

Trends of Eurasian Perch (Perca fluviatilis) mtDNA ATP6 Region Genetic Diversity within the Hydro-Systems of the Eastern Part of the Baltic Sea in the Anthropocene

 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
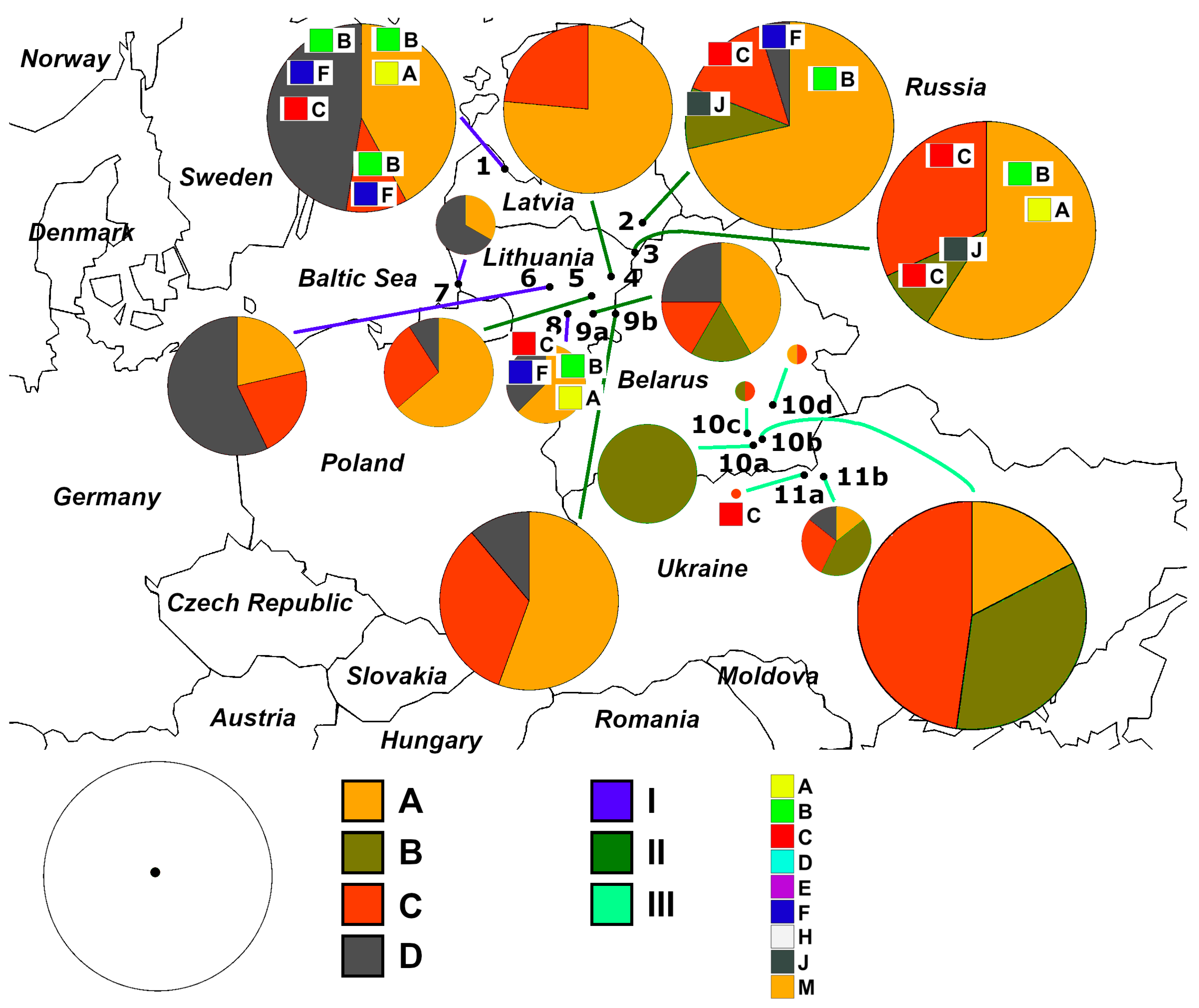
2.1. Sampling and Data Treatment
2.2. DNA Extraction, Amplification, and Sequencing
2.3. Analysis of Intraspecific Genetic Data
2.4. Comparisons of Genetic Diversity Parameters between mtDNA ATP6 and D-Loop Regions
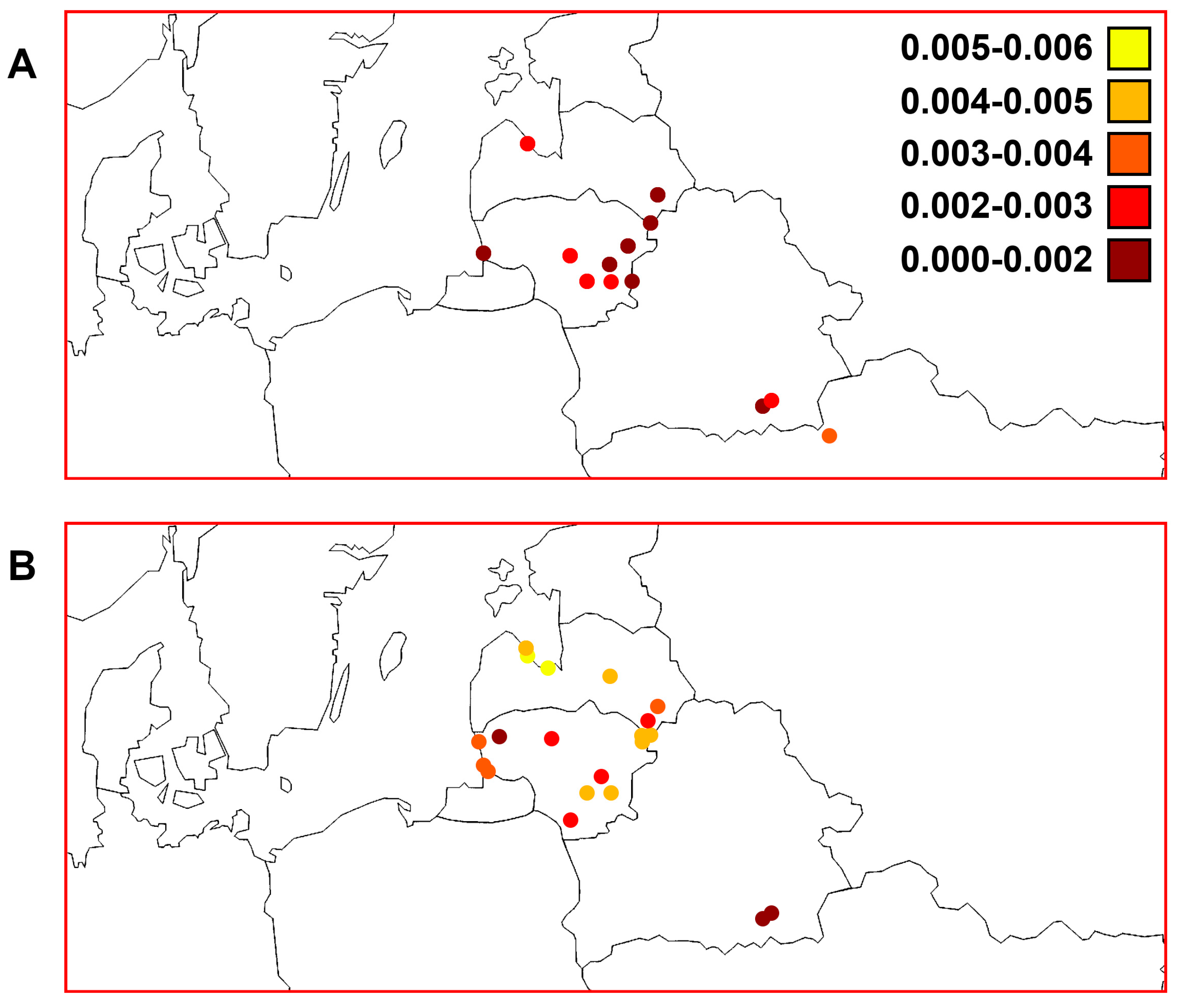
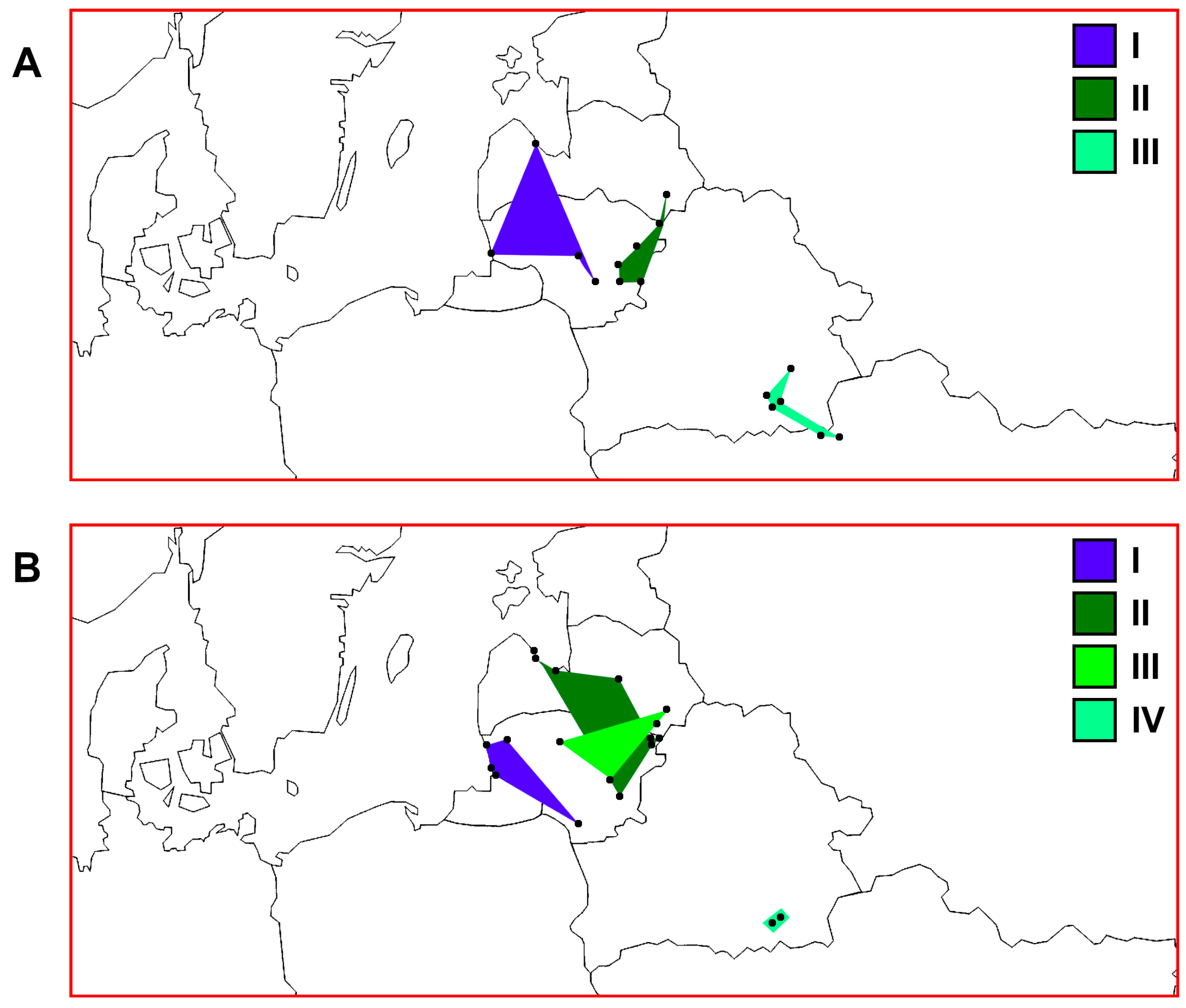
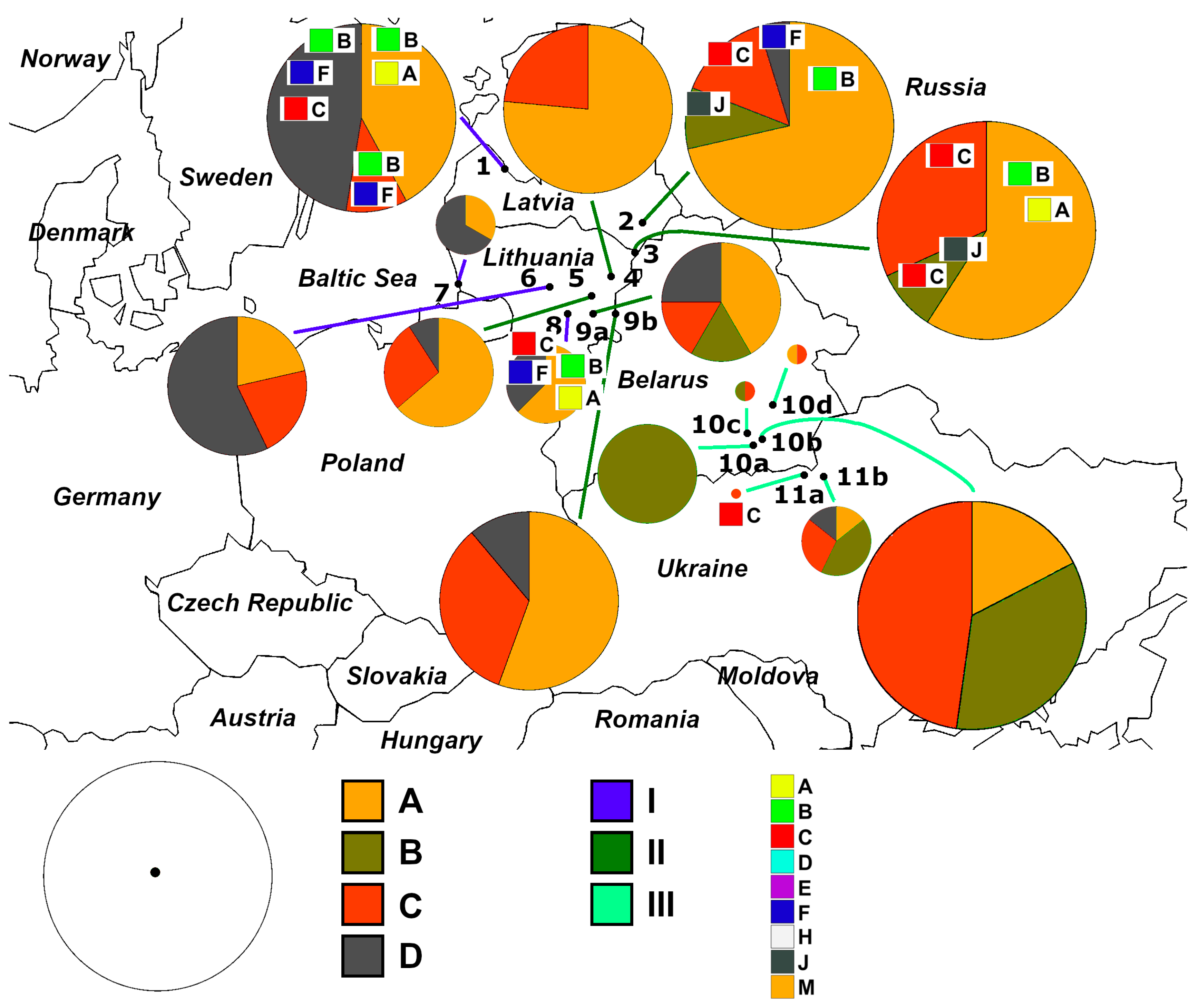
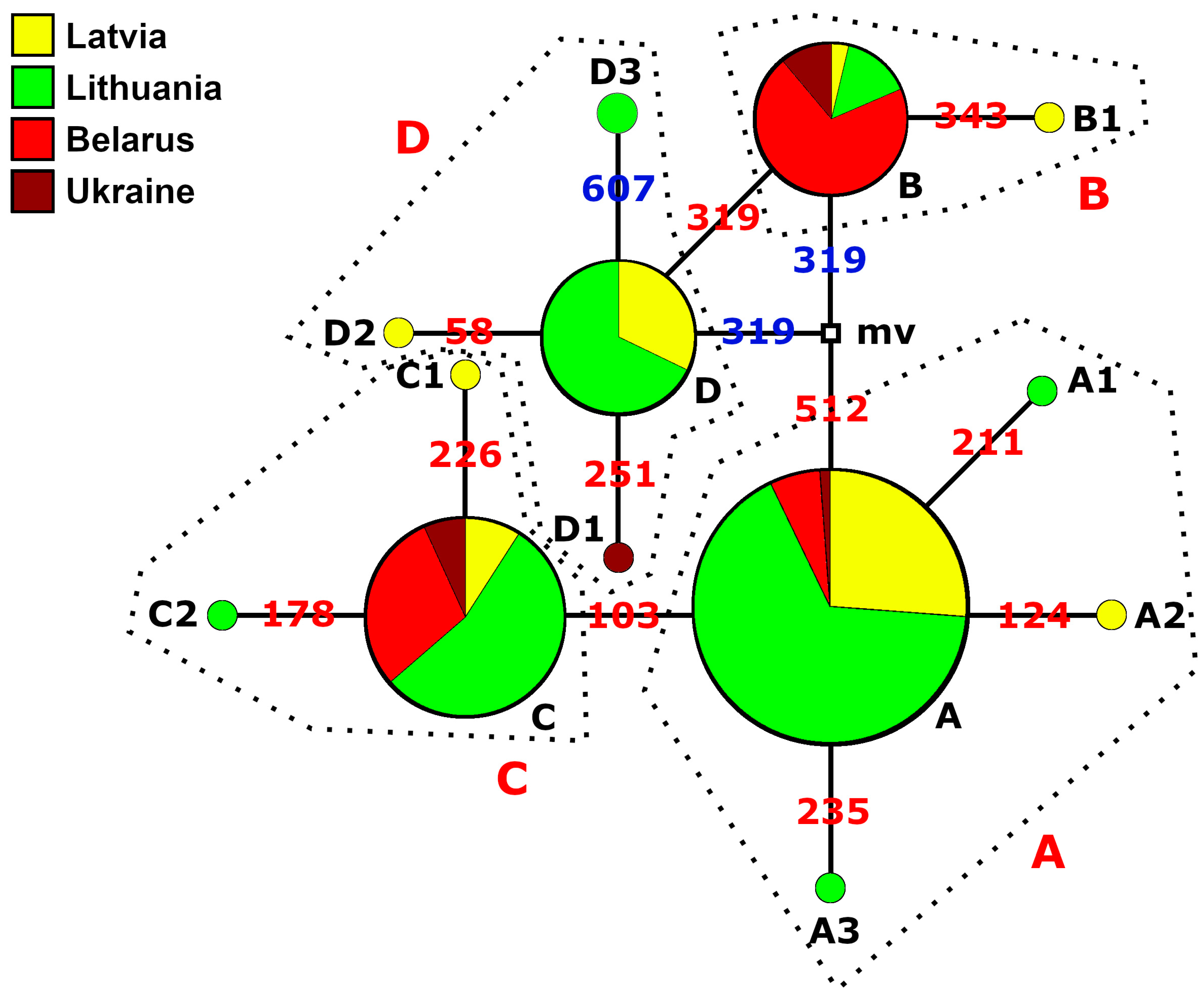
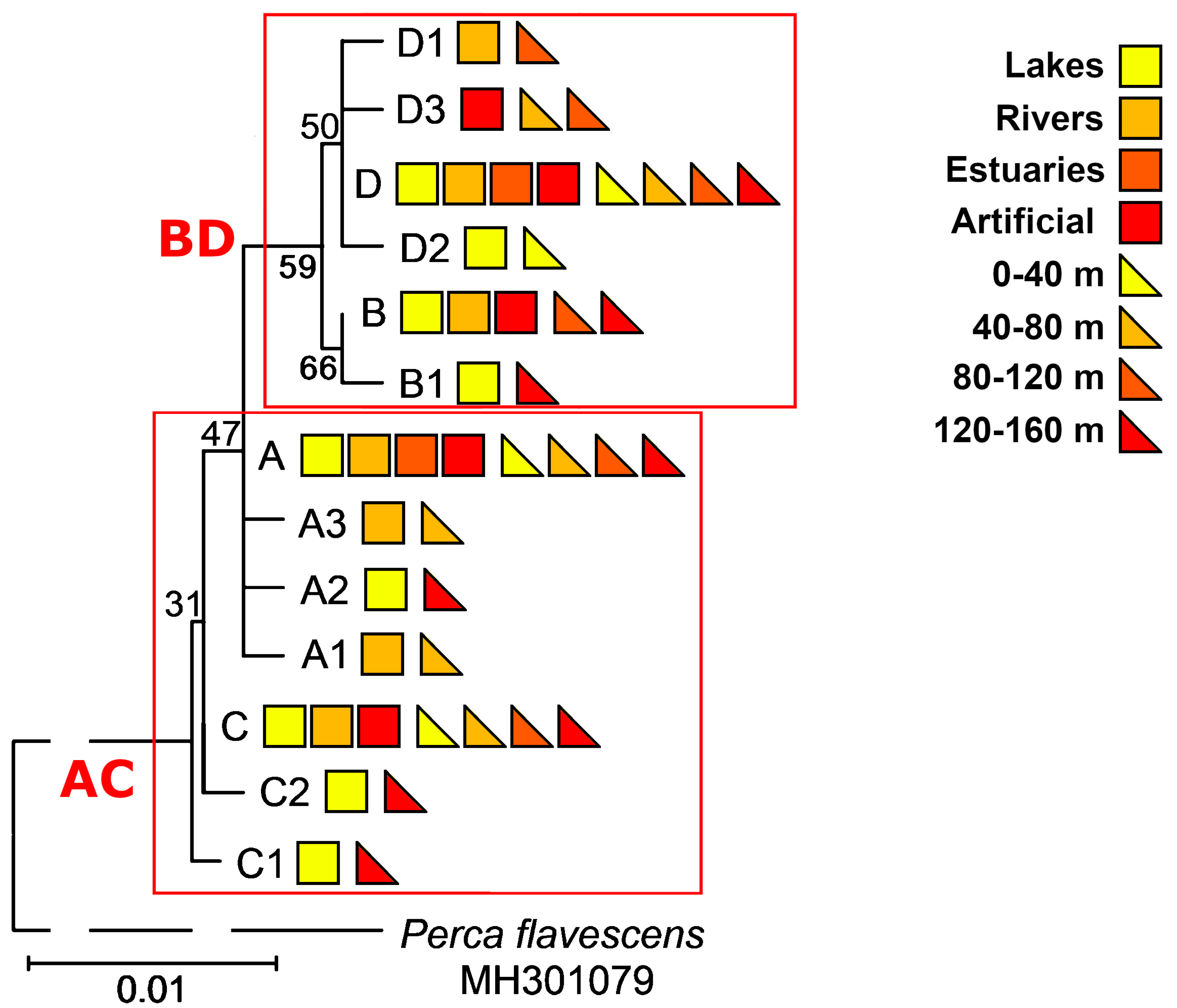
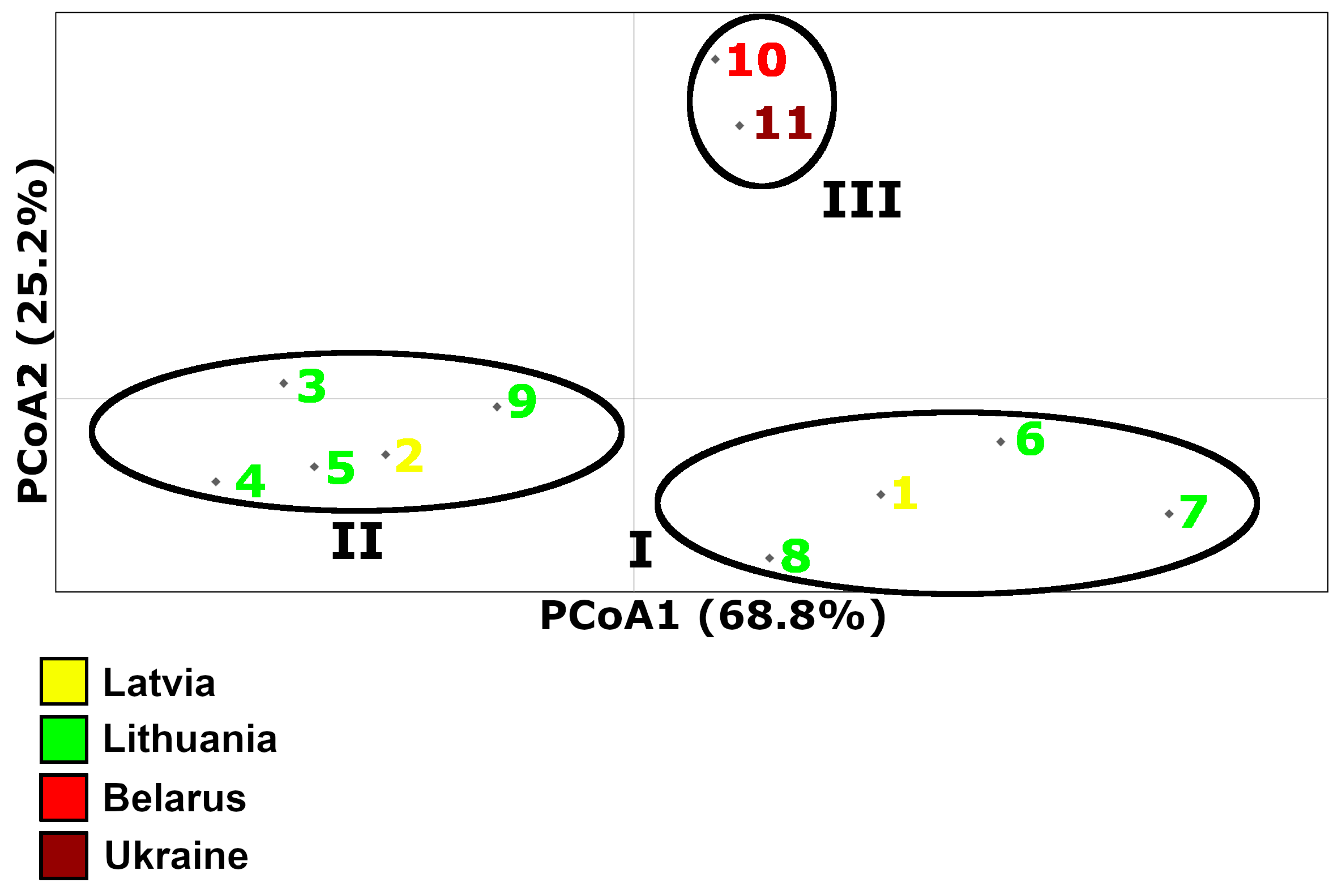
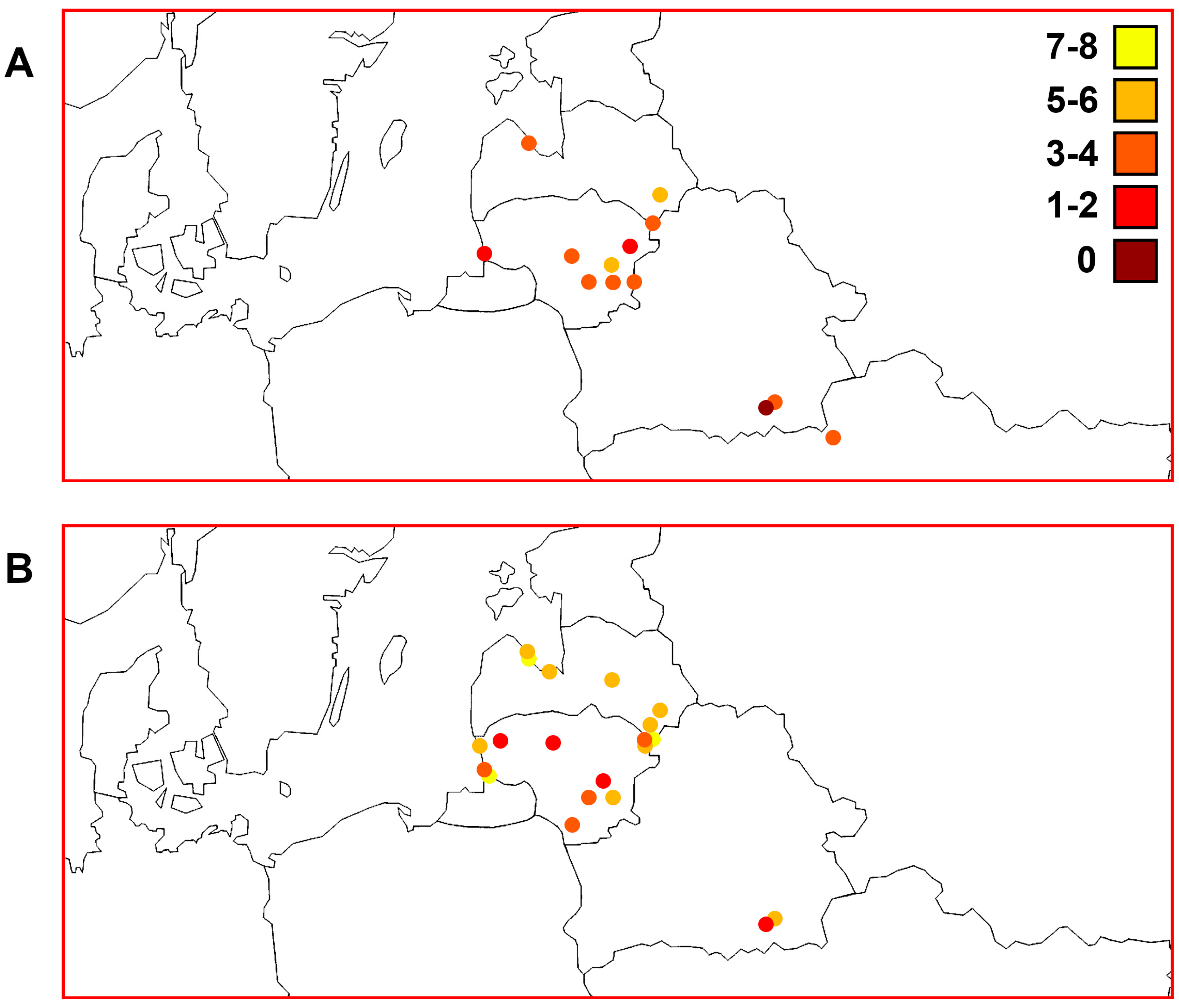
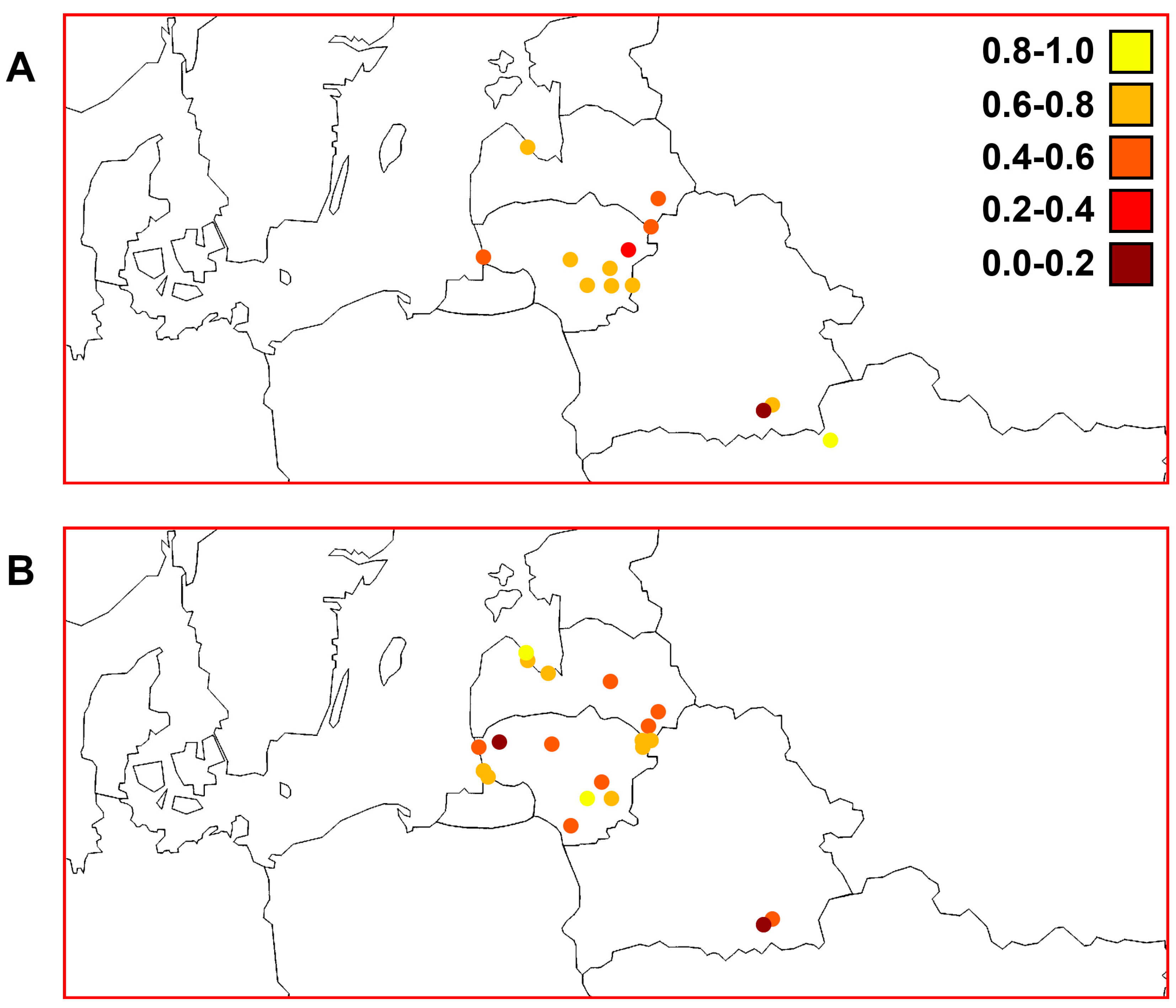
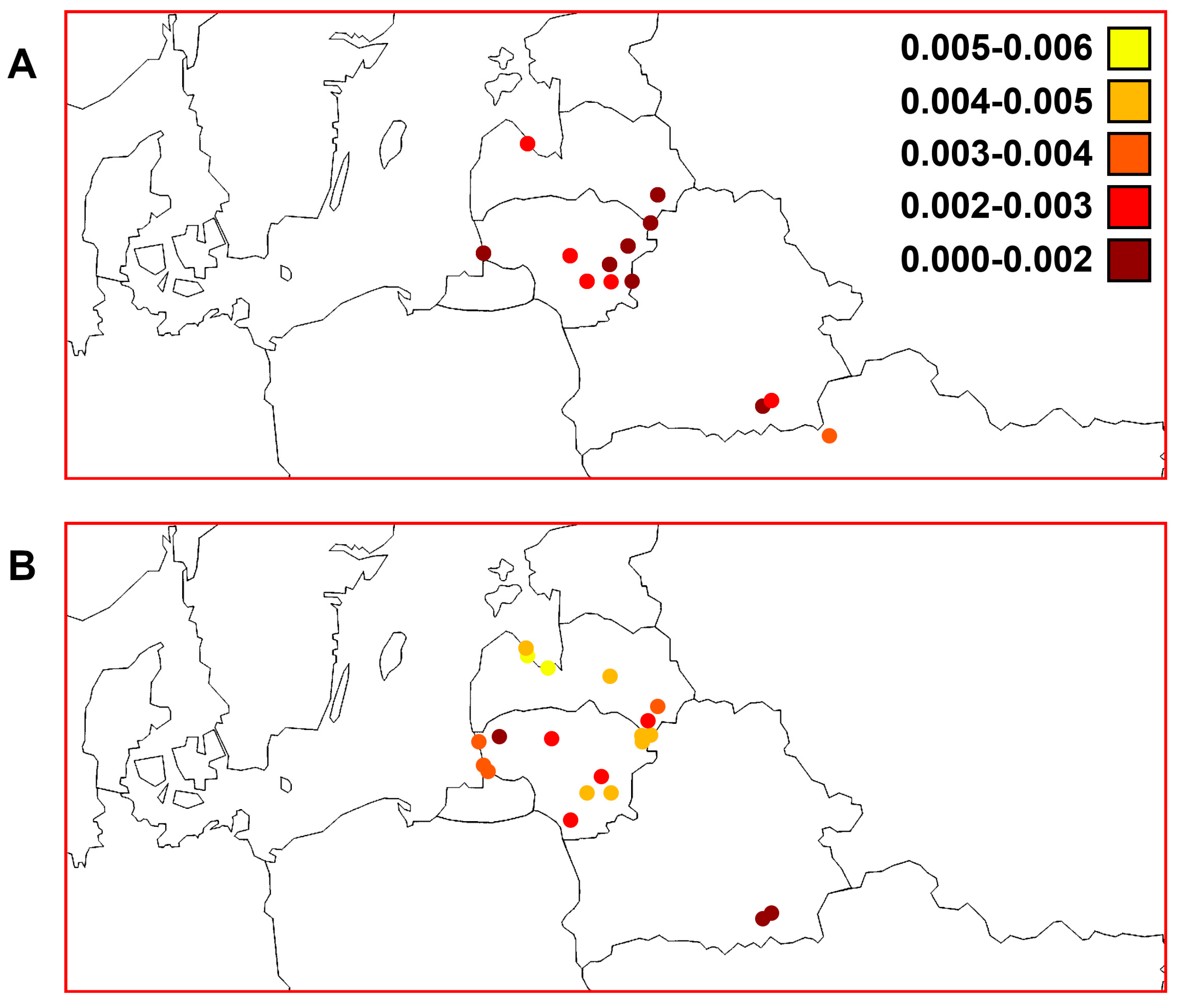
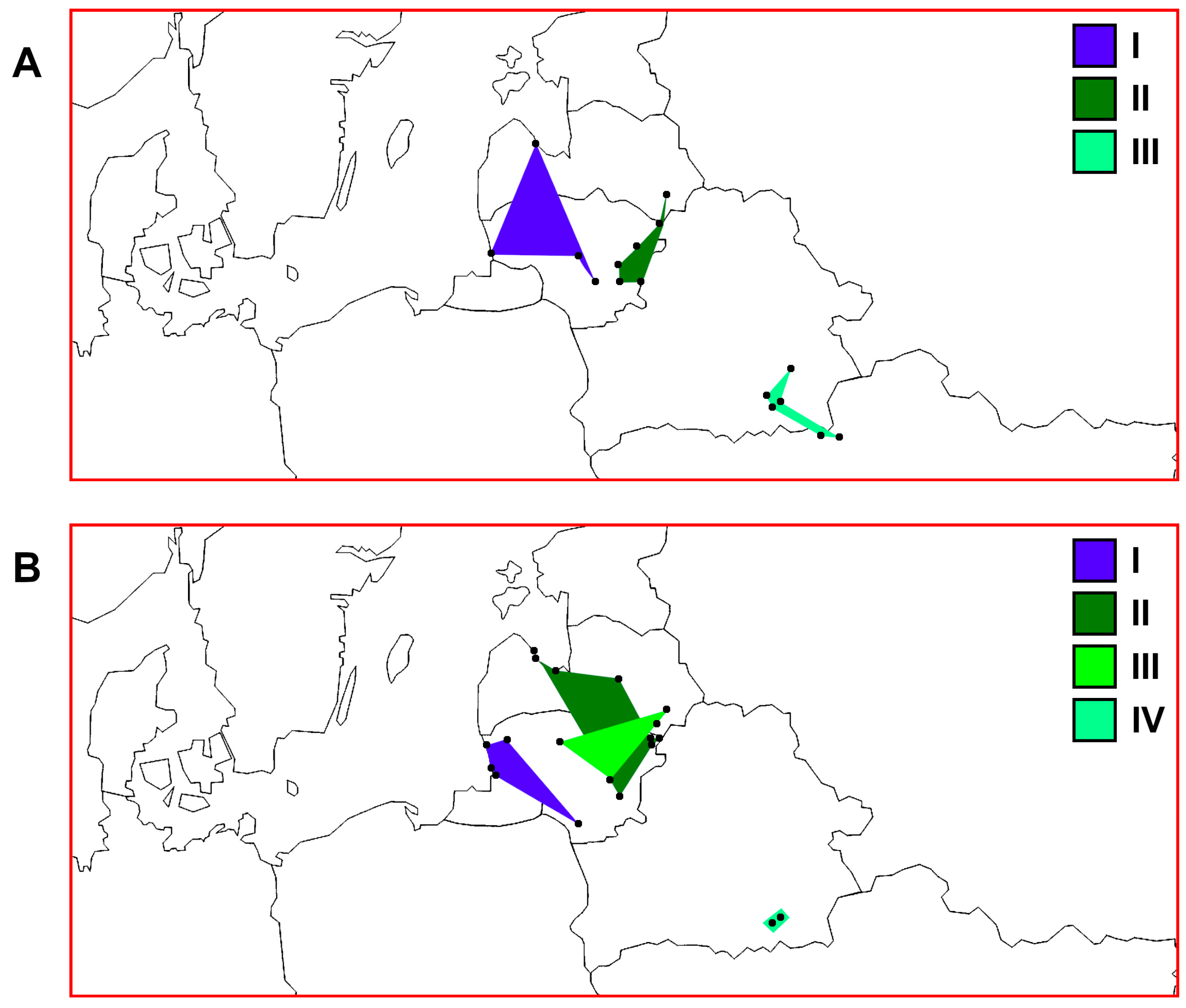
3. Results
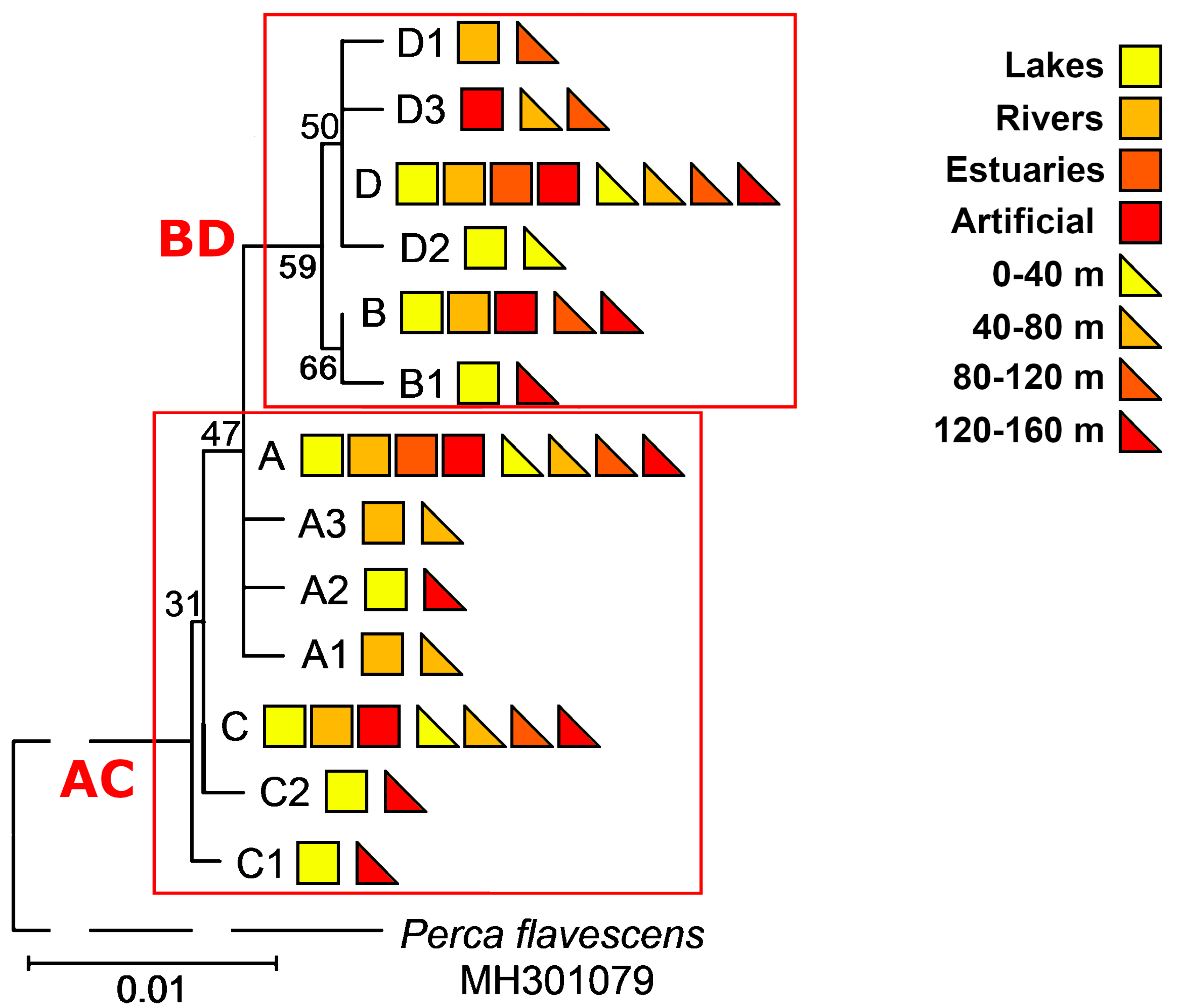
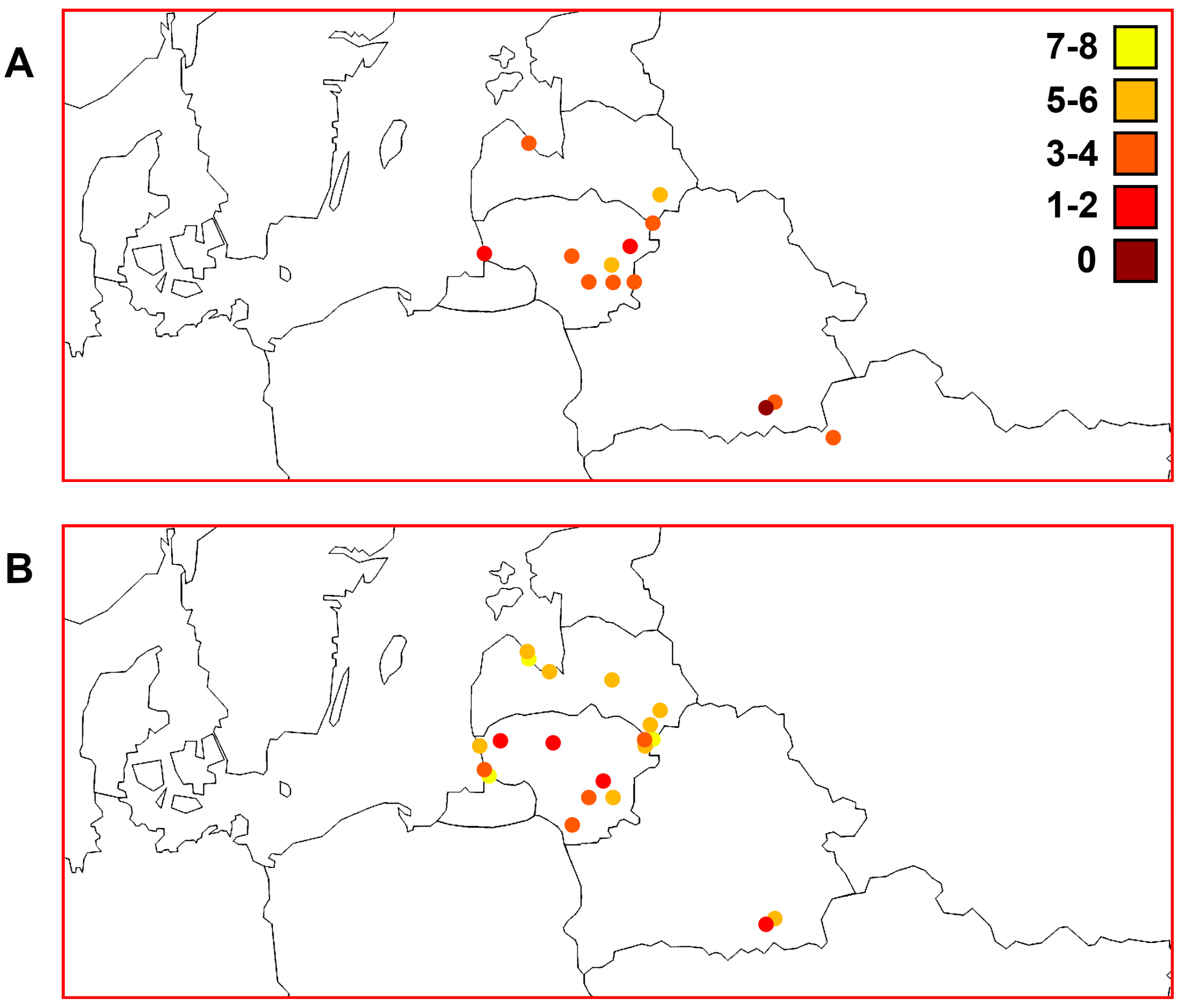
3.1. Identification of Haplotypes and Haplogroups and Their Distribution Patterns
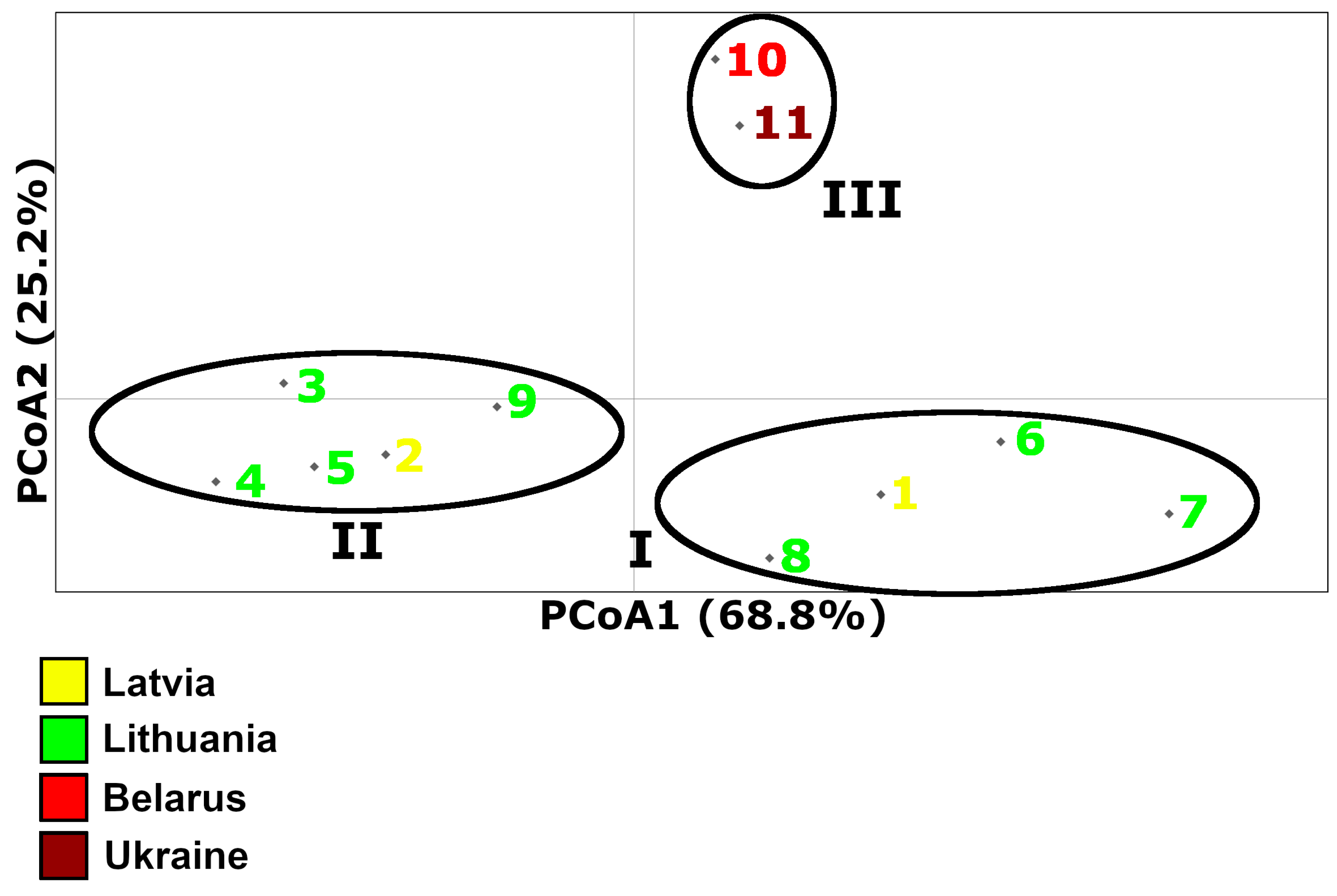
3.2. PCoA Results
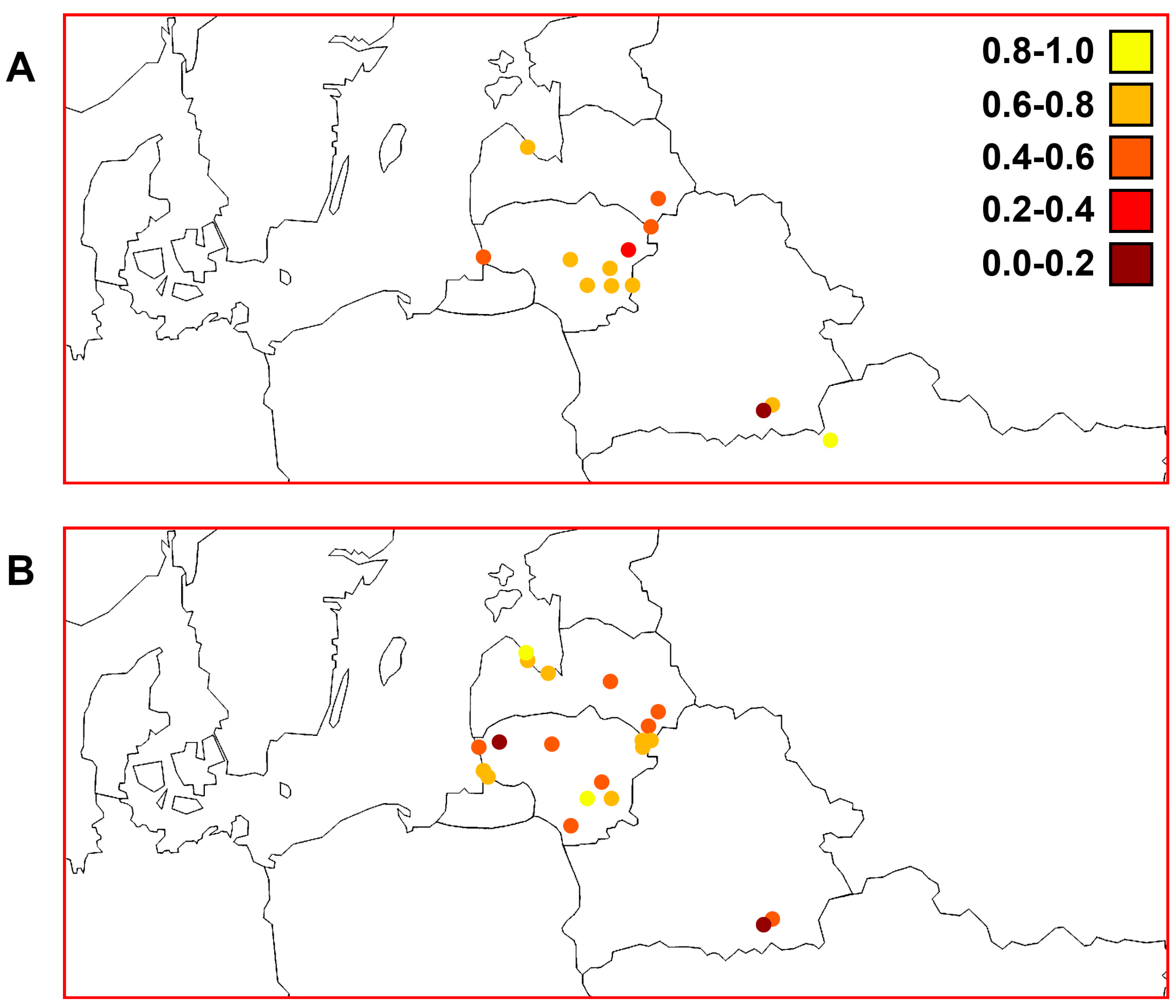
3.3. Quantitative Parameters of Genetic Variability and Genetic Differentiation among Perch Samples
3.4. Protein Sequence Variability of the mtDNA ATP6 Region
3.5. Natural Selection
3.6. Haplotypes and Haplogroups of mtDNA ATP6 and D-Loop Regions of the Same Specimens
3.7. Comparisons of Trends of Genetic Diversity of Perch within the Studied Macrogeographic Area Using Different mtDNA Markers
4. Discussion
4.1. Application of the ATP6 Region to Study Perch Population Genetics and Phylogeography
4.2. Anthropogenic Context
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Capps, K.A.; Chapman, S.; Clay, K.; Fresnedo-Ramírez, J.; Potts, D.L. Reshaping the tree of life: Ecological implications of evolution in the anthropocene. Front. Ecol. Environ. 2021, 20, 111–116. [Google Scholar] [CrossRef]
- Exposito-Alonso, M.; Booker, T.R.; Czech, L.; Gillespie, L.; Hateley, S.; Kyriazis, C.C.; Lang, P.L.M.; Leventhal, L.; Nogues-Bravo, D.; Pagowski, V.; et al. Genetic diversity loss in the anthropocene. Science 2022, 377, 1431–1435. [Google Scholar] [CrossRef]
- Rashad, S.M.; Hammad, F.H. Nuclear power and the environment: Comparative assessment of environmental and health impacts of electricity-generating systems. Appl. Energy 2000, 65, 211–229. [Google Scholar] [CrossRef]
- Montévil, M. Entropies and the anthropocene crisis. AI Soc. 2021, 1–32. [Google Scholar] [CrossRef]
- Leonard, M.D.; Michaelides, E.E.; Michaelides, D.M. Energy storage needs for the substitution of fossil fuel power plants with renewables. Renew. Energy 2020, 145, 951–962. [Google Scholar] [CrossRef]
- Deshmukh, M.K.G.; Sameeroddin, M.; Abdul, D.; Sattar, M.A. Renewable energy in the 21st century: A review. Mater. Today Proc. 2023, 80, 1756–1759. [Google Scholar] [CrossRef]
- Kashparov, V. Lessons learned from the Chernobyl accident. In Nuclear Emergencies Current Topics in Environmental Health and Preventive Medicine; Steinhauser, G., Koizumi, A., Shozugawa, K., Eds.; Springer: Singapore, 2019; pp. 23–36. [Google Scholar] [CrossRef]
- Buesseler, K.; Dai, M.; Aoyama, M.; Benitez-Nelson, C.; Charmasson, S.; Higley, K.; Maderich, V.; Masqué, P.; Morris, P.J.; Oughton, D.; et al. Fukushima Daiichi-derived radionuclides in the ocean: Transport, fate, and impacts. Ann. Rev. Mar. Sci. 2017, 9, 173–203. [Google Scholar] [CrossRef]
- Lerebours, A.; Robson, S.; Sharpe, C.; Nagorskaya, L.; Gudkov, D.; Haynes-Lovatt, C.; Smith, J.T. Transcriptional changes in the ovaries of perch from Chernobyl. ASAP 2020, 54, 10078–10087. [Google Scholar] [CrossRef] [PubMed]
- Steinhauser, G.; Brandl, A.; Johnson, T.E. Comparison of the Chernobyl and Fukushima nuclear accidents: A review of the environmental impacts. Environ. Sci. Technol. 2014, 470–471, 800–817. [Google Scholar] [CrossRef]
- Aliyu, A.S.; Evangeliou, N.; Mousseau, T.A.; Wu, J.; Ramli, A.T. An overview of current knowledge concerning the health and environmental consequences of the Fukushima Daiichi Nuclear Power Plant (FDNPP) accident. Environ. Int. 2015, 85, 213–228. [Google Scholar] [CrossRef]
- Beresford, N.A.; Fesenko, S.; Konoplev, A.; Skuterud, L.; Smith, J.T.; Voigt, G. Thirty years after the Chernobyl accident: What lessons have we learnt? J. Environ. Radioact. 2016, 157, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Geras’kin, S.A.; Fesenko, S.V.; Volkova, P.Y.; Isamov, N.N. What have we learned about the biological effects of radiation from the 35 Years of analysis of the consequences of the Chernobyl NPP accident? Biol. Bull. Russ. Acad. Sci. 2021, 48, 2105–2126. [Google Scholar] [CrossRef]
- Spatola, G.J.; Buckley, R.M.; Dillon, M.; Dutrow, E.M.; Betz, J.A.; Pilot, M.; Parker, H.G.; Bogdanowicz, W.; Thomas, R.; Chyzhevskyi, I.; et al. The dogs of Chernobyl: Demographic insights into populations inhabiting the nuclear exclusion zone. Sci. Adv. 2023, 9, eade2537. [Google Scholar] [CrossRef]
- Geras’kin, S.A.; Fesenko, S.V.; Alexakhin, R.M. Effects of non-human species irradiation after the Chernobyl NPP accident. Environ. Int. 2008, 34, 880–897. [Google Scholar] [CrossRef]
- Møller, A.P.; Mousseau, T.A. Biological consequences of Chernobyl: 20 years on. Trends Ecol. Evol. 2006, 21, 200–207. [Google Scholar] [CrossRef]
- Raptis, C.E.; Boucher, J.M.; Pfister, S. Assessing the environmental impacts of freshwater thermal pollution from global power generation in LCA. Sci. Total Environ. 2017, 580, 1014–1026. [Google Scholar] [CrossRef]
- Aydın, C.İ. Nuclear energy debate in Turkey: Stakeholders, policy alternatives, and governance issues. Energy Policy 2020, 136, 111041. [Google Scholar] [CrossRef]
- Virbickas, J.; Virbickas, T. Results and prospects of integrated long-term ecological research into Lake Drūkšiai. Acta Zool. Litu. 2005, 15, 195–198. [Google Scholar] [CrossRef]
- Makhrov, A.A.; Bolotov, I.N. Dispersal routes and species identification of freshwater animals in Northern Europe: A review of molecular evidence. Genetika 2006, 42, 1319–1334. [Google Scholar] [CrossRef]
- Brucet, S.; Pedron, S.; Mehner, T.; Lauridsen, T.L.; Argillier, C.; Winfield, I.J.; Volta, P.; Emmrich, M.; Hesthagen, T.; Holmgren, K.; et al. Fish diversity in European lakes: Geographical factors dominate over anthropogenic pressures. Freshw. Biol. 2013, 58, 1779–1793. [Google Scholar] [CrossRef]
- Manel, S.; Guerin, P.E.; Mouillot, D.; Blanchet, S.; Velez, L.; Albouy, C.; Pellissier, L. Global determinants of freshwater and marine fish genetic diversity. Nat. Commun. 2020, 11, 692. [Google Scholar] [CrossRef] [PubMed]
- Gordon, T.A.C.; Harding, H.R.; Clever, F.K.; Davidson, I.K.; Davison, W.; Montgomery, D.W.; Weatherhead, R.C.; Windsor, F.M.; Armstrong, J.D.; Bardonnet, A.; et al. Fishes in a changing world: Learning from the past to promote sustainability of fish populations. J. Fish Biol. 2018, 92, 804–827. [Google Scholar] [CrossRef] [PubMed]
- Skúlason, S.; Parsons, K.J.; Svanbäck, R.; Räsänen, K.; Ferguson, M.M.; Adams, C.E.; Amundsen, P.-A.; Bartels, P.; Bean, C.W.; Boughman, J.W.; et al. A way forward with eco evo devo: An extended theory of resource polymorphism with postglacial fishes as model systems. Biol. Rev. 2019, 94, 1786–1808. [Google Scholar] [CrossRef] [PubMed]
- Škute, N.; Oreha, J.; Krivmane, B.; Evarte, A. Genetic structure of perch Perca Fluviatilis (L.) populations in Latvian rivers that are fragmented (Daugava) and non-fragmented (Lielupe) By hydroelectric dams. Proc. Latv. Acad. Sci. B Nat. Exact Appl. Sci. 2021, 75, 211–219. [Google Scholar] [CrossRef]
- Brodeur, R.D.; Smith, B.E.; McBride, R.S.; Heintz, R.; Farley Jr, E. New perspectives on the feeding ecology and trophic dynamics of fishes. Environ. Biol. Fish 2017, 100, 293–297. [Google Scholar] [CrossRef]
- Kesminas, V.; Olechnovičienė, J. Fish community changes in the cooler of the Ignalina nuclear power plant. Ekologija 2008, 54, 124–131. [Google Scholar] [CrossRef]
- Ladich, F. Ecology of sound communication in fishes. Fish Fish. 2019, 20, 552–563. [Google Scholar] [CrossRef]
- Simpson, S.D.; Barber, I.; Winfield, I.J. Understanding fish populations. J. Fish Biol. 2018, 92, 563–568. [Google Scholar] [CrossRef]
- Tamario, C.; Sunde, J.; Petersson, E.; Tibblin, P.; Forsman, A. Ecological and evolutionary consequences of environmental change and management actions for migrating fish. Front. Ecol. Evol. 2019, 7, 271. [Google Scholar] [CrossRef]
- Laikre, L.; Palm, S.; Ryman, N. Genetic population structure of fishes: Implications for coastal zone management. Ambio 2005, 34, 111–119. [Google Scholar] [CrossRef]
- Schmidt, T.; Zagars, M.; Roze, A.; Schulz, R. Genetic diversity of a Daugava basin brown trout (Salmo trutta) brood stock. Knowl. Manag. Aquat. Ecosyst. 2017, 418, 55. [Google Scholar] [CrossRef]
- Wennerström, L.; Jansson, E.E.; Laikre, L. Baltic Sea genetic biodiversity: Current knowledge relating to conservation management. Aquat. Conserv. Mar. Freshw. 2017, 27, 1069–1090. [Google Scholar] [CrossRef]
- Seifertová, M.; Bryja, J.; Vyskočilová, M.; Martínková, N.; Šimokvá, A. Multiple Pleistocene refugia and post-glacial colonization in the European chub (Squalius cephalus) revealed by combined use of nuclear and mitochondrial markers. J. Biogeogr. 2012, 38, 1024–1040. [Google Scholar] [CrossRef]
- Ragauskas, A.; Butkauskas, D.; Prakas, P.; Gadliauskienė, K.; Gajduchenko, H.; Grauda, D. Complex phylogeographic relationships among the Eurasian perch (Perca fluviatilis) populations in the eastern part of the Baltic Sea Region. Hydrobiologia 2020, 847, 925–938. [Google Scholar] [CrossRef]
- Al Fatle, F.A.; Meleg, E.E.; Sallai, Z.; Szabo, G.; Varkonyi, E.; Urbanyi, B.; Kovacs, B.; Molnar, T.; Lehoczky, I. Genetic structure and diversity of native tench (Tinca tinca L. 1758) populations in Hungary—Establishment of basic knowledge base for a breeding program. Diversity 2022, 14, 336. [Google Scholar] [CrossRef]
- Bartáková, V.; Bryja, J.; Šanda, R.; Bektas, Y.; Stefanov, T.; Choleva, L.; Smith, C.; Reichard, M. High cryptic diversity of bitterling fish in the southern West Palearctic. Mol. Phylogenet. Evol. 2019, 133, 1–11. [Google Scholar] [CrossRef]
- Toomey, L.; Dellicour, S.; Vanina, T.; Pegg, J.; Kaczkowski, Z.; Kouřil, J.; Teletchea, F.; Bláha, M.; Fontaine, P.; Leco, T. Getting off on the right foot: Integration of spatial distribution of genetic variability for aquaculture development and regulations, the European perch case. Aquaculture 2020, 521, 734981. [Google Scholar] [CrossRef]
- Borovikova, E.; Nikulina, Y. The contact zone of phylogenetic lineages of freshwater fish in Arctic Eurasia: Genetic polymorphism of coregonid populations. Diversity 2023, 15, 163. [Google Scholar] [CrossRef]
- Fuller, N.; Smith, J.T.; Nagorskaya, L.L.; Gudkov, D.L.; Ford, A.T. Does Chernobyl-derived radiation impact the developmental stability of Asellus aquaticus 30 years on? Sci. Total Environ. 2017, 576, 242–250. [Google Scholar] [CrossRef]
- Nesbø, C.L.; Fossheim, T.; Vøllestad, L.; Jakobsen, K. Genetic divergence and phylogeographic relationships among European perch (Perca fluviatilis) populations reflect glacial refugia and postglacial colonization. Mol. Ecol. 1999, 8, 1387–1404. [Google Scholar] [CrossRef]
- Leliūna, E. Phenotype and Genotype Variation in Salmon (Salmo salar L.) and Sea Trout (S. trutta trutta L.) Populations of the Nemunas River Basin. PhD Thesis, Institute of Ecology of Vilnius University, Vilnius, Lithuania, 2008. [Google Scholar]
- Pimakhin, A.; Kouřil, J.; Stejskal, V.; Žák, J. The effect of geographical origin of perch (Perca fluviatilis L. 1758) populations on growth rates under natural and aquaculture conditions: A review. J. Appl. Ichthyol. 2015, 31, 56–63. [Google Scholar] [CrossRef]
- Olsson, J. Past and current trends of coastal predatory fish in the Baltic Sea with a focus on perch, pike, and pikeperch. Fishes 2019, 4, 7. [Google Scholar] [CrossRef]
- Hall, M.; Nordahl, O.; Larsson, P.; Forsman, A.; Tibblin, P. Intra-population variation in reproductive timing covaries with thermal plasticity of offspring performance in perch Perca fluviatilis. J. Anim. Ecol. 2021, 90, 2236–2347. [Google Scholar] [CrossRef] [PubMed]
- Vasemägi, A.; Ozerov, M.; Noreikiene, K.; López, M.E.; Gårdmark, A. Unlocking the genome of perch—From genes to ecology and back again. Ecol. Freshw. Fish 2023, 32, 677–702. [Google Scholar] [CrossRef]
- Fokina, O.; Grauda, D.; Rashal, I. The iPBS technique application for DNA fingerprinting of perch. Acta Biol. Univ. Daugavp. 2016, 16, 39–48. [Google Scholar]
- Christensen, A.F.; Skovrind, M.; Olsen, M.T.; Carl, H.; Gravlund, P.; Möller, P.R. Hatching success in brackish water of Perca fluviatilis eggs obtained from the western Baltic Sea. Int. J. Ichthyolo. 2016, 40, 133–138. [Google Scholar]
- Demandt, M.H. Temporal changes in genetic diversity of isolated populations of perch and roach. Conserv. Genet. 2010, 11, 249–255. [Google Scholar] [CrossRef]
- Yang, X.; Qian, L.; Wu, H.; Fan, Z.; Wang, C. Population differentiation, bottleneck and selection of Eurasian perch (Perca fluviatilis L.) at the Asian edge of its natural range. Biochem. Syst. Ecol. 2012, 40, 6–12. [Google Scholar] [CrossRef]
- Pukk, L.; Kuparinen, A.; Järv, L.; Gross, R.; Vasemägi, A. Genetic and life-history changes associated with fisheries-induced population collapse. Evol. Appl. 2013, 6, 749–760. [Google Scholar] [CrossRef]
- Ben Khadher, S. Analysis of Genetic Variability of Wild and Captive Population of Eurasian perch Perca fluviatilis, a Species under Domestication. Ph.D. Thesis, University of Lorraine, Nancy, France, 2015. [Google Scholar]
- Kokina, I.; Rubeniņa, I.; Bankovska, L.; Mickeviča, I.; Gavarāne, I. Case study of microsatellite polymorphism of European perch in selected commercially important lakes of Latvia. Biologia 2018, 73, 273–280. [Google Scholar] [CrossRef]
- Mankiewicz-Boczek, J.; Imsiridou, A.; Kaczkowski, Z.; Tsiora, A.; Karaiskou, N.; Łapińska, M.; Minos, G.; Zalewski, M. Genetic diversity of perch populations in three lowland reservoirs (Central Poland): Perspective for fish sustainable management. Pol. J. Ecol. 2013, 61, 385–390. [Google Scholar]
- Rolli, J.; Girardet, S.; Monachon, C.; Richard, C. Microsatellite analysis of perch (Perca fluviatilis) and its genetic authentication of geographical localization. Chimia 2014, 68, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Nesbø, C.L.; Magnhagen, C.; Jakobsen, K.S. Genetic differentiation among stationary and anadromous perch (Perca Fluviatilis) in the Baltic Sea. Hereditas 1998, 129, 241–249. [Google Scholar] [CrossRef]
- Faulks, L.; Svanbäck; Eklöv, P.R.; Östman, Ö. Genetic and morphological divergence along the littoral–pelagic axis in two common and sympatric fishes: Perch, Perca fluviatilis (Percidae) and roach, Rutilus rutilus (Cyprinidae). Biol. J. Linn. Soc. 2015, 114, 929–940. [Google Scholar] [CrossRef]
- Björklund, M.; Aho, T.; Behrmann-Godel, J. Isolation over 35 Years in a heated biotest basin causes selection on MHC class IIß genes in the European perch (Perca fluviatilis L.). Ecol. Evol. 2015, 5, 1440–1455. [Google Scholar] [CrossRef] [PubMed]
- Faulks, L.K.; Östman, Ö. Adaptive major histocompatibility complex (MHC) and neutral genetic variation in two native Baltic Sea fishes (Perch Perca fluviatilis and Zander Sander lucioperca) with comparisons to an introduced and disease susceptible population in Australia (P. fluviatilis): Assessing the risk of disease epidemics. J. Fish Biol. 2016, 88, 1564–1583. [Google Scholar] [CrossRef]
- Zhigileva, O.N.; Egorova, A.G. Genetic variability and phenotypic diversity in populations of the Eurasian perch, Perca fluviatilis (Actinopterygii, Percidae). Acta Biol. Sib. 2022, 8, 237–252. [Google Scholar]
- Nesbø, C.L.; Arab, M.O.; Jakobsen, K.S. Heteroplasmy, length and sequence variation in the mtDNA control regions of three percid fish species (Perca fluviatilis, Acerina cernua, Stizostedion lucioperca). Genetics 1998, 148, 1907–1919. [Google Scholar] [CrossRef]
- Refseth, U.H.; Nesbø, C.L.; Stacy, J.E.; Vøllestad, L.A.; Fjeld, E.; Jakobsen, K.S. Genetic evidence for different migration routes of freshwater fish into Norway revealed by analysis of current perch (Perca fluviatilis) populations in Scandinavia. Mol. Ecol. 1998, 7, 1015–1027. [Google Scholar] [CrossRef]
- Behrmann-Godel, J.; Gerlach, G.; Eckmann, R. Postglacial colonization shows evidence for sympatric population splitting of Eurasian perch (Perca fluviatilis L.) in Lake Constance. Mol. Ecol. 2004, 13, 491–497. [Google Scholar] [CrossRef]
- Behrmann-Godel, J.; Gerlach, G.; Eckmann, R. Kin and population recognition in sympatric Lake Constance perch (Perca fluviatilis L.): Can assortative shoaling drive population divergence? Behav. Ecol. Sociobiol. 2006, 59, 461–468. [Google Scholar] [CrossRef]
- Sruoga, A.; Butkauskas, D.; Rashal, I. Evaluation of genetic diversity of Perch (Perca fluviatilis) and Pikeperch (Sander lucioperca) populations from Curonian Lagoon and inshore waters of the Baltic Sea. Acta Biol. Univ. Daugavp. 2008, 8, 81–88. [Google Scholar]
- Bergek, S.; Björklund, M. Genetic and morphological divergence reveals local subdivision of perch (Perca fluviatilis L.). Biol. J. Linn. Soc. 2009, 96, 746–758. [Google Scholar] [CrossRef]
- Kalous, L.; Kuříková, P.; Kohout, J.; Rylková, K.; Petrtýl, M.; Čech, M. Differences in spatial communities of European perch (Perca fluviatilis Linnaeus, 1758) fry in a canyon-shaped reservoir are not attributable to genetics. J. Appl. Ichthyol. 2017, 33, 306–313. [Google Scholar] [CrossRef]
- Lin, G.-M.; Audira, G.; Juniardi, S.; Hsiao, C.-D. Metatranscriptomic analysis reveals co-expression pattern of mitochondrial oxidative phosphorylation (OXPHOS) genes among different species of bony fishes in muscle tissue. J. King Saud Univ. Sci. 2020, 32, 3084–3090. [Google Scholar] [CrossRef]
- Yang, R.; Ma, Z.; Beliczky, G.; Havasi, M.; Bercsenyi, M. The complete mitochondrial genome of perch Perca fluviatilis (Perciformes: Percidae). Mitochondrial DNA A DNA Mapp. Seq. Anal. 2016, 27, 1953–1954. [Google Scholar] [CrossRef]
- Satoh, T.P.; Miya, M.; Mabuchi, K.; Nishida, M. Structure and variation of the mitochondrial genome of fishes. BMC Genom. 2016, 17, 719. [Google Scholar] [CrossRef]
- Yang, X.; Wang, C.; Wang, J.; Ma, Y.; Yin, J.; Wu, H. Isolation and characterization of 12 polymorphic microsatellite loci in Eurasian perch (Perca fluviatilis L.). Conserv. Genet. Resour. 2009, 1, 229–231. [Google Scholar] [CrossRef]
- Pukk, L.; Kisand, V.; Ahmad, F.; Gross, R.; Vasemägi, A. Double-restriction-site-associated DNA (dRAD) approach for fast microsatellite marker development in Eurasian perch (Perca fluviatilis L.). Conserv. Genet. Resour. 2013, 6, 183–184. [Google Scholar] [CrossRef]
- Pukk, L.; Gross, R.; Vetemaa, M.; Vasemägi, A. Genetic discrimination of brackish and freshwater populations of Eurasian perch (Perca fluviatilis L.) in the Baltic Sea drainage: Implications for fish forensics. Fish. Res. 2016, 183, 155–164. [Google Scholar] [CrossRef]
- Xu, P.; Lu, C.; Sun, Z.; Kuang, Y.; Cao, D.; Huo, T.; Li, C.; Jin, H.; Zheng, X. In silico screening and development of microsatellite markers for genetic analysis in Perca fluviatilis. Animals 2022, 12, 1809. [Google Scholar] [CrossRef]
- Butkauskas, D.; Sruoga, A.; Ragauskas, A.; Kesminas, V.; Ložys, L.; Rashal, I.; Tzeng, W.N.; Žalakevičius, M. Investigations into genetic diversity of the perch inhabiting Ignalina nuclear power plant cooler and other inland water bodies of Lithuania on the basis of mtDNR analysis. Vet. ir Zootech. 2012, 60, 7–15. [Google Scholar]
- Jacobsen, M.W.; Pujolar, J.M.; Hansen, M.M. Relationship between amino acid changes in mitochondrial ATP6 and life-history variation in anguillid eels. Biol. Lett. 2015, 11, 20150014. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Wang, X.; Xu, S.; Gao, T.; Han, Z. Population genetic structure and selective pressure on the mitochondrial ATP6 gene of the Japanese sand lance Ammodytes personatus Girard. J. Mar. Biol. Assoc. UK 2019, 99, 1409–1416. [Google Scholar] [CrossRef]
- Shang, Y.; Wang, X.; Liu, G.; Wu, X.; Wei, Q.; Sun, G.; Mei, X.; Dong, Y.; Sha, W.; Zhang, H. Adaptability and evolution of gobiidae: A genetic exploration. Animals 2022, 12, 1741. [Google Scholar] [CrossRef] [PubMed]
- Nedveckaite, T.; Filistovic, V.; Marciulioniene, D.; Prokoptchuk, N.; Plukiene, R.; Gudelis, A.; Remeikis, V.; Yankovich, T.T.; Beresfordet, N.A. Background and anthropogenic radionuclide derived dose rates to freshwater ecosystem: Nuclear power plant cooling pond: Reference organisms. J. Environ. Radioact. 2011, 102, 788–795. [Google Scholar] [CrossRef]
- Barisevičiūtė, R.; Maceika, E.; Ežerinskis, Ž.; Šapolaitė, J.; Butkus, L.; Mažeika, J.; Rakauskas, V.; Juodis, L.; Steponėnas, A.; Druteikienė, R.; et al. Distribution of radiocarbon in sediments of the cooling pond of RBMK type Ignalina Nuclear Power Plant in Lithuania. PLoS ONE 2020, 15, e0237605. [Google Scholar] [CrossRef]
- Aljanabi, S.M.; Martinez, I. Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef]
- Rozen, S.; Skaletsky, H.J. Primer3 on the WWW for general users and for biologist programmers. In Bioinformatics Methods and Protocols: Methods in Molecular Biology; Misener, S., Krawetz, S.A., Eds.; Human Press: Totowa, NJ, USA, 2000; Volume 132, pp. 365–386. [Google Scholar]
- Villesen, P. FaBox: An online toolbox for FASTA sequences. Mol. Ecol. Notes 2007, 7, 965–968. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Kishino, H.; Taka-aki, Y. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987. [Google Scholar]
- Lynch, M.; Crease, T.J. The analysis of population survey data on DNA sequence variation. Mol. Biol. Evol. 1990, 7, 377–394. [Google Scholar] [CrossRef] [PubMed]
- Tajima, F. Evolutionary relationship of DNA sequences in finite populations. Genetics 1983, 105, 437–460. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin version 3.0, An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1, 47–50. [Google Scholar] [CrossRef]
- Nei, M. Genetic distance between populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Pichaud, N.; Ekström, A.; Breton, S.; Sundström, L.; Rowinski, P.; Blier, P.; Sandblom, E. Adjustments of cardiac mitochondrial phenotype in a warmer thermal habitat is associated with oxidative stress in European perch, Perca fluviatilis. Sci. Rep. 2020, 10, 17697. [Google Scholar] [CrossRef]
- Sruoga, A.; Rashal, I.; Butkauskas, D.; Ložys, L. Variety of mtDNA haplotypes in the populations of the European perch (Perca fluviatilis) of the Curonian lagoon, the coastal zone of the Baltic Sea and the Gulf of Riga. Proc. Latvian Acad. Sci. 2007, 61, 20–30. [Google Scholar]
- Gómez, A.; Lunt, D.H. Refugia within refugia: Patterns of phylogeographic concordance in the Iberian Peninsula. In Phylogeography of Southern European Refugia; Weiss, S., Ferrand, N., Eds.; Springer: Dordrecht, Germany, 2007; pp. 155–188. [Google Scholar]
- Laine, J.E. War in Europe: Health implications of environmental nuclear disaster amidst war. Eur. J. Epidemiol. 2022, 37, 221–225. [Google Scholar] [CrossRef]
- Hansen, M.M.; Olivieri, I.; Waller, D.M.; Nielsen, E.E.; The GeM Working Group. Monitoring adaptive genetic responses to environmental change. Mol. Ecol. 2012, 21, 1311–1329. [Google Scholar] [CrossRef]
- Fuller, N.; Ford, A.T.; Lerebours, A.; Gudkov, D.I.; Nagorskaya, L.L.; Smith, J.T. Chronic radiation exposure at Chernobyl shows no effect on genetic diversity in the freshwater crustacean, Asellus aquaticus thirty years on. Ecol. Evol. 2019, 9, 10135–10144. [Google Scholar] [CrossRef]
- Car, C.; Gilles, A.; Armant, O.; Burraco, P.; Beaugelin-Seiller, K.; Gashchak, S.; Camilleri, V.; Cavalié, I.; Laloi, P.; Adam-Guillermin, C.; et al. Unusual evolution of tree frog populations in the Chernobyl exclusion zone. Evol. Appl. 2020, 15, 203–219. [Google Scholar] [CrossRef]
- Uddin, A.; Mazum, T.H.; Barbhuiya, P.A.; Chakraborty, S. Similarities and dissimilarities of codon usage in mitochondrial ATP genes among fishes, aves, and mammals. IUBMB Life 2020, 72, 899–914. [Google Scholar] [CrossRef]
- Bergek, S.; Björklund. Cryptic barriers to dispersal within a lake allow genetic differentiation of Eurasian perch. Evolution 2007, 61, 1791–2041. [Google Scholar] [CrossRef]
- Jakubavičiūtė, E.; Pūtys, Ž.; Dainys, J.; Ložys, L. Perch (Perca fluviatilis) growth, mortality and stock exploitation by 40–45 mm mesh-sized gillnet fishery in the Curonian Lagoon. Acta Zool. Litu. 2011, 21, 215–220. [Google Scholar] [CrossRef]
- Souza, S.T.; Soukalová, K.; Děd, V.; Šmejkal, M.; Blabolil, P.; Říha, M.; Jůza, T.; Vašek, M.; Čech, M.; Peterka, J.; et al. Ontogenetic and interpopulation differences in otolith shape of the European perch (Perca fluviatilis). Fisher. Res. 2020, 230, 105673. [Google Scholar] [CrossRef]
- Rocha de Almeida, T.; Alix, M.; Le Cam, A.; Klopp, C.; Montfort, J.; Toomey, L.; Ledoré, Y.; Bobe, J.; Chardard, D.; Schaerlinger, B.; et al. Domestication may affect the maternal mRNA profile in unfertilized eggs, potentially impacting the embryonic development of Eurasian perch (Perca fluviatilis). PLoS ONE 2019, 14, e0226878. [Google Scholar] [CrossRef]
- Marić, S.; Razpet, A.; Nikolić, V.; Simonović. Genetic differentiation of European grayling (Thymallus thymallus) populations in Serbia, based on mitochondrial and nuclear DNA analyses. Genet. Sel. Evol. 2011, 43, 2. [Google Scholar] [CrossRef] [PubMed]
- Best, C.; Ikert, H.; Kostyniuk, D.J.; Craig, P.M.; Navarro-Martin, L.; Marandel, L.; Menningen, J.A. Epigenetics in teleost fish: From molecular mechanisms to physiological phenotypes. CBPB 2018, 224, 210–244. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.K.; Luikart, G.; Waples, R.S. Genetic monitoring as a promising tool for conservation and management. Trends Ecol. Evol. 2007, 22, 25–33. [Google Scholar] [CrossRef]
- Allendorf, F.W.; England, P.R.; Luikart, G.; Ritchie, P.A.; Ryman, N. Genetic effects of harvest on wild animal populations. Trends Ecol. Evol. 2008, 23, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Wang, W.; Huang, G.; Chen, Q.; Wei, G. Management of water resources assessment for nuclear power plants in China. Water Qual. Res. J. Can. 2015, 51, 282–295. [Google Scholar] [CrossRef]
- Kolar, M.V.; Gugleta, M. The consequences of disposal and leakage of radioactive materials on various species of marine and freshwater fish. Int. J. Fish Aquat. Stud. 2019, 7, 185–189. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Location | Basin | Sampling Time | Sample Size |
|---|---|---|---|---|
| Latvian samples | ||||
| 1 | Lake Engure North: 57.258, East: 23.117 Altitude 3.2 m, average depth 0.4 m, max depth 2.1 m, 4046 ha Eutrophic | Baltic Sea | 2015 | 19 |
| 2 | Lake Cirīšu North: 56.133, East: 26.966 Altitude ~140 m, average depth 4.1 m, max depth 10 m, 630.6 ha Eutrophic | Daugava River | 2011 | 21 |
| Lithuanian samples | ||||
| 3 | Lake Drūkšiai North: 55.621, East: 26.605 Altitude 141.6 m, average depth 7.6 m, max depth 33.3 m, 4487 ha Mesotrophic | Daugava River | 2009 | 22 |
| 4 | Lake Žeimenys North: 55.287, East: 26.06 Altitude 138.3 m, average depth 6.9 m, max depth 23.5 m, 443.8 ha Mesotrophic | Žeimena River | 2018 | 17 |
| 5 | Siesartis River North: 55.293, East: 24.909 Altitude 60 m | Siesartis River–Šventoji River | 2015 | 11 |
| 6 | Dotnuvėlė River (Akademijos Reservoir) North: 55.4, East: 23.85 Altitude 66.2 m, average depth 1.8 m, max depth 5.9 m, 35.4 ha Eutrophic | Dotnuvėlė River | 2014 | 14 |
| 7 | Curonian Lagoon North: 55.35, East: 21.197 Altitude 4 m, average depth 3.8 m, max depth 5.8 m, 158,400 ha Eutrophic | Baltic Sea | 2017 | 6 |
| 8 | Elektrėnai Reservoir North: 54.758, East: 24.669 Altitude 94.9 m, average depth 7.2 m, max depth 31 m, 1264 ha Eutrophic | Strėva River | 2017 | 8 |
| 9a | Neris River (Baltalaukis) North: 54.862, East: 25.401 Altitude 100 m | Neris River | 2009, 2010 | 12 |
| 9b | Neris River (Buivydžiai) North: 54.836, East: 25.734 Altitude 105 m | Neris River | 2018 | 18 |
| Belarusian samples | ||||
| 10a | Meleshkovichi River channel North: 51.91, East: 28.968 Altitude 100–120 m | Pripyat River | 2015 | 10 |
| 10b | Mozyr North: 52.06, East: 29.257 Altitude 120–140 m | Pripyat River | 2015 | 23 |
| 10c | Lake Aleksino North: 52.107, East: 28.744 Altitude 100–120 m | Pripyat River | 2015 | 2 |
| 10d | Berezina River North: 52.65, East: 29.772 Altitude ~116 m | Berezina River–Dnieper River | 2015 | 2 |
| Ukrainian samples | ||||
| 11a | Chernobyl cooling pond North: 51.385, East: 30.137 Altitude 110 m, average depth 6 m, max depth 18 m, 2290 ha Eutrophic | Pripyat River–Dnieper River | 2012 | 1 |
| 11b | Desna River North: 51.077, East: 30.889 Altitude ~111 m | Desna River | 2012 | 7 |
| Code—Sample | n | h | K | S | π |
|---|---|---|---|---|---|
| 1—Lake Engure | 19 | 0.667 ± 0.066 | 1.35673 | 4 | 0.00216 ± 0.00025 |
| 2—Lake Cirīšu | 21 | 0.562 ± 0.126 | 1.06667 | 6 | 0.00170 ± 0.00050 |
| 3—Lake Drūkšiai | 22 | 0.593 ± 0.089 | 0.89177 | 4 | 0.00142 ± 0.00035 |
| 4—Lake Žeimenys | 17 | 0.382 ± 0.113 | 0.38235 | 1 | 0.00061 ± 0.00018 |
| 5—Siesartis River | 11 | 0.764 ± 0.107 | 1.16364 | 5 | 0.00186 ± 0.00049 |
| 6—Dotnuvėlė River | 14 | 0.703 ± 0.095 | 1.56044 | 4 | 0.00249 ± 0.00037 |
| 7—Curonian Lagoon | 6 | 0.533 ± 0.172 | 1.06667 | 2 | 0.00170 ± 0.00055 |
| 8—Elektrėnai Reservoir | 8 | 0.607 ± 0.164 | 1.32143 | 3 | 0.00211 ± 0.00057 |
| 9—Neris River | 30 | 0.669 ± 0.060 | 1.16782 | 3 | 0.00186 ± 0.00028 |
| 10—Belarus | 37 | 0.611 ± 0.046 | 1.49550 | 3 | 0.00239 ± 0.00012 |
| 10a—Meleshkovichi River channel | 10 | 0 | 0 | 0 | 0 |
| 10b—Mozyr | 23 | 0.648 ± 0.055 | 1.47036 | 3 | 0.00235 ± 0.00022 |
| 11—Ukraine | 8 | 0.786 ± 0.113 | 2.03571 | 4 | 0.00325 ± 0.00055 |
| Overall a | 189 | 0.720 ± 0.021 | 1.37690 | 12 | 0.00220 ± 0.00011 |
| Overall b | 193 | 0.721 ± 0.021 | 1.37817 | 12 | 0.00220 ± 0.00011 |
| Code | 1 | 6 | 7 | 8 | 2 | 3 | 4 | 5 | 9 | 10a | 10b | 11 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 (I) | - | 0.653 | 0.601 | 0.706 | 0.015 | 0.002 | 0.001 | 0.020 | 0.050 | 0.001 | 0.014 | 0.263 |
| 6 (I) | −0.041 | - | 0.633 | 0.457 | 0.010 | 0.001 | 0.001 | 0.029 | 0.034 | 0.001 | 0.026 | 0.354 |
| 7 (I) | −0.047 | −0.077 | - | 0.591 | 0.014 | 0.001 | 0.001 | 0.009 | 0.021 | 0.001 | 0.019 | 0.272 |
| 8 (I) | −0.064 | −0.033 | −0.005 | - | 0.116 | 0.014 | 0.009 | 0.081 | 0.054 | 0.001 | 0.067 | 0.387 |
| 2 (II) | 0.156 | 0.208 | 0.323 | 0.080 | - | 0.311 | 0.390 | 0.614 | 0.475 | 0.001 | 0.066 | 0.097 |
| 3 (II) | 0.262 | 0.298 | 0.467 | 0.228 | 0.005 | - | 0.647 | 0.733 | 0.246 | 0.001 | 0.095 | 0.047 |
| 4 (II) | 0.331 | 0.389 | 0.641 | 0.344 | −0.001 | −0.026 | - | 0.880 | 0.110 | 0.001 | 0.021 | 0.010 |
| 5 (II) | 0.190 | 0.224 | 0.377 | 0.133 | −0.025 | −0.037 | −0.049 | - | 0.431 | 0.001 | 0.100 | 0.110 |
| 9 (II) | 0.097 | 0.132 | 0.257 | 0.190 | −0.011 | 0.015 | 0.055 | −0.012 | - | 0.001 | 0.132 | 0.242 |
| 10a (III) | 0.505 | 0.480 | 0.682 | 0.646 | 0.653 | 0.743 | 0.900 | 0.752 | 0.607 | - | 0.001 | 0.007 |
| 10b (III) | 0.164 | 0.159 | 0.271 | 0.143 | 0.081 | 0.066 | 0.156 | 0.077 | 0.035 | 0.498 | - | 0.454 |
| 11 (III) | 0.023 | 0.003 | 0.052 | −0.001 | 0.101 | 0.157 | 0.269 | 0.112 | 0.031 | 0.383 | −0.032 | - |
| Haplotype | Codons | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ATP6 | cox3 | ||||||||||||||||
| 19 | 34 | 41 | 59 | 68 | 70 | 75 | 78 | 84 | 106 | 114 | 121 | 125 | 155 | 171 | 202 | 208 | |
| A | ATC I | GGC G | CTA L | TAT Y | TCC S | AAT N | GTT V | TGA W | ATT I | CTA L | GAA E | CGC R | TTG L | CCA P | TTA L | GCA A | CCC P |
| A1 | AAC N | ||||||||||||||||
| A2 | CTG L | ||||||||||||||||
| A3 | TGG W | ||||||||||||||||
| B | CTC L | CTA L | |||||||||||||||
| B1 | CTC L | GAG E | CTA L | ||||||||||||||
| C | GGT G | ||||||||||||||||
| C1 | GGT G | GTC V | |||||||||||||||
| C2 | GGT G | TAC Y | |||||||||||||||
| D | CTT L | CTA L | |||||||||||||||
| D1 | GTT V | CTT L | CTA L | ||||||||||||||
| D2 | ATT I | CTT L | CTA L | ||||||||||||||
| D3 | CTT L | CTA L | CCA P | ||||||||||||||
| New1 AP005995 | CTT L | TTA L | |||||||||||||||
| New2 KM410088 | TTA L | TCT S | CGA R | CCC P | |||||||||||||
| Genetic Group | n | dS/dN | |
|---|---|---|---|
| ATP6 Region (627 bp) | ATP6 Partial (589 bp) | ||
| I | 47 | 0.088768 | 0 |
| II | 101 | 0 | 0 |
| III | 45 | 0.089767 | 0.090444 |
| Total | 193 | 0.065331 | 0.03314 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ragauskas, A.; Ignatavičienė, I.; Rakauskas, V.; Grauda, D.; Prakas, P.; Butkauskas, D. Trends of Eurasian Perch (Perca fluviatilis) mtDNA ATP6 Region Genetic Diversity within the Hydro-Systems of the Eastern Part of the Baltic Sea in the Anthropocene. Animals 2023, 13, 3057. https://doi.org/10.3390/ani13193057
Ragauskas A, Ignatavičienė I, Rakauskas V, Grauda D, Prakas P, Butkauskas D. Trends of Eurasian Perch (Perca fluviatilis) mtDNA ATP6 Region Genetic Diversity within the Hydro-Systems of the Eastern Part of the Baltic Sea in the Anthropocene. Animals. 2023; 13(19):3057. https://doi.org/10.3390/ani13193057
Chicago/Turabian StyleRagauskas, Adomas, Ieva Ignatavičienė, Vytautas Rakauskas, Dace Grauda, Petras Prakas, and Dalius Butkauskas. 2023. "Trends of Eurasian Perch (Perca fluviatilis) mtDNA ATP6 Region Genetic Diversity within the Hydro-Systems of the Eastern Part of the Baltic Sea in the Anthropocene" Animals 13, no. 19: 3057. https://doi.org/10.3390/ani13193057
APA StyleRagauskas, A., Ignatavičienė, I., Rakauskas, V., Grauda, D., Prakas, P., & Butkauskas, D. (2023). Trends of Eurasian Perch (Perca fluviatilis) mtDNA ATP6 Region Genetic Diversity within the Hydro-Systems of the Eastern Part of the Baltic Sea in the Anthropocene. Animals, 13(19), 3057. https://doi.org/10.3390/ani13193057