9, 9680. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, W.J. The Exploration of the Potter Creek Cave; University of California Press: Berkeley, CA, USA, 1907; Volume 2. [Google Scholar]
- Simpson, G.G. Additions to the Pleistocene of Florida; American Museum of Natural History: New York, NY, USA, 1930. [Google Scholar]
- Howard, H. Quaternary avian remains from Dark Canyon Cave, New Mexico. Condor 1971, 73, 237–240. [Google Scholar] [CrossRef]
- Alvarenga, H.M.; Olson, S.L. A new genus of tiny condor from the Pleistocene of Brazil (Aves: Vulturidae). Proc. Biol. Soc. Wash. 2004, 117, 1–9. [Google Scholar]
- Noriega, J.I.; Areta, J.I. First record of Sarcoramphus dumeril 1806 (Ciconiiformes: Vulturidae) from the Pleistocene of Buenos Aires province, Argentina. J. S. Am. Earth Sci. 2005, 20, 73–79. [Google Scholar] [CrossRef]
- Alvarenga, H.; Brito, G.R.; Migotto, R.; Hubbe, A.; Höflibg, E. Pleistovultur nevesi gen. et sp. nov. (Aves: Vulturidae) y la diversidad de cóndores y buitres en el Pleistoceno de América del Sur. Ameghiniana 2008, 45, 613–618. [Google Scholar]
- Tyrberg, T. The Late Pleistocene continental avian extinction: An evaluation of the fossil evidence. Oryctos 2008, 7, 249–269. [Google Scholar]
- Tambussi, C.P.; Degrange, F.J. The dominance of zoophagous birds: Just a cliché? In South American and Antarctic Continental Cenozoic Birds; Tambussi, C.P., Disparities, F., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 87–98. [Google Scholar]
- Jones, W.W.; Rinderknecht, A.; Vezzosi, R.I.; Montenegro, F.; Ubilla, M. First report of large cathartids (Aves, Cathartidae) from the late Pleistocene of Uruguay. J. S. Am. Earth Sci. 2020, 107, 102946. [Google Scholar] [CrossRef]
- Wallace, M.P.; Temple, S.A. Competitive interactions within and between species in a guild of avian scavengers. Auk 1987, 104, 290–295. [Google Scholar] [CrossRef]
- Hertel, F. Diversity in body size and feeding morphology within past and present vulture assemblages. Ecology 1994, 75, 1074–1084. [Google Scholar] [CrossRef]
- Grigg, N.P.; Krilow, J.M.; Gutierrez-Ibanez, C.; Wylie, D.R.; Graves, G.R.; Iwaniuk, A.N. Anatomical evidence for scent guide foraging in the turkey vulture. Sci. Rep. 2017, 7, 17408. [Google Scholar] [CrossRef]
- Kluever, B.M.; Pfeiffer, M.B.; Barras, S.C.; Dunlap, B.G.; Humberg, L.A. Black vulture conflict and management in the United States: Damage trends, management overview, and research needs. Hum.-Wildl. Interact. 2020, 14, 376–389. [Google Scholar]
- Behmke, S.; Fallon, J.; Duerr, A.E.; Lehner, A.; Buchweitz, J.; Katzner, T. Chronic lead exposure is epidemic in obligate scavenger populations in eastern North America. Environ. Int. 2015, 79, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Ballejo, F.; Plaza, P.I.; Lambertucci, S.A. The conflict between scavenging birds and farmers: Field observations do not support people’s perceptions. Biol. Conserv. 2020, 248, 108627. [Google Scholar] [CrossRef]
- Rideout, B.A.; Stalis, I.; Papendick, R.; Pessier, A.; Puschner, B.; Finkelstein, M.E.; Smith, D.R.; Johnson, M.; Mace, M.; Stroud, R.; et al. Patterns of mortality in free-ranging California Condors (Gymnogyps californianus). J. Wildl. Dis. 2012, 48, 95–112. [Google Scholar] [CrossRef] [PubMed]
- Naveda-Rodríguez, A.; Vargas, F.H.; Kohn, S.; Zapata-Ríos, G. Andean condor (Vultur gryphus) in Ecuador: Geographic distribution, population size and extinction risk. PLoS ONE 2016, 11, e0151827. [Google Scholar] [CrossRef] [PubMed]
- Plaza, P.I.; Lambertucci, S.A. Ecology and conservation of a rare species: What do we know and what may we do to preserve Andean condors? Biol. Conserv. 2020, 251, 108782. [Google Scholar] [CrossRef]
- Chamberlain, C.P.; Waldbauer, J.R.; Fox-Dobbs, K.; Newsome, S.D.; Koch, P.L.; Smith, D.R.; Church, M.E.; Chamberlain, S.D.; Sorenson, K.J.; Risebrough, R. Pleistocene to recent dietary shifts in California condors. Proc. Natl. Acad. Sci. USA 2005, 102, 16707–16711. [Google Scholar] [CrossRef]
- Noriega, J.I.; Tonni, E.P. Geronogyps reliquus Campbell (Ciconiiformes: Vulturidae) en el Pleistoceno tardío de la provincia de Entre Ríos y su significado paleoambiental. Ameghiniana 2007, 44, 245–250. [Google Scholar]
- Cenizo, M.M.; Agnolin, F.L.; Pomi, L.H. A new Pleistocene bird assemblage from the southern Pampas (Buenos Aires, Argentina). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 420, 65–81. [Google Scholar] [CrossRef]
- Wallace, R.B.; Reinaga, A.; Piland, N.; Piana, R.; Vargas, F.H.; Zegarra, R.E.; Alvarado, S.; Kohn, S.; Lambertucci, S.A.; Alarcón, P.; et al. Defining Spatial Conservation Priorities for the Andean Condor (Vultur gryphus). J. Raptor Res. 2021, 56, 1–16. [Google Scholar] [CrossRef]
- Sáenz-Jiménez, F.; Rojas-Soto, O.; Pérez-Torres, J.; Martínez-Meyer, E.; Sheppard, J.K. Effects of climate change and human influence in the distribution and range overlap between two widely distributed avian scavengers. Bird Conserv. Int. 2021, 31, 77–95. [Google Scholar] [CrossRef]
- Carrete, M.; Lambertucci, S.A.; Speziale, K.; Ceballos, O.; Travaini, A.; Delibes, M.; Hiraldo, F.; Donázar, J.A. Winners and losers in human-made habitats: Interspecific competition outcomes in two Neotropical vultures. Anim. Conserv. 2010, 13, 390–398. [Google Scholar] [CrossRef]
- Lambertucci, S.A.; Ruggiero, A. Cliffs used as communal roosts by Andean Condors protect the birds from weather and predators. PLoS ONE 2013, 8, e67304. [Google Scholar] [CrossRef]
- Walters, J.R.; Derrickson, S.R.; Michael, D.; Haig, S.M.; Marzluff, J.M.; Wunderle, J.M. Status of the California Condor (Gymnogyps californianus) and efforts to achieve its recovery. Auk 2010, 127, 969–1001. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Dinnage, R.; Tilman, D. Phylogenetic diversity promotes ecosystem stability. Ecology 2012, 93, S223–S233. [Google Scholar] [CrossRef]
- Jetz, W.; Thomas, G.H.; Joy, J.B.; Redding, D.W.; Hartmann, K.; Mooers, A.O. Global distribution and conservation of evolutionary distinctness in birds. Curr. Biol. 2014, 24, 919–930. [Google Scholar] [CrossRef]
- DeVault, T.L.; Beasley, J.C.; Olson, Z.H.; Moleón, M.; Carrete, M.; Margalida, A.; Sánchez-Zapata, J.A. Ecosystem services provided by avian scavengers. In Why Birds Matter. Avian Ecological Function and Ecosystem Services; Şekercioğlu, C.H., Wenny, D.G., Whelan, C.J., Eds.; University of Chicago Press: Chicago, IL, USA, 2016; pp. 235–270. [Google Scholar]
- Van Overveld, T.; Blanco, G.; Moleón, M.; Margalida, A.; Sánchez-Zapata, J.A.; de la Riva, M.; Donázar, J.A. Integrating vulture social behavior into conservation practice. Condor 2020, 12, duaa035. [Google Scholar] [CrossRef]
- Santangeli, A.; Girardello, M.; Buechley, E.; Botha, A.; Minin, E.D.; Moilanen, A. Priority areas for conservation of Old World Vultures. Conserv. Biol. 2019, 33, 1056–1065. [Google Scholar] [CrossRef]
- Antonelli, A.; Zizka, A.; Carvalho, F.A.; Scharn, R.; Bacon, C.D.; Silvestro, D.; Condamine, F.L. Amazonia is the primary source of Neotropical biodiversity. Proc. Natl. Acad. Sci. USA 2018, 115, 6034–6039. [Google Scholar] [CrossRef]
- Guevara-Andino, J.E. Integrating Community Phylogenetics Phylogenetic Beta Diversity to Understand Amazonian Trees Community Assembly: A Biogeographical Conservation Approach. Ph.D. Thesis, University of California, Berkeley, CA, USA, 2017. [Google Scholar]
- Gumbs, R.; Gray, C.L.; Böhm, M.; Hoffmann, M.; Grenyer, R.; Jetz, W.; Meiri, S.; Roll, U.; Owen, N.R.; Rosindell, J. Global priorities for conservation of reptilian phylogenetic diversity in the face of human impacts. Nat. Commun. 2020, 11, 2616. [Google Scholar] [CrossRef]
- Velazco, S.J.E.; Svenning, J.C.; Ribeiro, B.R.; Laureto, L.M.O. On opportunities and threats to conserve the phylogenetic diversity of Neotropical palms. Divers. Distrib. 2021, 27, 512–523. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Rank | Model | Parameters | d | e | J | AICc |
|---|---|---|---|---|---|---|
| 1 | DEC | 2 | 0.071 | 0.0084 | 0 | 22.07 |
| 2 | DIVALIKE | 2 | 0.084 | 0.0062 | 0 | 22.72 |
| 3 | BAYAREALIKE | 2 | 0.081 | 0.039 | 0 | 24.91 |
| 4 | DEC + J | 3 | 0.044 | 1.0 × 10−12 | 0.38 | 26.49 |
| 5 | DIVALIKE + J | 3 | 0.056 | 1.0 × 10−12 | 0.36 | 27.49 |
| 6 | BAYAREALIKE + J | 3 | 0.037 | 1.0 × 10−7 | 0.29 | 29.89 |
| Rank | Model | Parameters | d | e | J | AICc |
|---|---|---|---|---|---|---|
| 1 | DEC | 2 | 0.078 | 0.0089 | 0 | 31.73 |
| 2 | DIVALIKE | 2 | 0.088 | 1.0 × 10−12 | 0 | 31.93 |
| 3 | BAYAREALIKE | 2 | 0.10 | 0.078 | 0 | 33.76 |
| 4 | DEC + J | 3 | 0.061 | 1.0 × 10−12 | 0.56 | 36.6 |
| 5 | DIVALIKE + J | 3 | 0.071 | 1.0 × 10−12 | 0.50 | 37.57 |
| 6 | BAYAREALIKE + J | 3 | 0.011 | 0.079 | 1.0 × 10−5 | 40.76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortés-Díaz, D.; Buitrago-Torres, D.L.; Restrepo-Cardona, J.S.; Estellés-Domingo, I.; López-López, P. Bridging Evolutionary History and Conservation of New World Vultures. Animals 2023, 13, 3175. https://doi.org/10.3390/ani13203175
Cortés-Díaz D, Buitrago-Torres DL, Restrepo-Cardona JS, Estellés-Domingo I, López-López P. Bridging Evolutionary History and Conservation of New World Vultures. Animals. 2023; 13(20):3175. https://doi.org/10.3390/ani13203175
Chicago/Turabian StyleCortés-Díaz, Daniela, Diana L. Buitrago-Torres, Juan Sebastián Restrepo-Cardona, Irene Estellés-Domingo, and Pascual López-López. 2023. "Bridging Evolutionary History and Conservation of New World Vultures" Animals 13, no. 20: 3175. https://doi.org/10.3390/ani13203175
APA StyleCortés-Díaz, D., Buitrago-Torres, D. L., Restrepo-Cardona, J. S., Estellés-Domingo, I., & López-López, P. (2023). Bridging Evolutionary History and Conservation of New World Vultures. Animals, 13(20), 3175. https://doi.org/10.3390/ani13203175