Simple Summary
Tritrichomonas muris (T. muris) mainly parasitizes the ceca of different rodent species worldwide. To date, there are no ideal methods for the detection of T. muris infections in laboratory mice. This study developed a nested PCR reaction using a novel specific primer based on the conserved regions of the SSU rRNA gene of T. muris for a molecular epidemiological survey (investigation) of T. muris infections in laboratory mice. The results showed that the nested PCR system has higher sensitivity, reliability, and specificity. The nested PCR system showed an infection rate of T. muris of 18.96% (58/306), which was higher than the infection rate of 14.05% (43/306) that was detected via smear microscopy in fecal samples from five mouse strains. The sensitivity and specificity of nested PCR in detecting T. muris was found to be 100%, and it demonstrated a 26% increase in diagnostic sensitivity compared to the smear microscopy method. The present study provides a new method for the molecular epidemiological investigation of T. muris infections in laboratory mice.
Abstract
A variety of rodent ceca are parasitized by Tritrichomonas muris (T. muris), a flagellated protozoan. To date, there are no ideal methods for the detection of T. muris infections in laboratory mice; thus, new molecular methodologies for its specific detection need to be developed. In this study, using staining and SEM, it was observed that T. muris has a pear-shaped body and contains three anterior flagella. A nested PCR system with novel specific primers was designed based on the conserved regions of the SSU rRNA gene of T. muris. The nested PCR system for T. muris showed good specificity and high sensitivity for at least 100 T. muris trophozoites/mL and 0.1 ng/μL of fecal genomic DNA, which means that 176 trophozoites per gram of mouse feces could be detected. When using this nested PCR system, the detection rate was 18.96% (58/306), which was higher than the detection rate of 14.05% (43/306) detected via smear microscopy in fecal samples from five mouse strains. The sensitivity and specificity of nested PCR in detecting T. muris was found to be 100%, and it demonstrated a 26% increase in diagnostic sensitivity compared to the smear microscopy method in the present study. In conclusion, the nested PCR developed with novel primers based on the SSU rRNA gene of T. muris has good accuracy, specificity, and sensitivity for the detection of T. muris infections in laboratory mice.
1. Introduction
Tritrichomonas muris (T. muris), which belongs to the large group Tritrichomonadea of the phylum Parabasalia, is a unicellular, aerotolerant, flagellated protozoan parasite that was once considered a conditional pathogenic pathogen or a nonpathogenic member of murine flagellated protozoans in wild and laboratory mice [1,2,3,4]. However, studies have found that T. muris infections are related to host immune responses, which, nowadays, can change mucosal T-cell homeostasis and susceptibility to colitis [4]. As a result of T. muris infections, the tumor suppressor p53 can lead to an intestinal type 2 immune response [5]. Thus, T. muris infections seriously endanger the health of laboratory mice and test indicators.
T. muris mainly parasitizes the large intestines, especially the ceca, of different rodent species [1,6], including mice, rats, and hamsters [2,7,8,9,10]. T. muris is widespread worldwide, especially in Asia, North America, and Europe, where the infection rate is as high as 100 percent [8,9,10]. In addition, a mixed infection was observed in the intestinal tract of animals, which were simultaneously infected not only with T. muris but also with G. duodenalis, P. hominis, Cryptosporidium spp., and other parasites [11,12].
Currently, Trichomonas detection depends on fecal smear microscopic examinations and cultures, which have been regarded as the traditional detection method for many decades [13,14]. Although smear microscopy is capable of quick initial detection, its detection rate is low [15]. Researchers have reported that the 28 s rRNA gene of T. muris can be amplified via qPCR for species identification and classification [16]. The disadvantages of qPCR include the requirement for professional instruments, the use of fluorescent dyes, and the high demand for primers [17]. However, smear microscopy remains the main method of detection in clinical settings. Thus, to date, there are no on-site, reliable, and sensitive pathogen-detecting methods for T. muris infections in experimental animals.
The small subunit ribosomal RNA (SSU rRNA) gene, or the 16S-like ribosomal RNA gene, has proven to be an invaluable tool in molecular phylogenetic studies based on specific characteristics, such as its ubiquity, size, and high conservatism [18]. This molecule has accounted for numerous new insights in evolutionary biology, and its impact on our understanding of the origin and diversity of eukaryotes has been tremendous [19,20,21]. Due to its popularity as a molecular marker, SSU rRNA is widely used to detect and identify different genetic sequences of protozoans, such as T. vaginalis, Tritrichomonas foetus (T. foetus), G. duodenalis, and Cryptosporidium spp., but not T. muris [22,23,24,25,26,27,28]. Therefore, it is necessary to design a novel primer for clinical epidemiological studies of this pathogen based on SSU rRNA.
Nested PCR exhibits better performance with a high degree of sensitivity, specificity, repeatability, and reliability, and it eliminates the possibility of false positive signals [29]. Here, our study offers novel primers based on the SSU rRNA gene of T. muris to develop a nested PCR system that is specific and sensitive for the detection of T. muris infections in laboratory mice. This will make an important contribution to the field and ensure the quality of laboratory mice and experimental results.
2. Materials and Methods
2.1. Ethics Statement
All collection procedures were conducted in strict accordance with the guidelines of the Animal Care and Welfare Committee of Jilin University (IACUC Permit Number: 201606122).
2.2. Collection of Fecal Samples and Preparation of Genomic DNA
Randomly selected laboratory mice were provided by experimental animal breeding companies in different regions of China. Genomic DNA from fresh fecal samples was extracted using the QIAamp DNA stool mini kit (Qiagen, Valencia, CA, USA), according to the manufacturer’s instructions.
Additionally, other parasite genomic DNA, including P. hominis, G. duodenalis, Cryptosporidium tyzzeri (C. tyzzeri), T. vaginalis, Escherichia coli (E. coli), Trichomonas gallinae (T. gallinae) and Salmonella typhimurium (S. typhimurium), was extracted using the TIANamp Genomic DNA Kit (Tiangen, Beijing, China) and preserved at −40 °C in our laboratory.
2.3. Isolation of T. muris
The isolation and culture of T. muris were performed using a modified protocol described in the literature by other researchers [4,30,31]. Briefly, fecal contents were harvested from mice, passed through a 40 µm filter, and washed three times with PBS. The T. muris-containing mixtures were further treated through gradient centrifugation in Percoll (GE Healthcare, Uppsala, Sweden) at 1000× g for 15 min, and the purified T. muris trophozoites were collected from the interface using between 40% Percoll (GE Healthcare, Uppsala, Sweden) and 80% Percoll (GE Healthcare, Uppsala, Sweden). The number and viability of the isolated T. muris were determined by counting them using a hemocytometer.
T. muris was cultured under a density of 5 × 105 trophozoites per mL of complete growth media, which consisted of TrichoselTM broth (Becton Dickinson, San Jose, CA, USA) supplemented with 10% heat-inactivated horse serum (Gibco, Grand Island, NY, USA), 1% amphotericin B (Yuanye, Shanghai, China), 1% gentamicin (Yuanye, Shanghai, China), 1% penicillin (Yuanye, Shanghai, China), 1% streptomycin (Yuanye, Shanghai, China), and 1% vancomycin (Yuanye, Shanghai, China), and the pH was adjusted to 7.0. The T. muris trophozoites were cultured at 37 °C in a constant-temperature culture. The genomic DNA of T. muris trophozoites was extracted using a TIANamp Genomic DNA Kit (Tiangen, Beijing, China) and preserved at −40 °C.
2.4. Light Microscopy Observation
T. muris was observed microscopically using an Olympus CX43 microscope (Olympus, Tokyo, Japan) and photographed using an Olympus DP-71 camera (Olympus, Tokyo, Japan) with DP Controller 3.1.267 software. Pictures of T. muris at different staining morphologies were recorded at 400 or 1000 times magnification.
2.5. Scanning Electron Microscopy Observation
For SEM sample preparation, T. muris was cultured to 105 per mL in TrichoselTM broth (Becton Dickinson, San Jose, CA, USA) at 37 °C, collected via centrifugation at 1000× g for 10 min, and then, washed three times with PBS. A cell climbing test was conducted using a modified protocol described by other researchers’ literature [4,30,31]. The glass slides were pretreated with acetic acid for 2 h and absolute ethanol for 1 h, and washed with PBS 3 times. Post-drying, the slides were wrapped in aluminum foil and baked at 180 °C for 2 h in an oven (Thermo, Waltham, MA, USA). Tritrichomonas muris was isolated from the cecal contents of WT mice as described above, and suspended in PBS. 1.0 × 106 T. muris trophozoites were seeded on poly-L-lysine-coated coverslips, cultured for 1 h at 37 °C in incubators (Thermo, Waltham, MA, USA), and fixed in 2.5% glutaraldehyde (Solarbio, Beijing, China) in a 0.1 M cacodylate buffer (Solarbio, Beijing, China), pH 7.2. Following 3 buffer rinses, the protozoa were filtered through polycarbonate membrane filters (1.2 μm, Millipore, Darmstadt, Germany), post-fixed for 30 min in 1% OsO4 (Aladdin, Shanghai, China) in 0.1 M cacodylate buffer (Solarbio, Beijing, China), and dehydrated in a graded series of ethanol. The dry filter pieces were attached to stubs and subsequently coated with gold. The morphologies of the anterior flagellum (AF), recurrent flagellum (RF), undulating membrane (UM), and axostyle (AX) in T. muris were examined under an S-3400N scanning electron microscope (SEM, S-3400N; Hitachi, Tokyo, Japan).
2.6. Primer Design
Nested PCR primers were designed using the Premier 5.0 software based on conserved region sequences of the SSU rRNA gene deposited in GenBank (accession No. AY886846.1). The first-round PCR primer sets were as follows: the forward primer F1: 5′-CTG TGA ACA AAT CAG GAC GCT-3′, and the reverse primer R1: 5′-ACC TTT GTG CGT ACA CTC CG-3′. The second-round PCR primer sets were as follows: the forward primer F2: 5′-GAT TCA GAT AAC GAG CGA GAT T-3′, and the reverse primer R2: 5′-CCT TTG TGC GTA CAC TCC G-3′. The primers were synthesized by Comate Biological Technology Co., Ltd. (Changchun, China).
2.7. Optimization of the Nested PCR Reaction System
The nested PCR reaction system was mainly optimized through the following two aspects: Mg2+ concentrations and annealing temperatures. The nested PCR assays were performed at Mg2+ concentrations of 0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 3.5, 4.0, 4.5, and 5 mmol/L, respectively. The annealing temperatures of the first round were set with the following 10 gradients: 56, 57.0, 58.0, 59.0, 60.0, 61.0, 62.0, 63.0, 64.0, and 65.0 °C, respectively. The annealing temperatures of the second round were set with the following 10 gradients: 51.0, 52.0, 53.0, 54.0, 55.0, 56.0, 57.0, and 58.0 °C, respectively.
The reaction system was prepared as follows: The first-round PCR mixture (50 μL) contained 0.25 μL of Taq DNA polymerase (5 U/μL; Takara, Dalian, China), 5 μL of 10 × PCR buffer without Mg2+, 4 μL of dNTP mixture (2.5 mM each), 1 μL of forward and reverse primers (10 pmol/μL), 1 μL of a T. muris genomic DNA template, and different concentrations of MgCl2. In the second round of PCR, 1 µL of the PCR product from the first round was used as the DNA template for the PCR reaction, which included a standard PCR reaction mixture.
The reaction program was set as follows: pre-denaturation at 95 °C for 5 min, followed by 30 cycles of denaturation at 95 °C for 50 s, annealing at 55 °C for 50 s, extension at 72 °C for 1 min, and a final extension at 72 °C for 12 min. In order to avoid contamination, PCR was carried out under stringent conditions, as described previously [32,33]. Each batch of samples being analyzed contained both negative and positive controls. A standard PCR reaction, which contained water instead of sample, was included as a negative control, while the positive control contained 10 ng of T. muris genomic DNA instead of sample. The amplified products were electrophoresed on 1% agarose gel and stained with ethidium bromide to visualize the bands.
2.8. Sequence Analysis
The purified PCR product was cloned into the pMD-18T vector (TaKaRa, Dalian, China) and transformed into E. coli DH5α competent cells, to obtain the recombinant plasmid pMD-18T-muris. The single-cloning recombinant pMD-18T-muris bacterial colony was identified through PCR, and sequenced by Comate Biological Technology Co., Ltd.
The T. muris gene sequences were submitted to NCBI (Genbank No. OP437559), amplified via nested PCR, and cloned using E. coli DH5α. The SSU rRNA gene sequences were downloaded from NCBI, as shown in Table 1. Multiple sequence alignment was used to construct a phylogenetic tree using the maximum likelihood (ML) method. The phylogenetic tree was constructed using the MEGA software (version 6.0) [34].

Table 1.
Information about species in homologous evolution.
2.9. Validation of the Nested PCR Assays
2.9.1. Specificity Validation of the Nested PCR Reaction
In order to determine the specificity of the nested PCR reaction for T. muris, different parasites’ genomic DNA, including P. hominis, G. duodenalis, C. tyzzeri, T. vaginalis, E. coli, T. gallinae, and S. typhimurium was used for the nested PCR assays for T. muris, while the recombinant plasmid pMD-18T-muris was used as a positive control and ddH2O was set as a negative control. The reaction system and program were performed as described above.
2.9.2. Sensitivity Validation of the Nested PCR Reaction
In order to evaluate the sensitivity of the nested PCR reaction for the detection of fecal mouse samples infected by T. muris, mouse feces were used to extract DNA. The feces were homogenized on a homogenizer (Huxi, Shanghai, China). The genomic DNA was extracted from the fecal samples using a QIAamp DNA stool mini kit (Qiagen, Valencia, CA, USA), according to the manufacturer’s instructions. The genomic DNA concentration of the mouse fecal samples infected by T. muris was measured, after which it was diluted as follows: 200 ng/μL, 100 ng/μL, 10 ng/μL, 1 ng/μL, 0.1 ng/μL, 0.01 ng/μL, 0.001 ng/μL, and 0.0001 ng/μL. The sensitivity of the nested PCR reactions was determined using the above genomic DNA with different concentrations as templates (1 μL for each reaction), while the recombinant plasmid pMD-18T-muris was used as a positive control. The PCR products were resolved by using electrophoresis on 1% agarose gel and stained with ethidium bromide. Subsequently, the number of genomic DNAs equivalent to the number of T. muris parasites per gram of feces was converted.
In addition, the detection sensitivity of T. muris trophozoites was evaluated using nested PCR reactions. The trophozoite cells were counted and diluted with sterilized ddH2O to final concentrations ranging from 106 to 100 trophozoites/mL with a ten-fold gradient dilution. Genomic DNA was extracted from 1 mL of the trophozoites and used as template DNA in the nested PCR assay, while the recombinant plasmid pMD-18T-muris was used as a positive control. The PCR products were resolved via electrophoresis on 1% agarose gel and stained with ethidium bromide to observe the sensitivity to different T. muris quantities.
2.10. Clinical Application of the New Primer in the Nested PCR System for T. muris
In order to evaluate the application effect of the new primers in the nested PCR of T. muris, 306 feces samples of laboratory mice from different experimental animal breeding companies in China were detected via smear microscopy and nested PCR. The feces samples were collected from March 2021 to July 2022, of which 60 were collected from C57/BL-6J mice, 60 from Kunming mice, 60 from BALB/C mice, 60 from Gerbil mice, and 66 from BALB/C nude mice, respectively. Firstly, the fresh feces of each laboratory mouse were collected and divided into two parts. One part of the feces was detected via smear microscopy (three smears were made for each sample), and the detection rate of T. muris was counted and analyzed. Another part of the feces was used for the fecal genomic DNA extraction and nested PCR detection of T. muris. Additionally, a statistical analysis was carried out on the infection rate of T. muris.
2.11. Data Analysis
A statistical analysis was performed using the SPSS software version 20.0 (IBM, Armonk, NY, USA). A chi-square test was used to estimate the statistical significance of the laboratory mice’s fecal samples collected from different species. All statistical tests were two-sided. Odds ratios (ORs) with 95% confidence intervals were used to assess the strength of associations. All ORs were adjusted for both age and sex. A value of p < 0.05 was considered statistically significant.
3. Results
3.1. Morphological Identification of T. muris
T. muris was isolated and purified from the stools of laboratory mice and observed using smear microscopy. The results revealed that T. muris has a pear-shaped body and contains three anterior flagella, a recurrent flagellum, and nuclei structures (Figure 1). The SEM results showed that T. muris is a pear-shaped cell that is free-living and free-swimming. Three flagella protrude from the region of the pelta at the anterior end of the trophozoid cell. The recurrent flagellum, which is attached to the undulating membrane, runs along the entire cell surface toward the posterior end (Figure 2).
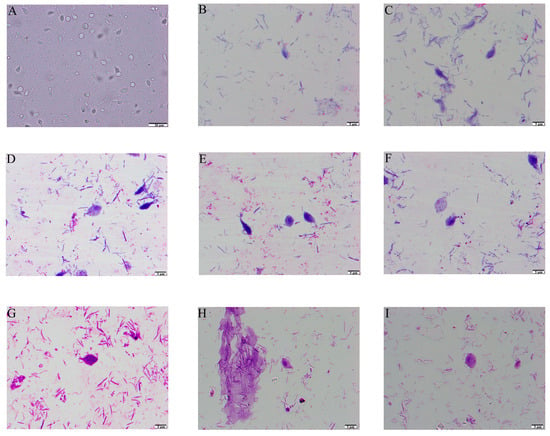
Figure 1.
Light microscopy observation of T. muris. (A) Optical microscope observation with 400 times magnification. (B,C) Swiss staining observation with 1000 times magnification. (D–H) Swiss-Giemsa staining observation with 1000 times magnification and the nuclei were stained in blue. (G–I) Flagella-staining observation with the anterior flagellum and the posterior flagellum stained in blue, and the nuclei stained in dark blue.
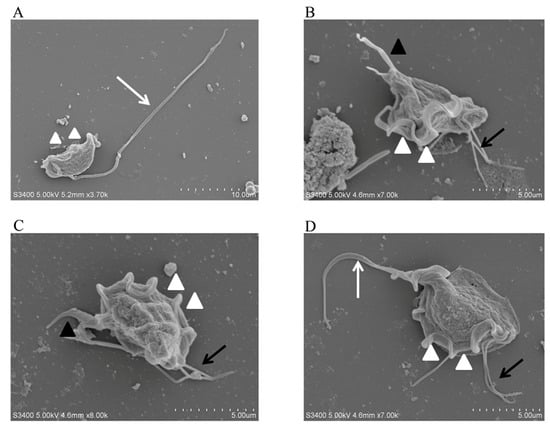
Figure 2.
SEM observation of T. muris. The morphology of T. muris under different conditions was observed using a scanning electron microscope (SEM). (A–D) The white arrows represent the recurrent flagellum (RF), the white triangles represent the undulating membrane (UM), the black arrows represent the anterior flagella (AF), the black triangles represent the axostyle (AX).
3.2. Sequence Analysis of T. muris’ Small Subunit Ribosomal RNA (SSU rRNA)
The SSU rRNA gene of T. muris was amplified using nested PCR assays and designed primers. The amplicon sizes of the first- and the second-round PCR products were 906 and 384 base pairs, respectively. A clear band with an expected size was obtained through agarose gel electrophoretic analysis (Figure 3A). The nested PCR-amplified products were subsequently cloned into pMD-18T and sequenced. The sequence alignment results revealed that the obtained SSU rRNA gene sequence isolated from the experimental mice belonged to the Tritrichomonas branch and shared a 98% gene sequence identity with the SSU rRNA gene of T. muris (Genbank No. AY886846.1; Figure 3B).
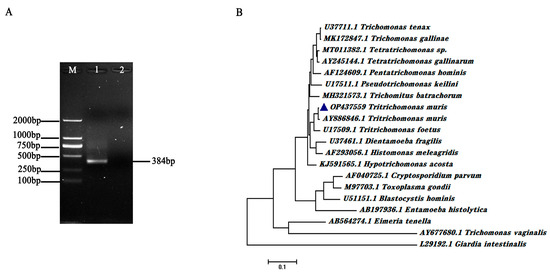
Figure 3.
Amplification performance of the nested PCR primer sets and sequence alignment of T. muris SSU rRNA gene. (A) Validation of the SSU rRNA nested PCR primer sets using isolated T. muris genomic DNA as templates. The amplification products were detected through agarose gel electrophoretic analysis. M represents DL2000 DNA Marker. (B) Sequence alignment of isolated T. muris SSU rRNA and that in other parasites, including Tririchomonas muris (AY886846.1), Tritrichomonas foetus (U17509.1), Trichomonas tenax (U37711.1), Pentatrichomonas hominis (AF124609.1), Dientamoeba fragilis (U37461.1), Entamoeba histolytica (AB197936.1), Cryptosporidium parvum (AF040725.1), Blastocystis hominis (U51151.1), Toxoplasma gondii (M97703.1), Eimeria tenella (AB564274.1), Giardia intestinalis (L29192.1), Trichomonas vaginalis (AY677680.1), Trichomonas gallinae (MK172847.1), Tetratrichomonas sp. (MT011382.1), Tetratrichomonas gallinarum (AY245144.1), Pseudotrichomonas keilini (U17511.1), Histomonas meleagridis (AF293056.1), Trichomitus batrachorum (MH321573.1), and Hypotrichomonas acosta (KJ591565.1). The blue triangles represent the SSU rRNA gene sequence of the isolated T. muris (OP437559).
3.3. Determination of the Optimal Nested PCR
Mg2+ concentrations and annealing temperatures were the two key factors for improving the performance of Taq DNA polymerase amplification and primer–template combinations. The Mg2+ concentration optimization results showed that the nested PCR assays could obtain target bands when Mg2+ concentrations were in a range from 1.0 mmol/L to 5.0 mmol/L, and the brightness of the target bands first displayed an increased, and then, a decreased trend along with the Mg2+ concentration changes. The amounts of amplification products peaked when using 4.0 mmol/L Mg2+ (Figure 4A).
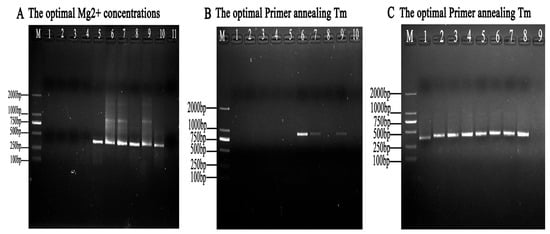
Figure 4.
Optimization of the nested PCR system parameters. (A) Optimization of the Mg2+ concentrations. The nested PCR assays were carried out with Mg2+ concentrations of 0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 3.5, 4.0, 4.5, and 5 mmol/L (lanes 1 to 10), respectively. (B) Optimization of the first-round annealing temperature in the nested PCR reaction system. The annealing temperature was set with 10 gradients. Lanes 1 to 10 indicate 56, 57.0, 58.0, 59.0, 60.0, 61.0, 62.0, 63.0, 64.0, and 65.0 °C, respectively. (C) Optimization of the second-round annealing temperature in the nested PCR reaction system. The annealing temperature was set with 10 gradients. Lanes 1 to 8 show 51.0, 52.0, 53.0, 54.0, 55.0, 56.0, 57.0, and 58.0, respectively. M represents DL2000 DNA Marker.
To justify the reliability of the nested PCR reaction system, different ranges of annealing temperature values were optimized. By comparing the Tm values of the primer, the following Tm ranges were established for each group of nested PCR reactions: For the determination of the optimal temperature screen, the first-round PCR results showed that a weak target band occurred at 61.0 °C (Figure 4B). The second-round PCR results showed that targeted bands were seen with annealing temperatures ranging from 51.0 °C to 58.0 °C (Figure 4C). Finally, the optimal annealing temperature was set to 56.0 °C (Table 2).
Table 2.
Optimal conditions of nested PCR reaction system.
3.4. Specificity of the Nested PCR System
To ensure the specificity of the nested PCR reaction, different species of parasites were selected and used for this assay. The results showed no target bands when using P. hominis, G. duodenalis, C. tyzzeri, T. vaginalis, E. coli, T. gallinae, and S. typhimurium as templates, while clear bands with a size of 384 bp occurred with the introduction of T. muris, which illustrated that the established nest PCR assays are highly specific to T. muris and had no cross-reactions with other parasites (Figure 5A).
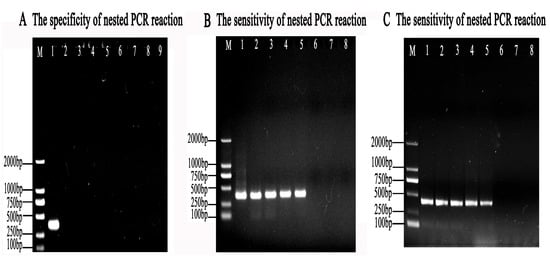
Figure 5.
The specificity and sensitivity of the nested PCR system. (A) The specificity of the nested PCR system for T. muris was evaluated using pMD-18T-muris as a positive control template (lane 1); ddH2O as a negative control template (lane 2); and G. duodenalis (lane 3), C. tyzzeri (lane 4), E. coli (lane 5), P. hominis (lane 6), T. vaginalis (lane 7), T. gallinae (lane 8), and S. typhimurium (lane 9) as the tested templates. M represents the DL2000 DNA Marker. (B) The sensitivity of the second-round nested PCR for mice feces infected by T. muris was detected using template concentrations of 200, 100, 10, 1, 0.1, 0.01, 0.001, and 0.0001ng/μL genomic DNAs (lane 1–lane 7), respectively. (C) The sensitivity of the second-round nested PCR system for T. muris was measured using DNA of 106, 105, 104, 103, 102, 101, and 100 T. muris (lane 1–lane 7), respectively. Lane 8 represents no template control. M represents the DL2000 DNA Marker.
3.5. Sensitivity of the Nested PCR
At least 0.1 ng/μL of genomic DNA of the mouse feces infected with T. muris could be detected using the nested PCR system (Figure 5B), which amounted to 176 trophozoites/g of feces.
In addition, the genomic DNA of at least 100 T. muris trophozoites/mL could be detected using the nested PCR system according to gel electrophoresis after PCR amplification (Figure 5C).
3.6. Clinical Application of the New Primer of the Nested PCR System for T. muris
The overall infection rate of T. muris measured using nested PCR was 18.96% (58/306), while the infection rate detected via smear microscopy was 14.05% (43/306), which was lower than that of the nested PCR. The infection rates of T. muris in C57/BL-6J mice detected using nested PCR were 13.33% (8/60) and 8.33% (5/60) via smear microscopy examination (χ2 = 6.708; df = 1; p < 0.01). The infection rates of T. muris in Kunming mice detected using nested PCR were 21.67% (13/60) and 20% (12/60) via smear microscopy examination (χ2 = 2.048; df = 1; p > 0.01). The infection rates of T. muris in BALB/C mice detected using nested PCR were 20% (12/60) and 13.33% (8/60) via smear microscopy examination (χ2 = 2.727; df = 1; p < 0.05). The infection rates of T. muris in Gerbil mice detected using nested PCR were 33.33% (20/60) and 26.67% (16/60) via smear microscopy examination. The infection rates of T. muris in BALB/C nude mice detected using nested PCR were 7.5% (5/60) and 3.03% (2/60) via smear microscopy examination (χ2 = 13.11; df = 1; p < 0.01) (Table 3).
Table 3.
Occurrence of T. muris infections in laboratory mice.
The present study determined the sensitivity and specificity of nested PCR in detecting T. muris to be 100%, with no instances of false negative or false positive samples. In contrast, smear microscopy exhibited a sensitivity percentage of 74% and a specificity percentage of 100% in the clinical diagnosis of T. muris, with 15 false negative samples and no false positive samples. The nested PCR method demonstrated a 26% increase in diagnostic sensitivity compared to the smear microscopy method in the clinical application of detecting T. muris in 306 laboratory mice (Table 4).
Table 4.
Diagnostic sensitivity and specificity of the assays used in this study for the diagnosis of T. muris.
4. Discussion
The majority of Trichomonas monitoring was based on smear microscopy. In the case of poor experimental conditions, the sensitivity of direct smear detection averages between 50% and 60% [35,36,37]. Smear microscopy is fast and inexpensive but has low sensitivity and specificity [38]. For the sake of abandoning the shortcomings of smear microscopy, the detection of Trichomonas using nested PCR was developed, which has the advantages of accurate reaction, excellent specificity, and high sensitivity [39,40,41,42]. In this study, two rounds of nested PCR were established to screen for T. muris infections in the digestive tracts of mice. Through sensitivity detection, the genomic DNA of at least 100 T. muris trophozoites/mL and 0.1 ng of genome DNA of mouse feces infected by T. muris per μL, which amounted to 176 trophozoites/g of feces, could be detected. Additionally, there were no cross-reactions with P. hominis, G. duodenalis, C. tyzzeri, T. vaginalis, or T. gallinae. The nested PCR improved the sensitivity of detecting T. muris by 26% over smear microscopy examination, and its sensitivity was significantly higher than that of smear microscopy examination. It is important for clinical applications to detect T. muris infection rates and prevalence. In addition, clinical studies have shown that using nested PCR to detect Trichomonas in bronchoalveolar lavage fluid significantly increases its sensitivity percentage compared to smear microscopy examination [43]. Moreover, Trichomonas vaginalis diagnosis was based on wet mount microscopy, which has poor sensitivity (60%) compared with PCR assays [44]. The results of this study indicated that nested PCR reactions have higher sensitivity and specificity. As a result of using the PCR method, Da Costa et al. [45] were able to detect the ITS gene of T. muris, and the PCR product length was 65 bp, which was too short to be confused with a dimer or primer. For this PCR method, high electrophoresis time and operation requirements were needed; otherwise, the electrophoresis gel would have easily escaped. However, the nested PCR product in this study was 384 bp, which was moderately sized and could avoid this problem. Additionally, nested PCR was more sensitive for detecting Trichomonas genes in clinical applications than PCR [46,47]. But Escalante NK et al. [4] used the qPCR method to detect 28 s rRNA genes of T. muris, which has the disadvantages of high cost and high instrument requirements, and many laboratories cannot obtain relatively expensive qPCR machines. One key disadvantage of existing qPCR techniques is their reliance on the reference standard curve, which makes comparing qPCR findings between labs challenging [48,49]. However, the nested PCR method used in this study to detect T. muris avoids the above issues in clinical application.
It is known that PCR is a tool of high sensitivity and specificity, particularly for identifying morphologically indistinguishable parasites such as species of Giardia and assemblages of Cryptosporidium, and for detecting their genetic variation [50]. Extensive genetic variation was observed within Giardia, of which there have been at least eight assemblages (A-H) identified in G. duodenalis. As a result of the genetic diversity of the Giardia population, this allows us to investigate Giardia duodenalis’ potential ecologically and evolutionarily [51]. Similarly, there is extensive genetic variation within the genus Cryptosporidium, with more than 120 genotypes of Cryptosporidium detected by analyzing the SSU rRNA gene [52]. Additionally, nested PCR is a considerably more specific and sensitive technique than PCR, so nested PCR was used to investigate the genetic variability within T. muris [50]. Based on the results of this study, one genotype of T. muris was identified, which indicates that it has a relatively wide distribution and stable transmission pattern. It will be necessary to conduct further research and exploration on other genotypes and the genetic variability of T. muris.
The commonly used homologous evolutionary analysis of conservative genes in intestinal protozoa mainly includes the 18S rRNA and ITS gene sequences. For example, the 18S gene and ITS gene were used to analyze the epidemic situation and homologous evolution of P. hominis [53,54]. SSU rRNA, as a conserved gene, has been widely used in phylogenetic comparisons; e.g., the SSU rRNA gene was used to perform a phylogenetic analysis among the genera of Trichomonas by comparing the base sequence length and secondary structure characteristics in T. foetus and T. vaginalis [22,23,24,55]. In addition, it was used to analyze the genetic characterization of Tritrichomonas gallinae (T. gallinae), Tritrichomonas tenax (T. tenax), and Tritrichomonas canistomae (T. canistomae), which were isolated from European turtle doves (Streptopelia turtur) and racing pigeons (Columba livia) [56]. PCR amplification and sequence analysis using SSU rRNA as a reference gene were helpful for investigating its prevalence in herbivores (cattle, sheep, and horses) in different regions of the world [57,58]. There are at least 44 effective species of Cryptosporidium and more than 120 genotypes of Cryptosporidium in humans and animals that have been detected by analyzing the SSU rRNA gene [52]. Furthermore, in this study, a novel primer for the detection of T. muris infections using nested PCR based on SSU rRNA was designed, which exhibited higher specificity and sensitivity when compared to the classical method of smear microscopy.
Cross-infection of intestinal parasites occurs in experimental mice, and, sometimes, it is difficult to distinguish between them using common smear microscopy. The most dramatic illustration of this is the phenomenon of a mixed infection of intestinal protozoa that exists in the intestinal tracts of laboratory mice. Laboratory mice can be infected with G. duodenalis and Cryptosporidium, which can parasitize their intestinal tracts [59,60]. T. muris and P. hominis are two kinds of flagellate parasites that can parasitize the ceca of rats, mice, and hamsters and have similar morphological characteristics. According to reports by Escalante et al. (2016) and Ghindilis et al. (2019) [4,17], T. muris showed morphological characteristics similar to those described in this study. It appears pear-shaped with an anterior vesicular nucleus. Three anterior flagella arise from the anterior region, as well as a posterior flagellum that is attached to its body by means of an undulating membrane that continues posteriorly as a free flagellum. It has a large oval-shaped nucleus, a slit-like cytostome, and a sausage-shaped parabasal body in the anterior region. P. hominis is piriform, measuring 8 to 20 by 3 to 14 μm, and ordinarily has five anterior flagella plus one posterior flagellum [53,54]. However, the nested PCR-specific primers designed in this study are particularly effective in distinguishing T. muris from other trichomonas due to their specificity.
In fact, it is still a worldwide problem that experimental mice’s digestive tracts are infested with T. muris, which is a serious threat to their health. For example, according to a study by Won et al. (2006) [8], laboratory mice in Korea had a 69% prevalence of T. muris infections. Surprisingly, the infection rate of Tritrichomonas sp. was up to 100% in different animal houses in Brazil [10]. Similarly, Japanese mice were found to be exclusively infected with T. muris, with a prevalence of infection as high as 67.8% in some areas [9]. A number of previous studies have shown that T. muris infections are largely caused by fecal contamination and pseudocyst transmission [1,2]. In view of these findings, it may be beneficial to prioritize the need to strengthen the management of experimental animal housing and detect animal health whenever possible [61]. Therefore, to prevent T. muris infections, animal breeding laws and regulations should be improved [62,63,64]. The following aspects should be improved: (i) Increasing awareness of the importance of clean and SPF animal production and the health and epidemic prevention of animal houses [62,65,66]. (ii) Mouse breeding environments should be clean and dust-free with fresh air, temperatures of 18–22 °C, relative humidities of 50–60%, and noise below 85 decibels [67]. In addition to cleaning and disinfecting rat houses, mouse boxes, water dispensers, bedding, and food, regular pest control is required [68]. (iii) Regular parasite detection should be performed in laboratory mice. (iv) A regular monitoring schedule should be maintained for intestinal parasites, such as P. hominis, G. duodenalis, Cryptosporidium sp., and T. muris.
In the present study, novel and specific SSU rRNA primer sets were designed for T. muris detection in laboratory mice. These results clearly demonstrate that nested PCR has higher sensitivity, reliability, and specificity. Thus, the present study provides further understanding of novel approaches for the detection of T. muris and a new means to ensure experimental animal health and welfare.
5. Conclusions
In conclusion, a nested PCR system with new primers has successfully been developed, which has higher specificity and sensitivity for the detection of T. muris infections in laboratory mice. Therefore, these findings should make an important contribution to the field and ensure the quality of laboratory mice.
Author Contributions
Writing—original draft, H.Z., X.W., X.Z. (Xu Zhang), J.L. and B.Y.; Formal Analysis, H.Z. and J.L.; Investigation, H.Z., N.Z. and P.G.; Conceptualization, X.Z. (Xichen Zhang); Project Administration, X.Z. (Xichen Zhang); Writing—Review, P.Z. and X.L.; Editing, F.G.; Validation, P.G.; Supervision: P.G. and X.Z. (Xichen Zhang); Funding acquisition, N.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the National Key Research and Development Program of China (No. 2021YFF0702900) and the National Natural Science Foundation of China (No. 32102696).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No data were used for the research described in this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mattern, C.F.; Daniel, W.A. Tritrichomonas muris in the Hamster: Pseudocysts and the infection of newborn. J. Protozool. 1980, 27, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Lipman, N.S.; Lampen, N.; Nguyen, H.T. Identification of pseudocysts of Tritrichomonas muris in Armenian hamsters and their transmission to mice. Lab. Anim. Sci. 1999, 49, 313–315. [Google Scholar] [PubMed]
- Maritz, J.M.; Land, K.M.; Carlton, J.M.; Hirt, R.P. What is the importance of zoonotic trichomonads for human health? Trends Parasitol. 2014, 30, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Escalante, N.K.; Lemire, P.; Cruz Tleugabulova, M.; Prescott, D.; Mortha, A.; Streutker, C.J.; Girardin, S.E.; Philpott, D.J.; Mallevaey, T. The common mouse protozoa Tritrichomonas muris alters mucosal T cell homeostasis and colitis susceptibility. J. Exp. Med. 2016, 213, 2841–2850. [Google Scholar] [CrossRef]
- Chang, C.Y.; Wang, J.; Zhao, Y.; Liu, J.; Yang, X.; Yue, X.; Wang, H.; Zhou, F.; Inclan-Rico, J.M.; Ponessa, J.J.; et al. Tumor suppressor p53 regulates intestinal type 2 immunity. Nat. Commun. 2021, 12, 3371. [Google Scholar] [CrossRef]
- Roach, P.D.; Wallis, P.M.; Olson, M.E. The use of Metronidazole, Tinidazole and Dimetridazole in eliminating Trichomonads from laboratory mice. Lab. Anim. 1988, 22, 361–364. [Google Scholar] [CrossRef]
- Cepicka, I.; Hampl, V.; Kulda, J.; Flegr, J. New evolutionary lineages, unexpected diversity, and host specificity in the parabasalid genus Tetratrichomonas. Mol. Phylogenet. Evol. 2006, 39, 542–551. [Google Scholar] [CrossRef]
- Won, Y.S.; Jeong, E.S.; Park, H.J.; Lee, C.H.; Nam, K.H.; Kim, H.C.; Hyun, B.H.; Lee, S.K.; Choi, Y.K. Microbiological contamination of laboratory mice and rats in Korea from 1999 to 2003. Exp. Anim. 2006, 55, 11–16. [Google Scholar] [CrossRef][Green Version]
- Hayashimoto, N.; Morita, H.; Ishida, T.; Uchida, R.; Tanaka, M.; Ozawa, M.; Yasuda, M.; Itoh, T. Microbiological survey of mice (Mus. musculus) purchased from commercial pet shops in Kanagawa and Tokyo, Japan. Exp. Anim. 2015, 64, 155–160. [Google Scholar] [CrossRef]
- Chagas, C.R.F.; Gonzalez, I.H.L.; Favoretto, S.M.; Ramos, P.L. Parasitological surveillance in a rat (Rattus norvegicus) colony in São Paulo Zoo animal house. Ann. Parasitol. 2017, 63, 291–297. [Google Scholar]
- Steenkeste, N.; Rogers, W.O.; Okell, L.; Jeanne, I.; Incardona, S.; Duval, L.; Chy, S.; Hewitt, S.; Chou, M.; Socheat, D.; et al. Sub-microscopic malaria cases and mixed malaria infection in a remote area of high malaria endemicity in Rattanakiri province, Cambodia: Implication for malaria elimination. Malar. J. 2010, 9, 108. [Google Scholar] [CrossRef]
- Daniels, M.E.; Smith, W.A.; Jenkins, M.W. Estimating Cryptosporidium and Giardia disease burdens for children drinking untreated groundwater in a rural population in India. PLoS Negl. Trop. Dis. 2018, 12, e0006231. [Google Scholar] [CrossRef]
- Hess, J. Review of current methods for the detection of Trichomonas in clinical material. J. Clin. Pathol. 1969, 22, 269–272. [Google Scholar] [CrossRef]
- Inoue, T.; Hayashimoto, N.; Yasuda, M.; Sasaki, E.; Itoh, T. Pentatrichomonas hominis in laboratory-bred common marmosets. Exp. Anim. 2015, 64, 363–368. [Google Scholar] [CrossRef]
- Abou Gamra, M.M.; Kamel, H.H.; Ezz Eldin, H.M.; Taha Zahran, F.M. Efficacy of netropsin dihydrochloride against the viability, cytopathogenicity and hemolytic activity of Trichomonas vaginalis clinical isolates. J. Infect. Chemother. 2019, 25, 955–964. [Google Scholar] [CrossRef]
- Chudnovskiy, A.; Mortha, A.; Kana, V.; Kennard, A.; Ramirez, J.D.; Rahman, A.; Remark, R.; Mogno, I.; Ng, R.; Gnjatic, S.; et al. Host-protozoan interactions protect from mucosal infections through activation of the inflammasome. Cell 2016, 167, 444–456.e14. [Google Scholar] [CrossRef]
- Ghindilis, A.L.; Chesnokov, O.; Ngasala, B.; Smith, M.W.; Smith, K.; Mårtensson, A.; Oleinikov, A.V. Detection of sub-microscopic blood levels of Plasmodium falciparum using Tandem Oligonucleotide Repeat Cascade Amplification (TORCA) assay with an attomolar detection limit. Sci. Rep. 2019, 9, 2901. [Google Scholar] [CrossRef]
- Van de Peer, Y.; De Rijk, P.; Wuyts, J.; Winkelmans, T.; De Wachter, R. The European small subunit ribosomal RNA database. Nucleic Acids Res. 2000, 28, 175–176. [Google Scholar] [CrossRef]
- Olsen, G.J.; Woese, C.R. Ribosomal RNA: A key to phylogeny. FASEB J. 1993, 7, 113–123. [Google Scholar] [CrossRef]
- Van de Peer, Y.; De Wachter, R. Evolutionary relationships among the eukaryotic crown taxa taking into account site-to-site rate variation in 18S rRNA. J. Mol. Evol. 1997, 45, 619–630. [Google Scholar] [CrossRef]
- Yu, F.; Li, D.; Chang, Y.; Wu, Y.; Guo, Z.; Jia, L.; Xu, J.; Li, J.; Qi, M.; Wang, R.; et al. Molecular characterization of three intestinal protozoans in hospitalized children with different disease backgrounds in Zhengzhou, central China. Parasites Vectors 2019, 12, 543. [Google Scholar] [CrossRef]
- Chakrabarti, D.; Dame, J.B.; Gutell, R.R.; Yowell, C.A. Characterization of the rDNA unit and sequence analysis of the small subunit rRNA and 5.8S rRNA genes from Tritrichomonas foetus. Mol. Biochem. Parasitol. 1992, 52, 75–83. [Google Scholar] [CrossRef]
- Edgcomb, V.; Viscogliosi, E.; Simpson, A.G.; Delgado-Viscogliosi, P.; Roger, A.J.; Sogin, M.L. New insights into the phylogeny of Trichomonads inferred from small subunit rRNA sequences. Protist 1998, 149, 359–366. [Google Scholar] [CrossRef]
- Lumsden, T.; Bentley, A.A.; Beutler, W.; Ghosh, A.; Galkin, O.; Komar, A.A. Yeast strains with N-terminally truncated ribosomal protein S5: Implications for the evolution, structure and function of the Rps5/Rps7 proteins. Nucleic Acids Res. 2010, 38, 1261–1272. [Google Scholar] [CrossRef]
- Hawley, A.K.; Torres-Beltrán, M.; Zaikova, E.; Walsh, D.A.; Mueller, A.; Scofield, M.; Kheirandish, S.; Payne, C.; Pakhomova, L.; Bhatia, M.; et al. A compendium of multi-omic sequence information from the Saanich Inlet water column. Sci. Data 2017, 4, 170160. [Google Scholar] [CrossRef]
- Ajonina, C.; Buzie, C.; Möller, J.; Otterpohl, R. The detection of Entamoeba histolytica and Toxoplasma gondii in wastewater. J. Toxicol. Environ. Health A 2018, 81, 1–5. [Google Scholar] [CrossRef]
- Li, W.C.; Wang, K.; Li, Y.; Zhao, L.P.; Xiao, Y.; Gu, Y.F. Survey and molecular characterization of Trichomonads in pigs in Anhui Province, East China, 2014. Iran. J. Parasitol. 2018, 13, 602–610. [Google Scholar]
- Vejano, M.R.A.; Dela Peña, L.B.R.O.; Rivera, W.L. Occurrence of Giardia duodenalis in selected stations and tributary rivers of Laguna Lake, Philippines. Environ. Monit. Assess. 2021, 193, 466. [Google Scholar] [CrossRef]
- Keller, M.; Naue, J.; Zengerle, R.; von Stetten, F.; Schmidt, U. Automated forensic animal family identification by nested PCR and melt curve analysis on an Off-the-Shelf thermocycler augmented with a centrifugal microfluidic disk segment. PLoS ONE 2015, 10, e0131845. [Google Scholar] [CrossRef]
- Saeki, H.; Togo, M.; Imai, S.; Ishii, T. A new method for the serial cultivation of Tritrichomonas muris. Nihon Juigaku Zasshi. 1983, 45, 151–156. [Google Scholar] [CrossRef]
- Howitt, M.R.; Lavoie, S.; Michaud, M.; Blum, A.M.; Tran, S.V.; Weinstock, J.V.; Gallini, C.A.; Redding, K.; Margolskee, R.F.; Osborne, L.C.; et al. Tuft cells, taste-chemosensory cells, orchestrate parasite type 2 immunity in the gut. Science 2016, 351, 1329–1333. [Google Scholar] [CrossRef] [PubMed]
- Riggio, M.P.; Lennon, A.; Wray, D. Detection of Helicobacter pylori DNA in recurrent aphthous stomatitis tissue by PCR. J. Oral Pathol. Med. 2000, 29, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Riggio, M.P.; Lennon, A. Development of a novel PCR assay for detection of Prevotella oris in clinical specimens. FEMS Microbiol. Lett. 2007, 276, 123–128. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Steingart, K.R.; Henry, M.; Ng, V.; Hopewell, P.C.; Ramsay, A.; Cunningham, J.; Urbanczik, R.; Perkins, M.; Aziz, M.A.; Pai, M. Fluorescence versus conventional sputum smear microscopy for tuberculosis: A systematic review. Lancet Infect. Dis. 2006, 6, 570–581. [Google Scholar] [CrossRef]
- Namagembe, A.; Ssekabira, U.; Weaver, M.R.; Blum, N.; Burnett, S.; Dorsey, G.; Sebuyira, L.M.; Ojaku, A.; Schneider, G.; Willis, K.; et al. Improved clinical and laboratory skills after team-based, malaria case management training of health care professionals in Uganda. Malar. J. 2012, 11, 44. [Google Scholar] [CrossRef]
- Bjerrum, S.; Schiller, I.; Dendukuri, N.; Kohli, M.; Nathavitharana, R.R.; Zwerling, A.A.; Denkinger, C.M.; Steingart, K.R.; Shah, M. Lateral flow urine lipoarabinomannan assay for detecting active tuberculosis in people living with HIV. Cochrane Database Syst. Rev. 2019, 10, CD011420. [Google Scholar] [CrossRef]
- Ren, N.; JinLi, J.; Chen, Y.; Zhou, X.; Wang, J.; Ge, P.; Khan, F.A.; Zhang, L.; Hu, C.; Robertson, I.D.; et al. Identification of new diagnostic biomarkers for Mycobacterium tuberculosis and the potential application in the serodiagnosis of human tuberculosis. Microb. Biotechnol. 2018, 11, 893–904. [Google Scholar] [CrossRef]
- Lam, W.Y.; Yeung, A.C.; Tang, J.W.; Ip, M.; Chan, E.W.; Hui, M.; Chan, P.K. Rapid multiplex nested PCR for detection of respiratory viruses. J. Clin. Microbiol. 2007, 45, 3631–3640. [Google Scholar] [CrossRef]
- Osaki, S.C.; Soccol, V.T.; Costa, A.O.; Oliveira-Silva, M.B.; Pereira, J.T.; Procópio, A.E. Polymerase chain reaction and nested-PCR approaches for detecting Cryptosporidium in water catchments of water treatment plants in Curitiba, State of Paraná, Brazil. Rev. Soc. Bras. Med. Trop. 2013, 46, 270–276. [Google Scholar] [CrossRef]
- Šeligová, B.; Lukáč, Ľ.; Bábelová, M.; Vávrová, S.; Sulo, P. Diagnostic reliability of nested PCR depends on the primer design and threshold abundance of Helicobacter pylori in biopsy, stool, and saliva samples. Helicobacter 2020, 25, e12680. [Google Scholar] [CrossRef] [PubMed]
- Smolejová, M.; Cihová, I.; Sulo, P. Reliable and sensitive nested PCR for the detection of chlamydia in sputum. Microorganisms 2021, 9, 935. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Ying, F.; Lai, Y.; Li, X.; Xue, X.; Zhou, T.; Hu, D. Use of nested PCR for the detection of Trichomonads in bronchoalveolar lavage fluid. BMC Infect. Dis. 2019, 19, 512. [Google Scholar] [CrossRef] [PubMed]
- Overbaugh, J.; McClelland, R.S.; John-Stewart, G. HIV acquisition during pregnancy and postpartum is associated with genital infections and partnership characteristics. AIDS 2015, 29, 2025–2033. [Google Scholar]
- Da Costa, A.; Graham, T.; Duncan, J.; Pillai, S.; Jennifer, L. Detection and treatment strategy for Tritrichomonas muris in the common laboratory mouse. bioRxiv 2019, 11, 827055. [Google Scholar] [CrossRef]
- Aguirre, E.; Tesouro, M.; Amusategui, I.; Rodríguez-Franco, F.; Sainz, A. Comparison between different polymerase chain reaction methods for the diagnosis of Ehrlichia canis infection. Ann. N. Y. Acad. Sci. 2008, 1149, 118–120. [Google Scholar] [CrossRef]
- Noh, C.S.; Kim, S.S.; Park, S.Y.; Moon, H.S.; Hong, Y.; Ryu, J.S. Comparison of Two PCR Assays for Trichomonas vaginalis. Korean J. Parasitol. 2019, 57, 27–31. [Google Scholar] [CrossRef]
- Dandasena, D.; Bhandari, V.; Sreenivasamurthy, G.S.; Murthy, S.; Roy, S.; Bhanot, V.; Arora, J.S.; Singh, S.; Sharma, P. A Real-Time PCR based assay for determining parasite to host ratio and parasitaemia in the clinical samples of Bovine Theileriosis. Sci. Rep. 2018, 8, 15441. [Google Scholar] [CrossRef]
- Murthy, S.; Suresh, A.; Dandasena, D.; Singh, S.; Subudhi, M.; Bhandari, V.; Bhanot, V.; Arora, J.S.; Sharma, P. Multiplex ddPCR: A promising diagnostic assay for early detection and drug monitoring in Bovine Theileriosis. Pathogens 2023, 12, 296. [Google Scholar] [CrossRef]
- Fayer, R.; Santin, M.; Macarisin, D. Detection of concurrent infection of dairy cattle with Blastocystis, Cryptosporidium, Giardia, and Enterocytozoon by molecular and microscopic methods. Parasitol. Res. 2012, 111, 1349–1355. [Google Scholar] [CrossRef]
- Belkessa, S.; Thomas-Lopez, D.; Houali, K.; Ghalmi, F.; Stensvold, C.R. Molecular characterization of Giardia duodenalis in children and adults sampled in Algeria. Microorganisms 2020, 9, 54. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Zahedi, A.; Feng, Y.; Xiao, L. An update on zoonotic Cryptosporidium species and genotypes in Humans. Animals 2021, 11, 3307. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhang, H.; Yu, Y.; Gong, P.; Li, J.; Li, Z.; Li, T.; Cong, Z.; Tian, C.; Liu, X.; et al. High prevalence of Pentatrichomonas hominis infection in gastrointestinal cancer patients. Parasites Vectors 2019, 12, 423. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, N.; Gong, P.; Cheng, S.; Wang, X.; Li, X.; Hou, Z.; Liu, C.; Bi, T.; Wang, B.; et al. Prevalence and molecular characterization of Pentatrichomonas hominis in Siberian tigers (Panthera tigris altaica) in northeast China. Integr. Zool. 2022, 17, 543–549. [Google Scholar] [CrossRef]
- Herath, S.; Balendran, T.; Herath, A.; Iddawela, D.; Wickramasinghe, S. Comparison of diagnostic methods and analysis of socio-demographic factors associated with Trichomonas vaginalis infection in Sri Lanka. PLoS ONE 2021, 16, e0258556. [Google Scholar] [CrossRef]
- Martínez-Herrero, M.C.; Garijo-Toledo, M.M.; Liebhart, D.; Ganas, P.; Martínez-Díaz, R.A.; Ponce-Gordo, F.; Carrero-Ruiz, A.; Hess, M.; Gómez-Muñoz, M.T. Novel avian oropharyngeal trichomonads isolated from European turtle doves (Streptopelia turtur) and racing pigeons (Columba livia): Genetic and morphometric characterisation of clonal cultures. Infect. Genet. Evol. 2017, 55, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Santín, M.; Cortés Vecino, J.A.; Fayer, R. A large scale molecular study of Giardia duodenalis in horses from Colombia. Vet. Parasitol. 2013, 196, 31–36. [Google Scholar] [CrossRef]
- Jian, Y.; Zhang, X.; Li, X.; Karanis, G.; Ma, L.; Karanis, P. Prevalence and molecular characterization of Giardia duodenalis in cattle and sheep from the Qinghai-Tibetan Plateau Area (QTPA), northwestern China. Vet. Parasitol. 2018, 250, 40–44. [Google Scholar] [CrossRef]
- Fan, Y.; Wang, X.; Yang, R.; Zhao, W.; Li, N.; Guo, Y.; Xiao, L.; Feng, Y. Molecular characterization of the waterborne pathogens Cryptosporidium spp., Giardia duodenalis, Enterocytozoon bieneusi, Cyclospora cayetanensis and Eimeria spp. in wastewater and sewage in Guangzhou, China. Parasites Vectors 2021, 14, 66. [Google Scholar] [CrossRef]
- Xu, J.; Liu, H.; Jiang, Y.; Jing, H.; Cao, J.; Yin, J.; Li, T.; Sun, Y.; Shen, Y.; Wang, X. Genotyping and subtyping of Cryptosporidium spp. and Giardia duodenalis isolates from two wild rodent species in Gansu Province, China. Sci. Rep. 2022, 12, 12178. [Google Scholar] [CrossRef]
- Zhang, H.; Shan, F.; Zhou, X.; Li, B.; Zhai, J.Q.; Zou, S.Z.; Wu, M.F.; Chen, W.; Zhai, S.L.; Luo, M.L. Outbreak and genotyping of canine distemper virus in captive Siberian tigers and red pandas. Sci. Rep. 2017, 7, 8132. [Google Scholar] [CrossRef] [PubMed]
- Loew, F.M. The animal welfare bête noire in veterinary medicine. Can. Vet. J. 1987, 28, 689–692. [Google Scholar] [PubMed]
- Marx, J.O.; Jacobsen, K.O.; Petervary, N.A.; Casebolt, D.B. A survey of laboratory animal veterinarians regarding mouse welfare in biomedical research. J. Am. Assoc. Lab. Anim. Sci. 2021, 60, 139–145. [Google Scholar] [CrossRef]
- Korbonits, L.; Kleinwort, K.J.H.; Amann, B.; Didier, A.; Märtlbauer, E.; Hauck, S.M.; Deeg, C.A. Mycobacterium avium subsp. paratuberculosis infected cows reveal divergent immune response in bovine peripheral blood derived lymphocyte proteome. Metabolites 2022, 12, 924. [Google Scholar] [CrossRef]
- Whiting, T.L.; Brennan, S.C.; Wruck, G.C. The veterinary profession’s role in policing animal welfare. Can. Vet. J. 2006, 47, 1065–1072. [Google Scholar]
- Thodberg, K.; Gould, L.M.; Støier, S.; Anneberg, I.; Thomsen, P.T.; Herskin, M.S. Experiences and opinions of Danish livestock drivers transporting sows regarding fitness for transport and management choices relevant for animal welfare. Transl. Anim. Sci. 2020, 4, txaa015. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, S.; Lei, Z.; Chen, G.; Huang, L.; Yang, F.; Lei, Y.; Liu, Y.; Yang, L.; Liu, W.; et al. Circular RNA profile in liver tissue of EpCAM knockout mice. Int. J. Mol. Med. 2019, 44, 1063–1077. [Google Scholar] [CrossRef]
- Vonesch, N.; Binazzi, A.; Bonafede, M.; Melis, P.; Ruggieri, A.; Iavicoli, S.; Tomao, P. Emerging zoonotic viral infections of occupational health importance. Pathog. Dis. 2019, 77, ftz018. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).