Blood-Based Markers for Skeletal and Cardiac Muscle Function in Eventing Horses before and after Cross-Country Rides and How They Are Influenced by Plasma Volume Shift

 , , , ,
, , , , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.1.1. Horses
2.1.2. Riders
2.1.3. Training Schedules
2.1.4. Exercise
2.1.5. Sampling Times
2.2. Sample Collection
2.2.1. Sample Storage and Transport to the Testing Laboratory
2.2.2. Blood Parameters
2.2.3. Measurement Techniques
2.3. Missing Values
2.4. Data Analysis
2.4.1. Plasma Volume Loss Adjustment
2.4.2. Mixed Model
3. Results
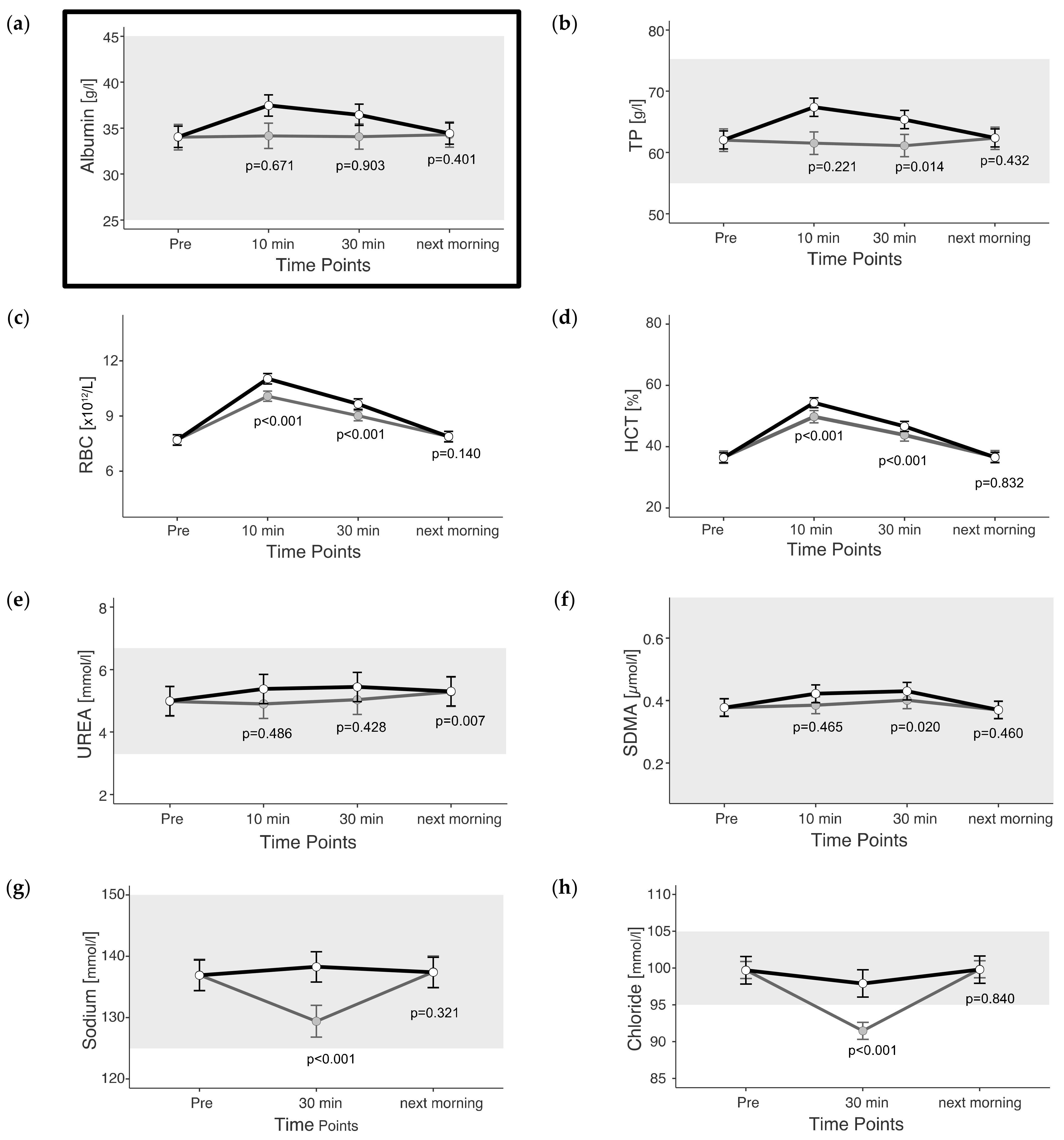
3.1. Fluid-Balance-Related Parameters
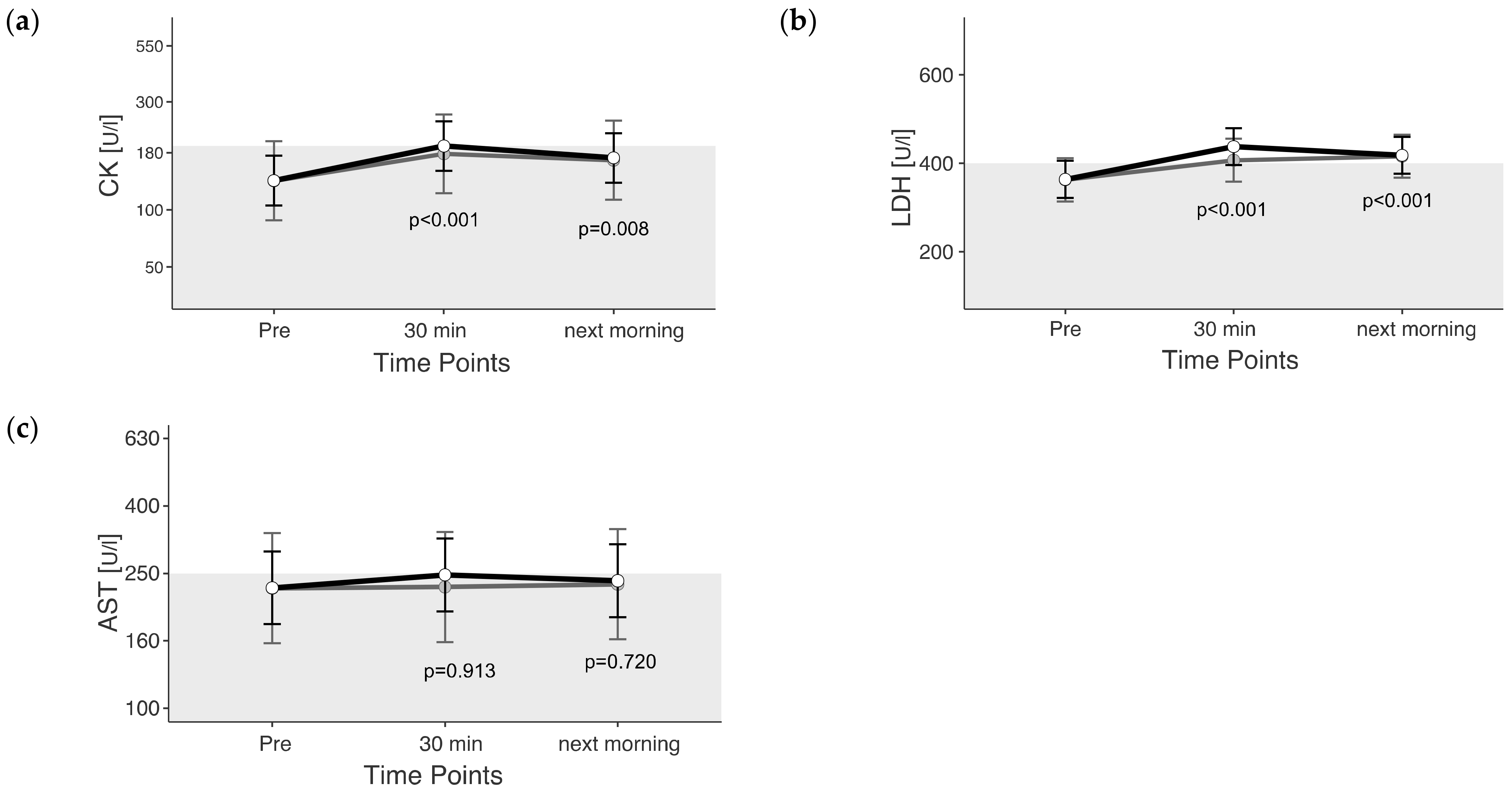
3.2. Muscle Enzymes
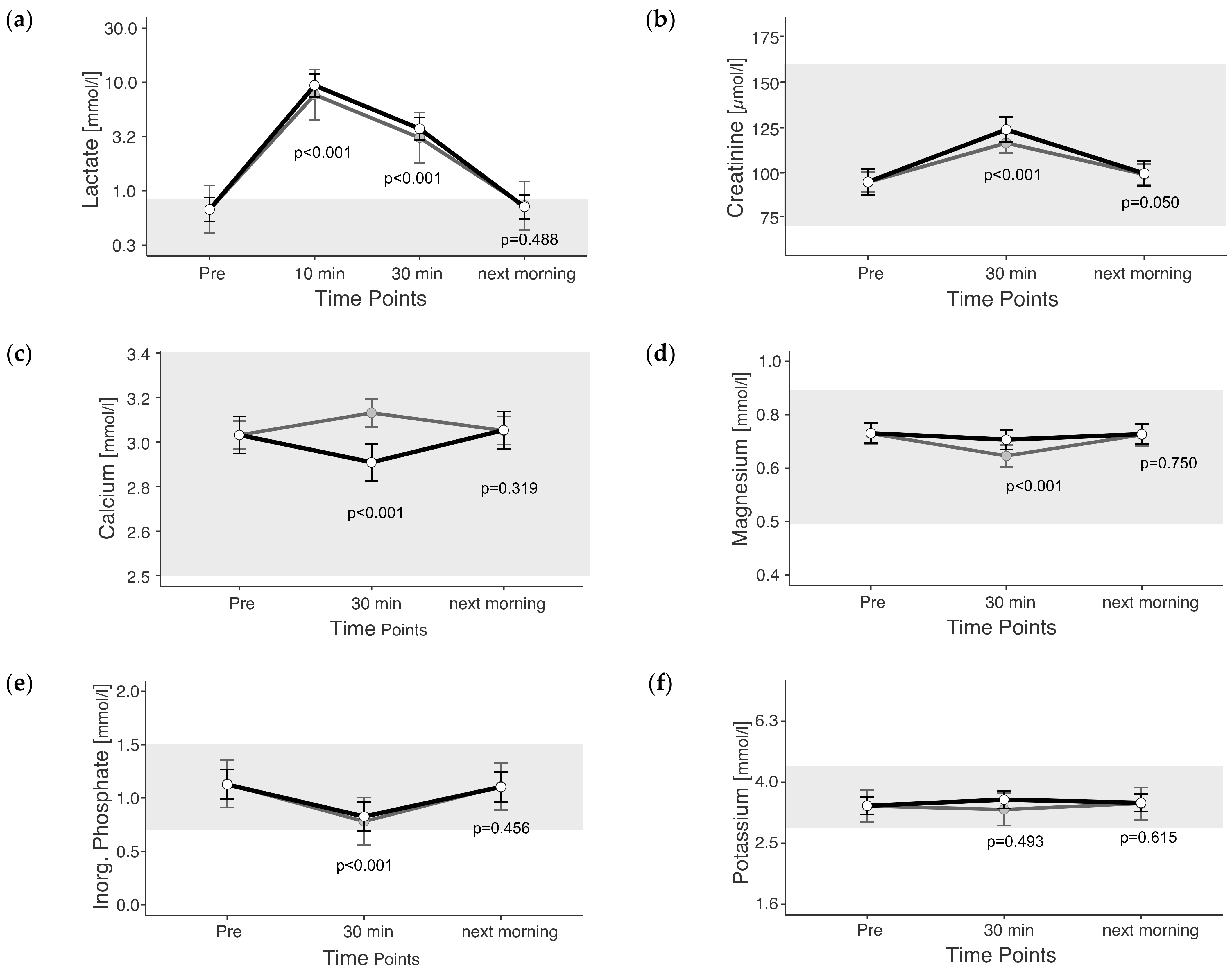
3.3. Muscle Metabolites
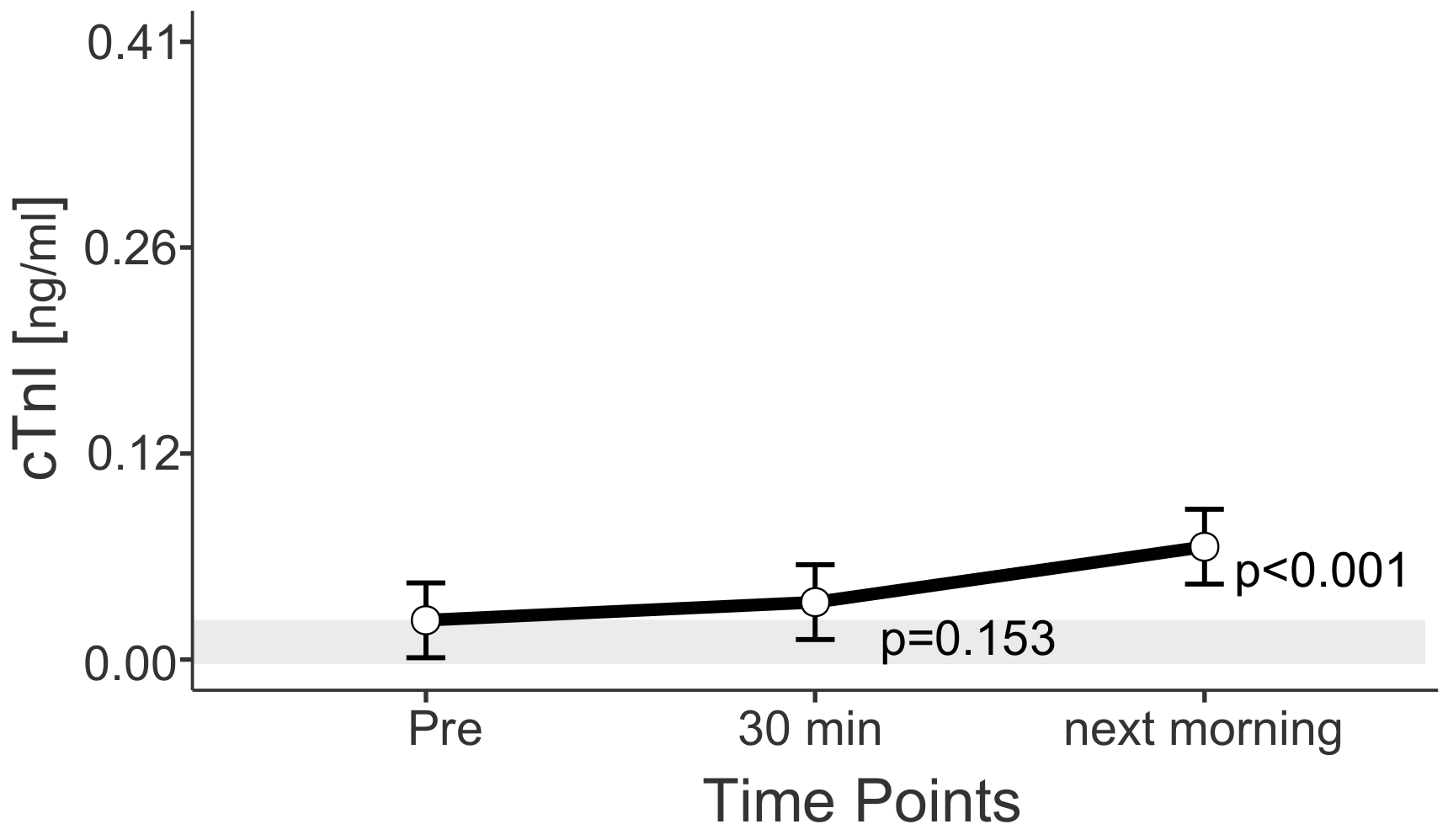
3.4. Cardiac Markers
4. Discussion
4.1. Fluid Balance
4.2. Muscle Enzymes
4.3. Muscle Metabolites
4.4. Cardiac Markers
4.5. Muscle and Cardiac Fatigue
4.6. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hecksteden, A.; Kraushaar, J.; Scharhag-Rosenberger, F.; Theisen, D.; Senn, S.; Meyer, T. Individual response to exercise training—A statistical perspective. J. Appl. Physiol. 2015, 118, 1450–1459. [Google Scholar] [CrossRef] [PubMed]
- Hecksteden, A.; Pitsch, W.; Julian, R.; Pfeiffer, M.; Kellmann, M.; Ferrauti, A.; Meyer, T. A New Method to Individualize Monitoring of Muscle Recovery in Athletes. Int. J. Sports Physiol. Perform. 2017, 12, 1137–1142. [Google Scholar] [CrossRef] [PubMed]
- Julian, R.; Meyer, T.; Fullagar, H.H.; Skorski, S.; Pfeiffer, M.; Kellmann, M.; Ferrauti, A.; Hecksteden, A. Individual Patterns in Blood-Borne Indicators of Fatigue-Trait or Chance. J. Strength Cond. Res. 2017, 31, 608–619. [Google Scholar] [CrossRef]
- Hecksteden, A.; Meyer, T. Personalized Sports Medicine—Principles and tailored implementations in preventive and competitive sports. Dtsch. Z. Sportmed. 2018, 2018, 73–80. [Google Scholar] [CrossRef]
- Hecksteden, A.; Pitsch, W.; Rosenberger, F.; Meyer, T. Repeated testing for the assessment of individual response to exercise training. J. Appl. Physiol. 2018, 124, 1567–1579. [Google Scholar] [CrossRef] [PubMed]
- Barth, V.; Käsbauer, H.; Ferrauti, A.; Kellmann, M.; Pfeiffer, M.; Hecksteden, A.; Meyer, T. Individualized Monitoring of Muscle Recovery in Elite Badminton. Front. Physiol. 2019, 10, 778. [Google Scholar] [CrossRef]
- Hacker, S.; Reichel, T.; Hecksteden, A.; Weyh, C.; Gebhardt, K.; Pfeiffer, M.; Ferrauti, A.; Kellmann, M.; Meyer, T.; Krüger, K. Recovery-Stress Response of Blood-Based Biomarkers. Int. J. Environ. Res. Public Health 2021, 18, 5776. [Google Scholar] [CrossRef]
- Kellmann, M.; Bertollo, M.; Bosquet, L.; Brink, M.; Coutts, A.J.; Duffield, R.; Erlacher, D.; Halson, S.L.; Hecksteden, A.; Heidari, J.; et al. Recovery and Performance in Sport: Consensus Statement. Int. J. Sports Physiol. Perform. 2018, 13, 240–245. [Google Scholar] [CrossRef]
- Delsmann, M.M.; Stürznickel, J.; Amling, M.; Ueblacker, P.; Rolvien, T. Musculoskeletal laboratory diagnostics in competitive sport. Orthopade 2021, 50, 700–712. [Google Scholar] [CrossRef]
- Plisak, U.; Szczepaniak, J.; Żmigrodzka, M.; Giercuszkiewicz-Hecold, B.; Witkowska-Piłaszewicz, O. Changes in novel anti-infalmmatory cytokine concetration in the bood of endurance and race horses at different levels of training. Comput. Struct. Biotechnol. J. 2023, 21, 418–424. [Google Scholar] [CrossRef]
- Kirsch, K.; Sandersen, C. Traditional and quantitative analysis of acid-base and electrolyte imbalances in horses competing in cross-country competitions at 2-star to 5-star level. J. Vet. Intern. Med. 2020, 34, 909–921. [Google Scholar] [CrossRef]
- Brehm, W.; Gehlen, H.; Ohnesorge, B.; Wehrend, A.; Dietz, O.; Huskamp, B.; Bartmann, C.P. (Eds.) Handbuch Pferdepraxis. 4; Vollständig Überarbeitete und Erweiterte Auflage ed.; 1239 Seiten; Enke: Stuttgart, Germany, 2017. [Google Scholar]
- Williams, C.A.; Burk, A.O. Antioxidant status in elite three-day event horses during competition. Oxid. Med. Cell. Longev. 2012, 2012, 572090. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lejeune, J.P.; Sandersen, C.; Votion, D.; Caudron, I.; Vander Heyden, L.; Franck, T.; Ceusters, J.; Mouithys-Mickalad, A.; Niesten, A.; De La Rebière de Pouyade, G.; et al. Effect of intensive exercise on plasmatic neutrophil elastase level in eventing and endurance horses. Equine Vet. J. Suppl. 2010, 42, 12–16. [Google Scholar] [CrossRef]
- Art, T.; Franck, T.; Gangl, M.; Votion, D.; Kohnen, S.; Deby-Dupont, G.; Serteyn, D. Plasma concentrations of myeloperoxidase in endurance and 3-day event horses after a competition. Equine Vet. J. Suppl. 2006, 38, 298–302. [Google Scholar] [CrossRef]
- Marlin, D.J.; Harris, P.A.; Schroter, R.C.; Harris, R.C.; Roberts, C.A.; Scott, C.M.; Orme, C.E.; Dunnett, M.; Dyson, S.J.; Barrelet, F. Physiological, metabolic and biochemical responses of horses competing in the speed and endurance phase of a CCI*****3-day-event. Equine Vet. J. Suppl. 1995, 27, 37–46. [Google Scholar] [CrossRef]
- Andrews, F.M.; Geiser, D.R.; White, S.L.; Williamson, L.H.; Maykuth, P.L.; Green, E.M. Haematological and biochemical changes in horses competing in a 3 Star horse trial and 3-day-event. Equine Vet. J. Suppl. 1995, 27, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Mair, J.; Jaffe, A.; Apple, F.; Lindahl, B. Cardiac Biomarkers. Dis. Markers 2015, 2015, 370569. [Google Scholar] [CrossRef] [PubMed]
- Lippi, G.; Sanchis-Gomar, F. Cardiac troponins in diagnostics of equine myocardial injury. J. Lab. Precis. Med. 2020, 5, 31. [Google Scholar] [CrossRef]
- Alpert, J.S.; Thygesen, K.; Antman, E.; Bassand, J.P. Myocardial infarction redefined—A consensus document of The Joint European Society of Cardiology/American College of Cardiology Committee for the redefinition of myocardial infarction. J. Am. Coll. Cardiol. 2000, 36, 959–969. [Google Scholar] [CrossRef]
- Baker, P.; Leckie, T.; Harrington, D.; Richardson, A. Exercise-induced cardiac troponin elevation: An update on the evidence, mechanism and implications. Int. J. Cardiol. Heart Vasc. 2019, 22, 181–186. [Google Scholar] [CrossRef]
- O’Brien, P.J.; Smith, D.E.; Knechtel, T.J.; Marchak, M.A.; Pruimboom-Brees, I.; Brees, D.J.; Spratt, D.P.; Archer, F.J.; Butler, P.; Potter, A.N.; et al. Cardiac troponin I is a sensitive, specific biomarker of cardiac injury in laboratory animals. Lab. Anim. 2006, 40, 153–171. [Google Scholar] [CrossRef]
- Phillips, W.; Giguère, S.; Franklin, R.P.; Hernandez, J.; Adin, D.; Peloso, J.G. Cardiac troponin I in pastured and race-training Thoroughbred horses. J. Vet. Intern. Med. 2003, 17, 597–599. [Google Scholar] [CrossRef]
- Tjora, S.; Gjestland, H.; Mordal, S.; Agewall, S. Troponin rise in healthy subjects during exercise test. Int. J. Cardiol. 2011, 151, 375–376. [Google Scholar] [CrossRef]
- Gehlen, H.; Rohn, K.; Deegen, E.; Stadler, P. Analysis of laboratory values in horses with cardiac disease: Diagnostic value of different cardiac biomarkers. Pferdeheilkunde 2006, 22, 532–541. [Google Scholar] [CrossRef]
- Holbrook, T.C.; Birks, E.K.; Sleeper, M.M.; Durando, M. Endurance exercise is associated with increased plasma cardiac troponin I in horses. Equine Vet. J. Suppl. 2006, 38, 27–31. [Google Scholar] [CrossRef]
- Flethøj, M.; Kanters, J.K.; Haugaard, M.M.; Pedersen, P.J.; Carstensen, H.; Balling, J.D.; Olsen, L.H.; Buhl, R. Changes in heart rate, arrhythmia frequency, and cardiac biomarker values in horses during recovery after a long-distance endurance ride. J. Am. Vet. Med. Assoc. 2016, 248, 1034–1042. [Google Scholar] [CrossRef]
- Ertelt, A.; Merle, R.; Stumpff, F.; Bollinger, L.; Liertz, S.; Weber, C.; Gehlen, H. Evaluation of Different Blood Parameters From Endurance Horses Competing at 160 km. J. Equine Vet. Sci. 2021, 104, 103687. [Google Scholar] [CrossRef]
- Fazio, F.; Aragona, F.; Piccione, G.; Pino, C.; Giannetto, C. Cardiac Biomarker Responses to Acute Exercise in Show Jumping Horses. J. Equine Vet. Sci. 2023, 128, 104882. [Google Scholar] [CrossRef] [PubMed]
- Ayvazoglu, C.; Kiziltpe, S.; Yaşar, Ü.; Yaşar, Z.; Demir, P.; Tunc, A. Changes in cardiac troponin I (cTnI), T (cTnT), and some biochemical parameters in Arabian racehorses after training. S. Afr. J. Anim. Sci. 2023, 53, 1–6. [Google Scholar] [CrossRef]
- Pourmohammad, R.; Mohri, M.; Seifi, H.A.; Sardari, K. Evaluation of cardiac troponin I, atrial natriuretic peptide and some oxidative/antioxidative biomarkers in the serum and hemolysate of trained Arabian horses after exercise. Iran. J. Vet. Res. 2020, 21, 211–215. [Google Scholar] [PubMed]
- Rossi, T.M.; Kavsak, P.A.; Maxie, M.G.; Pearl, D.L.; Pyle, W.G.; Physick-Sheard, P.W. Effect of racing on cardiac troponin I concentration and associations with cardiac rhythm disturbances in Standardbred racehorses. J. Vet. Cardiol. 2021, 35, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Gunther-Harrington, C.T.; Arthur, R.; Estell, K.; Martinez Lopez, B.; Sinnott, A.; Ontiveros, E.; Varga, A.; Stern, J.A. Prospective pre- and post-race evaluation of biochemical, electrophysiologic, and echocardiographic indices in 30 racing thoroughbred horses that received furosemide. BMC Vet. Res. 2018, 14, 18. [Google Scholar] [CrossRef] [PubMed]
- Fédération Equestre Internationale. FEI Database. Available online: https://data.fei.org/Horse/Search.aspx (accessed on 11 November 2022).
- Fédération Equestre Internationale. Available online: https://inside.fei.org/fei/disc/eventing/rules (accessed on 11 November 2022).
- The Jamovi Project (2022) Jamovi (Version 2.3) [Computer Software]. Available online: https://www.jamovi.org (accessed on 5 December 2022).
- Witkowska-Pilaszewicz, O.; Bąska, P.; Czopowicz, M.; Żmigrodzka, M.; Szczepaniak, J.; Szarska, E.; Winnicka, A.; Cywińska, A. Changes in serum amyloid a (SAA) concentration in Arabian endurance horses during first training season. Animals 2019, 9, 330. [Google Scholar] [CrossRef]
- Masri, M.; Freestone, J.F.; Wolfsheimer, K.J.; Shoemaker, K. Alterations in plasma volume, plasma constituents, renin activity and aldosterone induced by maximal exercise in the horse. Equine Vet. J. Suppl. 1990, 22, 72–77. [Google Scholar] [CrossRef]
- EclinPath. Available online: https://eclinpath.com/ (accessed on 25 May 2023).
- Assunção, P.; Barbosa, T.; Yonezawa, L.; Barbosa, L.; Watanabe, M.; Kohayagawa, A.; Schmidt, E. Acute-phase protein profile in horses subjected to different exercise protocols. Can. J. Vet. Res. 2019, 83, 272–278. [Google Scholar]
- McKeever, K.H.; Hinchcliff, K.W.; Reed, S.M.; Robertson, J.T. Plasma constituents during incremental treadmill exercise in intact and splenectomised horses. Equine Vet. J. 1993, 25, 233–236. [Google Scholar] [CrossRef]
- McKeever, K.H.; Hinchcliff, K.W.; Reed, S.M.; Robertson, J.T. Role of decreased plasma volume in hematocrit alterations during incremental treadmill exercise in horses. Am. J. Physiol. 1993, 265 Pt 2, R404–R408. [Google Scholar] [CrossRef] [PubMed]
- Flethøj, M.; Schwarzwald, C.C.; Haugaard, M.M.; Carstensen, H.; Kanters, J.K.; Olsen, L.H.; Buhl, R. Left Ventricular Function After Prolonged Exercise in Equine Endurance Athletes. J. Vet. Intern. Med. 2016, 30, 1260–1269. [Google Scholar] [CrossRef]
- Robert, C.; Goachet, A.G.; Fraipont, A.; Votion, D.M.; Van Erck, E.; Leclerc, J.L. Hydration and electrolyte balance in horses during an endurance season. Equine Vet. J. Suppl. 2010, 42, 98–104. [Google Scholar] [CrossRef]
- Muñoz, A.; Riber, C.; Trigo, P.; Castejón-Riber, C.; Castejón, F.M. Dehydration, electrolyte imbalances and renin-angiotensin-aldosterone-vasopressin axis in successful and unsuccessful endurance horses. Equine Vet. J. Suppl. 2010, 42, 83–90. [Google Scholar] [CrossRef]
- Riccioni, G.; Scotti, L.; Guagnano, M.T.; Bosco, G.; Bucciarelli, V.; Di Ilio, E.; Speranza, L.; Martini, F.; Bucciarelli, T. Physical exercise reduces synthesis of ADMA, SDMA, and L-Arg. Front. Biosci. Elite Ed. 2015, 7, 417–422. [Google Scholar] [CrossRef]
- Nyborg, C.; Bonnevie-Svendsen, M.; Melsom, H.S.; Melau, J.; Seljeflot, I.; Hisdal, J. Reduced L-Arginine and L-Arginine-ADMA-Ratio, and Increased SDMA after Norseman Xtreme Triathlon. Sports 2021, 9, 120. [Google Scholar] [CrossRef] [PubMed]
- Arfuso, F.; Rizzo, M.; Giannetto, C.; Giudice, E.; Cirincione, R.; Cassata, G.; Cicero, L.; Piccione, G. Oxidant and Antioxidant Parameters’ Assessment Together with Homocysteine and Muscle Enzymes in Racehorses: Evaluation of Positive Effects of Exercise. Antioxidants 2022, 11, 1176. [Google Scholar] [PubMed]
- Kowalik, S.; Tomaszewska, E. Does routine exercise induce stress and in consequence serum amyloid A (SAA) response in purebred Arabian racehorses? Pferdeheilkunde 2018, 34, 121–125. [Google Scholar] [CrossRef]
- Brooks, G.A. Cell–cell and intracellular lactate shuttles. J. Physiol. 2009, 587, 5591–5600. [Google Scholar] [CrossRef] [PubMed]
- Volfinger, L.; Lassourd, V.; Michaux, J.; Braun, J.; Toutain, P. Kinetic evaluation of muscle damage during exercise by calculation of amount of creatine kinase released. Am. J. Physiol. 1994, 266, R434–R441. [Google Scholar] [CrossRef]
- Glitz, F. Muskelenzymmuster und Elimination von iv Applizierten, Homologen Muskelenzymen Beim Pferd; Tierärztliche Hochschule Hannover: Hannover, Germany, 1997. [Google Scholar]
- Allaam, M.; ELseady, Y.; Nayel, M.; Elsify, A.; Salama, A.; Hassan, H.; Hassan, M.; Kamar, A. Physiological and hemato-chemical evaluation of thoroughbred race horse after exercise. IJAVMS 2014, 8, 81–93. [Google Scholar]
- Ebrahim, Z.K.; Metwally, A.M.; Elshahawy, I.I. Some clinical, hematological and biochemical alterations in endurance horses after 40 km endurance race. Alex. J. Vet. Sci. 2019, 61, 133–139. [Google Scholar] [CrossRef]
- Hassan, H.Y.; Aly, M.A.; Elseady, Y.M.; Nayel, M.A.; Elsify, A.M.; Salama, A.A.; Hassan, M.S.; Elbarody, E.F.; Kamar, A.B. The effect of race in the clinical, hematological and biochemical biomarkers in Thoroughbred horses. Alex. J. Vet. Sci. 2015, 46, 161–169. [Google Scholar] [CrossRef]
- Miglio, A.; Cappelli, K.; Capomaccio, S.; Mecocci, S.; Silvestrelli, M.; Antognoni, M.T. Metabolic and biomolecular changes induced by incremental long-term training in young thoroughbred racehorses during first workout season. Animals 2020, 10, 317. [Google Scholar] [CrossRef] [PubMed]
- McKeever, K.; Hinchcliff, K.; Schmall, L.; Muir, W., 3rd. Renal tubular function in horses during submaximal exercise. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1991, 261, R553–R560. [Google Scholar] [CrossRef]
- Hinchcliff, K.; McKeever, K.; Schmall, L.; Kohn, C.; Muir, W., 3rd. Renal and systemic hemodynamic responses to sustained submaximal exertion in horses. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1990, 258, R1177–R1183. [Google Scholar] [CrossRef]
- Rose, R.J.; Arnold, K.S.; Church, S.; Paris, R. Plasma and sweat electrolyte concentrations in the horse during long distance exercise. Equine Vet. J. 1980, 12, 19–22. [Google Scholar] [CrossRef]
- Weiss, D.; Weishoupt, M.A.; Forrer, R.; Fakler, A.; Spichiger, U.E.; Burger, D.; Wanner, M.; Riond, J.L. Effects of sweat loss induced by treadmill exercise on magnesium and calcium homeostasis in Franches-Montagnes horses. Pferdeheilkunde 2002, 18, 5–10. [Google Scholar] [CrossRef]
- Hoyt, J.K.; Potter, G.D.; Greene, L.W.; Anderson, J.G. Mineral balance in resting and exercised miniature horses. J. Equine Vet. Sci. 1995, 15, 310–314. [Google Scholar] [CrossRef]
- Degens, H.; Jones, D.A. Are Force Enhancement after Stretch and Muscle Fatigue Due to Effects of Elevated Inorganic Phosphate and Low Calcium on Cross Bridge Kinetics? Medicina 2020, 56, 249. [Google Scholar] [CrossRef]
- Zobba, R.; Ardu, M.; Niccolini, S.; Cubeddu, F.; Dimauro, C.; Bonelli, P.; Dedola, C.; Visco, S.; Pinna Parpaglia, M.L. Physical, Hematological, and Biochemical Responses to Acute Intense Exercise in Polo Horses. J. Equine Vet. Sci. 2011, 31, 542–548. [Google Scholar] [CrossRef]
- Lhotta, K. Störungen des Phosphathaushaltes. J. Klin. Endokrinol. Stoffwechs. 2011, 4, 20–23. [Google Scholar]
- Hureau, T.J.; Broxterman, R.M.; Weavil, J.C.; Lewis, M.T.; Layec, G.; Amann, M. On the role of skeletal muscle acidosis and inorganic phosphates as determinants of central and peripheral fatigue: A (31) P-MRS study. J. Physiol. 2022, 600, 3069–3081. [Google Scholar] [CrossRef] [PubMed]
- Von Engelhardt, W.; Breves, G. Physiologie der Haustiere. Swiss Arch. Vet. Med. 2005, 147, 507. [Google Scholar] [CrossRef]
- Hickman, P.E.; Potter, J.M.; Aroney, C.; Koerbin, G.; Southcott, E.; Wu, A.H.; Roberts, M.S. Cardiac troponin may be released by ischemia alone, without necrosis. Clin. Chim. Acta 2010, 411, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Sabatine, M.S.; Morrow, D.A.; De Lemos, J.A.; Jarolim, P.; Braunwald, E. Detection of acute changes in circulating troponin in the setting of transient stress test-induced myocardial ischaemia using an ultrasensitive assay: Results from TIMI 35. Eur. Heart J. 2008, 30, 162–169. [Google Scholar] [CrossRef]
- Wijnberg, I.D.; Franklin, S.H. The heart remains the core: Cardiac causes of poor performance in horses compared to human athletes. Comp. Exerc. Physiol. 2017, 13, 149–174. [Google Scholar] [CrossRef]
- Trachsel, D.S.; Schwarzwald, C.C.; Bitschnau, C.; Grenacher, B.; Weishaupt, M.A. Atrial natriuretic peptide and cardiac troponin I concentrations in healthy Warmblood horses and in Warmblood horses with mitral regurgitation at rest and after exercise. J. Vet. Cardiol. 2013, 15, 105–121. [Google Scholar] [CrossRef]
- Eijsvogels, T.M.; Hoogerwerf, M.D.; Oudegeest-Sander, M.H.; Hopman, M.T.; Thijssen, D.H. The impact of exercise intensity on cardiac troponin I release. Int. J. Cardiol. 2014, 171, e3–e4. [Google Scholar] [CrossRef]
- Legaz-Arrese, A.; López-Laval, I.; George, K.; Puente-Lanzarote, J.J.; Mayolas-Pi, C.; Serrano-Ostáriz, E.; Revilla-Martí, P.; Moliner-Urdiales, D.; Reverter-Masià, J. Impact of an endurance training program on exercise-induced cardiac biomarker release. Am. J. Physiol. Heart Circ. Physiol. 2015, 308, H913–H920. [Google Scholar] [CrossRef]
- Gresslien, T.; Agewall, S. Troponin and exercise. Int. J. Cardiol. 2016, 221, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Degens, H.; Veerkamp, J.H. Changes in oxidative capacity and fatigue resistance in skeletal muscle. Int. J. Biochem. 1994, 26, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Arfuso, F.; Giannetto, C.; Giudice, E.; Fazio, F.; Panzera, M.; Piccione, G. Peripheral Modulators of the Central Fatigue Development and Their Relationship with Athletic Performance in Jumper Horses. Animals 2021, 11, 743. [Google Scholar] [CrossRef] [PubMed]
- Ament, W.; Verkerke, G.J. Exercise and fatigue. Sports Med. 2009, 39, 389–422. [Google Scholar] [CrossRef] [PubMed]
- Edwards, R.H. Human muscle function and fatigue. In Proceedings of the Ciba Foundation Symposium 82-Human Muscle Fatigue: Physiological Mechanisms, London, UK, 9–11 September 1980; Volume 82, pp. 1–18. [Google Scholar] [CrossRef]
- Scott, J.M.; Warburton, D.E. Mechanisms underpinning exercise-induced changes in left ventricular function. Med. Sci. Sports Exerc. 2008, 40, 1400–1407. [Google Scholar] [CrossRef] [PubMed]
- Brancaccio, P.; Limongelli, F.M.; Maffulli, N. Monitoring of serum enzymes in sport. Br. J. Sports Med. 2006, 40, 96–97. [Google Scholar] [CrossRef] [PubMed]
- Friedrichs, K.R.; Harr, K.E.; Freeman, K.P.; Szladovits, B.; Walton, R.M.; Barnhart, K.F.; Blanco-Chavez, J. ASVCP reference interval guidelines: Determination of de novo reference intervals in veterinary species and other related topics. Vet. Clin. Pathol. 2012, 41, 441–453. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Horse | Age | Sex | Breed |
|---|---|---|---|
| 1 | 7 | Mare | Holsteiner |
| 2 | 12 | Mare | Stud Book du Cheval Selle Français |
| 3 | 12 | Gelding | Hanoverian |
| 4 | 15 | Mare | Hanoverian |
| 5 | 12 | Gelding | Irish Sport Horse |
| 6 | 14 | Gelding | Hanoverian |
| 7 | 9 | Gelding | Hanoverian |
| 8 | 10 | Mare | Polish Horse Breeders Association |
| 9 | 15 | Mare | Rheinlander |
| 10 | 9 | Mare | German Sport Horse |
| 11 | 7 | Mare | Oldenburger |
| 12 | 8 | Mare | Oldenburger |
| 13 | 8 | Gelding | Westphalian |
| 14 | 14 | Gelding | Holsteiner |
| 15 | 11 | Gelding | Hanoverian |
| 16 | 12 | Mare | Hanoverian |
| 17 | 10 | Gelding | Irish Sport Horse |
| 18 | 11 | Gelding | Holsteiner |
| 19 | 15 | Gelding | Hanoverian |
| 20 | 7 | Mare | Hanoverian |
| Competition | Level | Week | Venue | Horses N = | Distance [m] | Speed Required [m/min] | Temperature [°C] | Ground | Altitude Profile | Weather |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | CCI3*-S | 12 | A | 11 | 3401 | 550 | 13 | normal | flat | cloudy |
| 2 | CCI3*-S | 15 | B | 7 | 3007 | 520 | 16 | normal | intermediate | sunny |
| 3 | CCI2*-S | 16 | A | 2 | 3000 | 520 | 16 | normal | flat | cloudy |
| 4 | CIC2*-L | 18 | C | 1 | 3787 | 520 | 16 | deep | hilly | sunny |
| 5 | CCI2*-S | 18 | C | 2 | 3085 | 520 | 18 | normal | hilly | cloudy |
| 6 | CCI4*-S | 18 | C | 7 | 3705 | 570 | 20 | deep | hilly | sunny |
| 7 | CCI4*-S | 24 | A | 3 | 3772 | 570 | 29 | normal | flat | sunny |
| 8 | CIC3*-L | 25 | A | 3 | 4455 | 550 | 26 | normal | flat | sunny |
| 9 | CCI3*-S | 25 | A | 3 | 3364 | 550 | 25 | normal | flat | sunny |
| 10 | CCI2*-S | 30 | D | 2 | 2661 | 520 | 20 | normal | hilly | cloudy |
| 11 | CCI3*-S | 30 | D | 6 | 3538 | 550 | 20 | normal | hilly | cloudy |
| 12 | CCI2*-S | 35 | E | 1 | 3087 | 520 | 28 | normal | flat | sunny |
| 13 | CCI3*-S | 35 | E | 2 | 3470 | 550 | 28 | normal | flat | sunny |
| 14 | CCI4*-S | 35 | E | 5 | 3580 | 570 | 28 | normal | flat | sunny |
| Category | Biomarker |
|---|---|
| Fluid Balance | RBC, HCT, TP, Albumin, UREA, SDMA, Chloride, Sodium |
| Muscle Enzymes | CK, LDH, AST |
| Muscle Metabolites | Lactate, Creatinine, Inorg. Phosphate, Calcium, Magnesium, Potassium |
| Cardiac Markers | Cardiac Troponin I (cTnI) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giers, J.; Bartel, A.; Kirsch, K.; Müller, S.F.; Horstmann, S.; Gehlen, H. Blood-Based Markers for Skeletal and Cardiac Muscle Function in Eventing Horses before and after Cross-Country Rides and How They Are Influenced by Plasma Volume Shift. Animals 2023, 13, 3110. https://doi.org/10.3390/ani13193110
Giers J, Bartel A, Kirsch K, Müller SF, Horstmann S, Gehlen H. Blood-Based Markers for Skeletal and Cardiac Muscle Function in Eventing Horses before and after Cross-Country Rides and How They Are Influenced by Plasma Volume Shift. Animals. 2023; 13(19):3110. https://doi.org/10.3390/ani13193110
Chicago/Turabian StyleGiers, Johanna, Alexander Bartel, Katharina Kirsch, Simon Franz Müller, Stephanie Horstmann, and Heidrun Gehlen. 2023. "Blood-Based Markers for Skeletal and Cardiac Muscle Function in Eventing Horses before and after Cross-Country Rides and How They Are Influenced by Plasma Volume Shift" Animals 13, no. 19: 3110. https://doi.org/10.3390/ani13193110
APA StyleGiers, J., Bartel, A., Kirsch, K., Müller, S. F., Horstmann, S., & Gehlen, H. (2023). Blood-Based Markers for Skeletal and Cardiac Muscle Function in Eventing Horses before and after Cross-Country Rides and How They Are Influenced by Plasma Volume Shift. Animals, 13(19), 3110. https://doi.org/10.3390/ani13193110