
To Flee or Not to Flee: How Age, Reproductive Phase, and Mate Presence Affect White Stork Flight Decisions

 , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
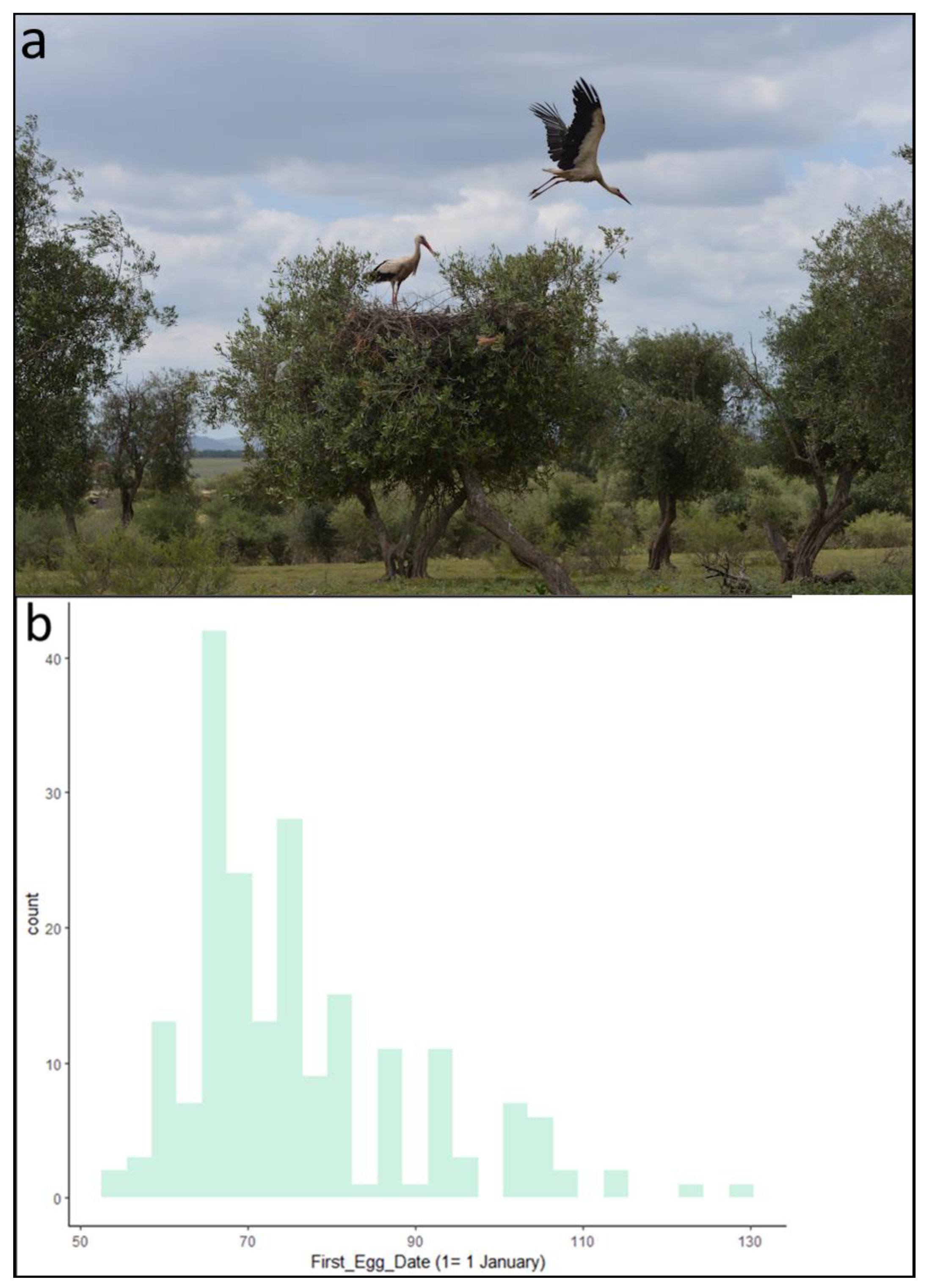
2.2. Data Collection
2.3. Statistical Analysis
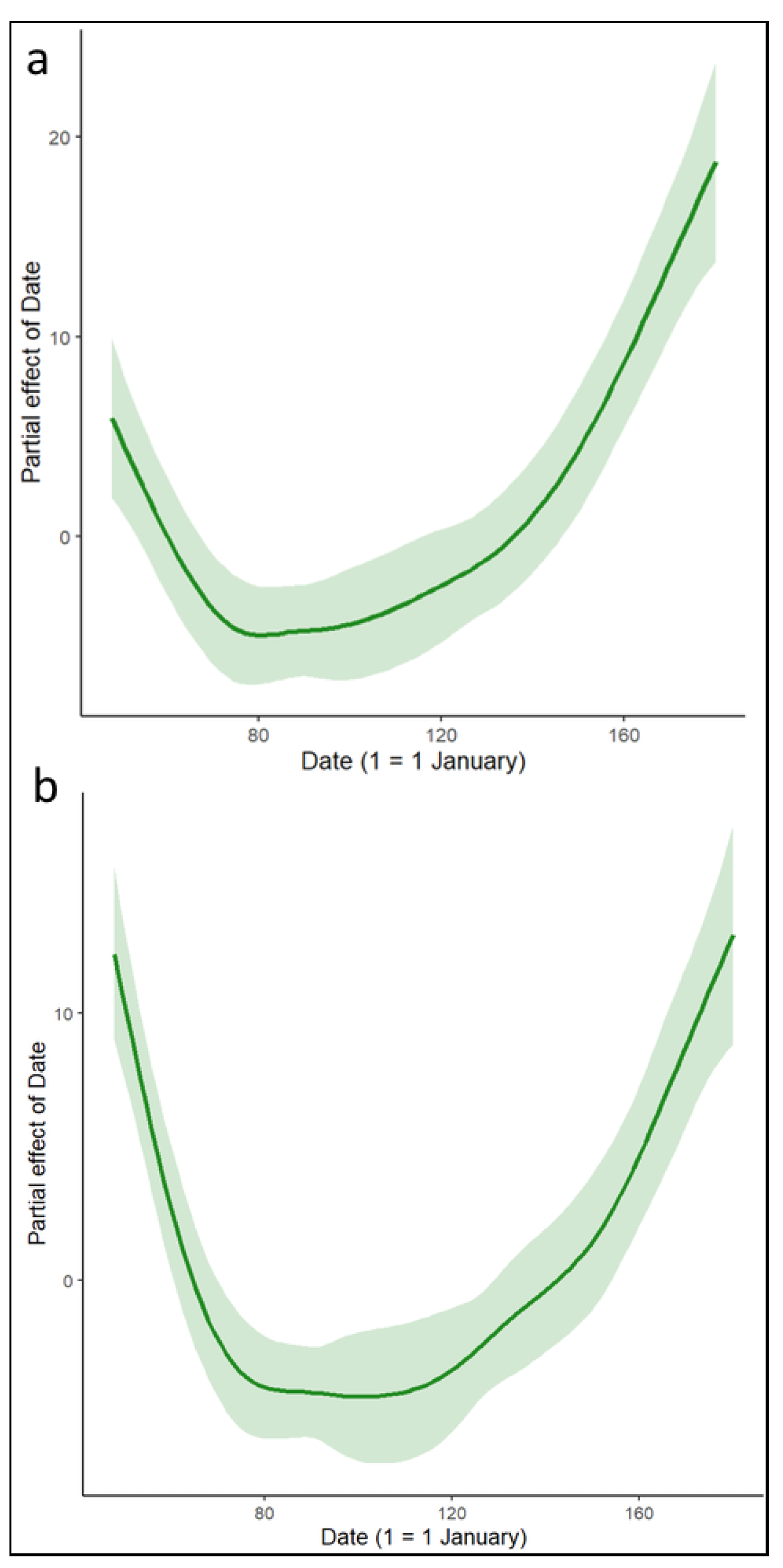
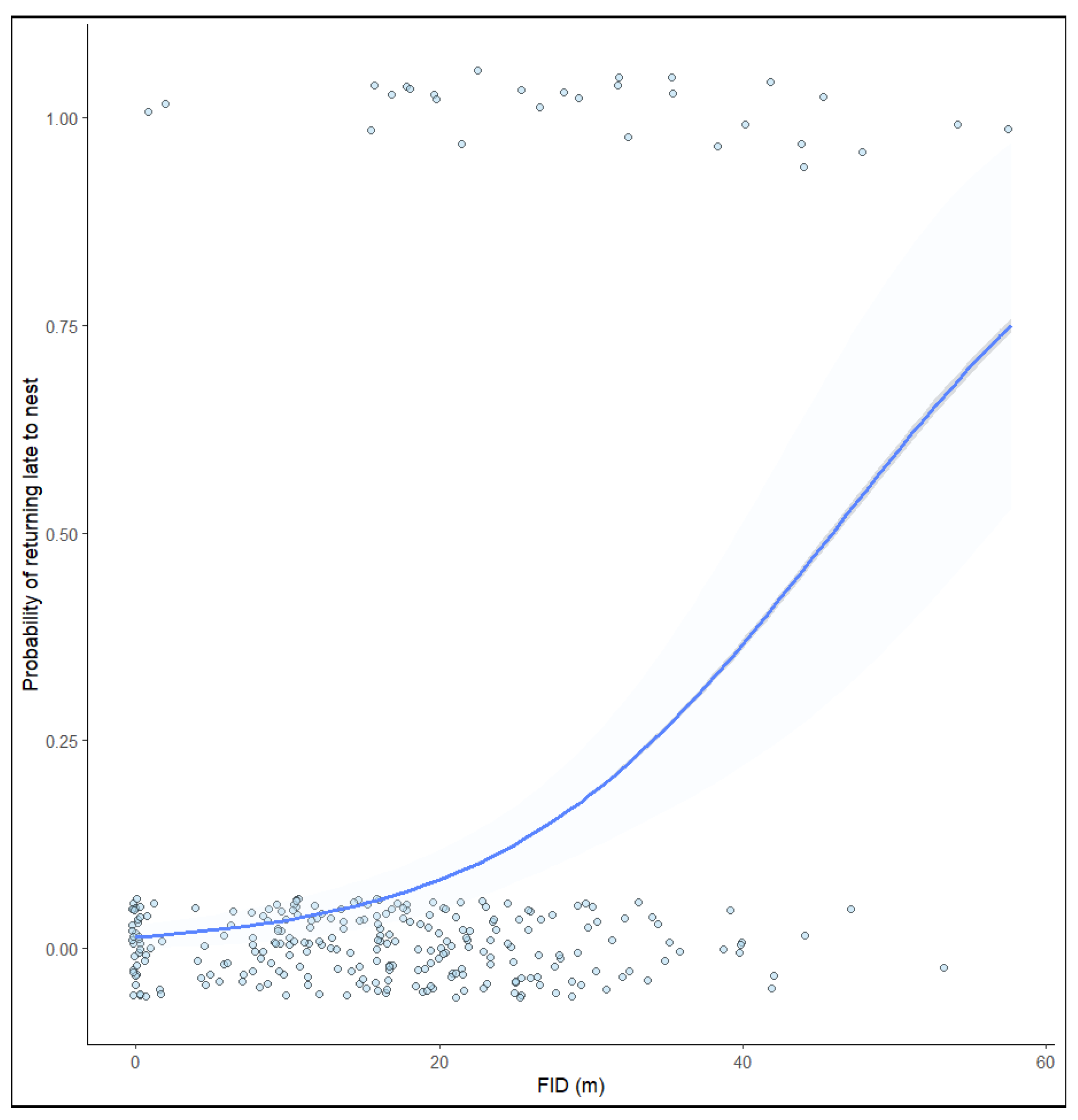
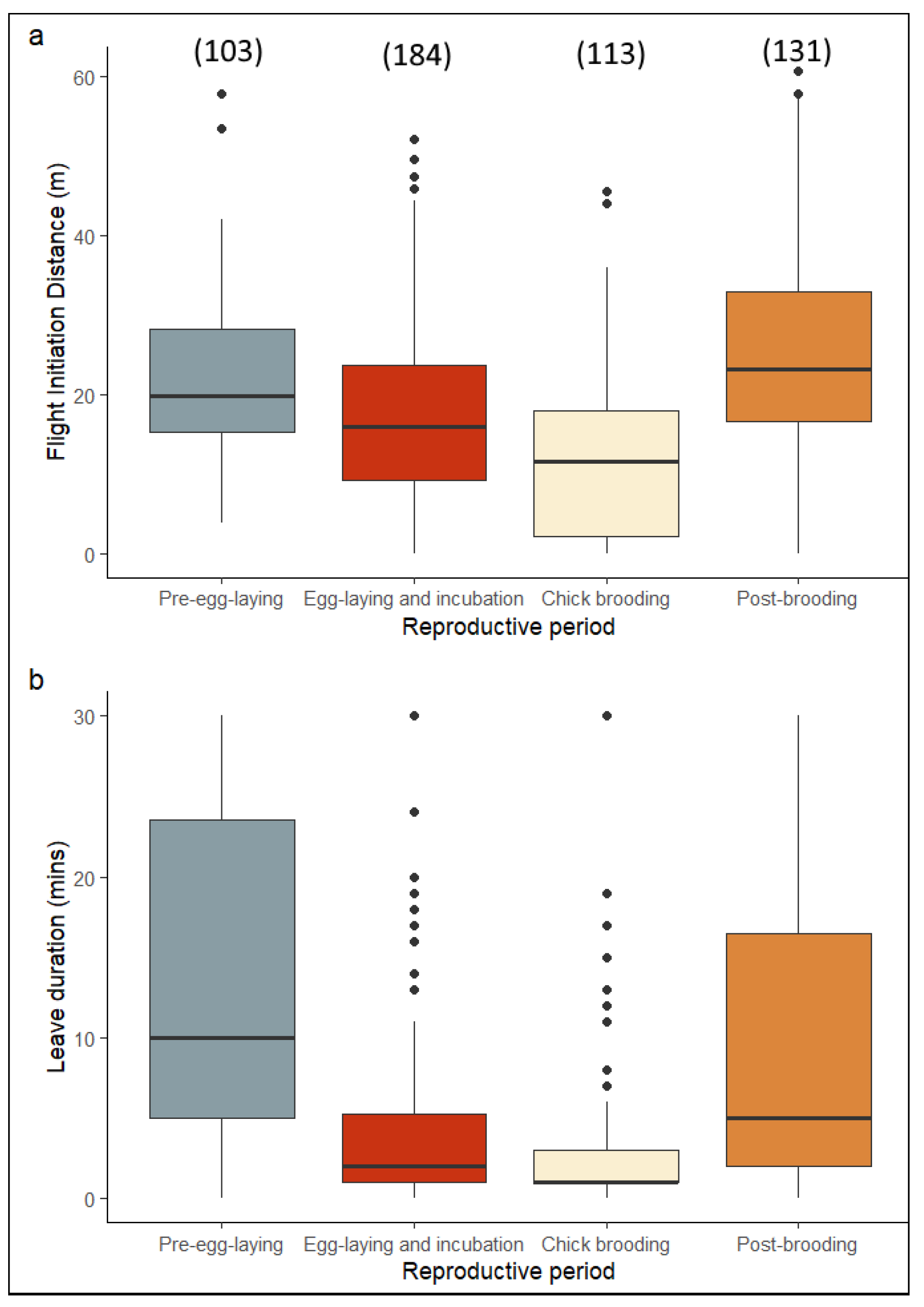
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ricklefs, R.E. An analysis of nesting mortality in birds. Smithson. Contrib. Zool. 1969, 9, 1–48. [Google Scholar] [CrossRef]
- Little, I.T.; Hockey, P.A.R.; Jansen, R. Predation drives nesting success in moist highland grasslands: The importance of maintaining vegetation cover for bird conservation. Ostrich 2015, 86, 97–111. [Google Scholar] [CrossRef]
- Trivers, R.L. Parental investment and sexual selection. In Sexual Selection and the Descent of Man 1871–1971; Campbell, B., Ed.; Aldine: Chicago, IL, USA, 1972; pp. 136–179. [Google Scholar]
- Clutton-Brock, T.H. The Evolution of Parental Care; Princeton University Press: Princeton, NJ, USA, 1991. [Google Scholar] [CrossRef]
- Rytkönen, S. Nest defence in great tits Parus major: Support for parental investment theory. Behav. Ecol. Sociobiol. 2002, 52, 379–384. [Google Scholar] [CrossRef]
- Sergio, F.; Bogliani, G.; Bildstein, K. Nest defense as parental care in the Northern Hobby (Falco subbuteo). The Auk 2001, 118, 1047–1052. [Google Scholar] [CrossRef][Green Version]
- Tryjanowski, P.; Golawski, A. Sex differences in nest defence by the red-backed shrike Lanius collurio: Effects of offspring age, brood size, and stage of breeding season. J. Ethol. 2004, 22, 13–16. [Google Scholar] [CrossRef]
- Regelmann, K.; Curio, E. Determinants of brood defence in the great tit Parus major L. Behav. Ecol. Sociobiol. 1983, 13, 131–145. [Google Scholar] [CrossRef]
- Pavel, S.; Bures, S. Nest defence in the meadow pipit Anthus pratensis: The influence of renesting potential. J. Ethol. 2008, 26, 367–373. [Google Scholar] [CrossRef]
- Andersson, M.; Wicklund, C.G.; Rundgren, H. Parental defence of offspring: A model and an example. Anim. Behav. 1980, 28, 536–542. [Google Scholar] [CrossRef]
- Curio, E.; Regelmann, K.; Zimmermann, U. The defence of first and second broods by great tit (Parus major): A test of predictive sociobiology. Z. Tierpsychol. 1984, 66, 101–127. [Google Scholar] [CrossRef]
- Curio, E.; Regelmann, K.; Zimmermann, U. Brood defence in the great tit (Parus major): The influence of life-history and habitat. Behav. Ecol. Sociobiol. 1985, 16, 273–283. [Google Scholar] [CrossRef]
- Wallin, K. Defence as parental care in tawny owls (Strixaluco). Behaviour 1987, 102, 213–230. [Google Scholar] [CrossRef]
- Weatherhead, P.J.; Robertson, R.J. Sexual recognition and anticuckoldry behavior in savannah sparrows. Can. J. Zool. 1980, 58, 991–996. [Google Scholar] [CrossRef]
- Dowling, L.; Bonier, F. Should I stay, or should I go: Modeling optimal flight initiation distance in nesting birds. PLoS ONE 2018, 13, e0208210. [Google Scholar] [CrossRef]
- Ydenberg, R.C.; Dill, M. The economics of fleeing from predators. Adv. Study Behav. 1986, 16, 229–249. [Google Scholar] [CrossRef]
- Cooper, W.E.; Frederick, W.G. Optimal flight initiation distance. J. Theor. Biol. 2007, 244, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Blumstein, D.T.; Samia, D.S.M.; Cooper, W.E., Jr. Escape behavior: Dynamic decisions and a growing consensus. Curr. Opin. Behav. Sci. 2016, 12, 24–29. [Google Scholar] [CrossRef]
- Ehrlich, P.R.; Ehrlich, A.H. Can a collapse of global civilization be avoided? Proc. R. Soc. B 2013, 280, 29122845. [Google Scholar] [CrossRef]
- Vieira, P. Is overpopulation a growth? The pathology of permanent expansion. Oxf. Lit. Rev. 2016, 38, 67–83. [Google Scholar] [CrossRef]
- Cresswell, W. Non-lethals effects of predation in birds. IBIS 2008, 150, 3–17. [Google Scholar] [CrossRef]
- Hall, M.J.; Burns, A.L.; Martin, J.M.; Hochuli, D.F. Flight initiation distance changes across landscapes and habitats in a successful urban coloniser. Urban Ecosyst. 2020, 23, 785–791. [Google Scholar] [CrossRef]
- Gill, J.A. Approaches to measuring the effects of human disturbance on birds. IBIS 2007, 149, 9–14. [Google Scholar] [CrossRef]
- Cooper, W.E., Jr. Dynamic risk assessment: Prey rapidly adjust flight initiation distance to changes in predator approach speed. Ethology 2006, 112, 858–864. [Google Scholar] [CrossRef]
- Fouzari, A.; Samraoui, F.; Alfarhan, A.H.; Samraoui, B. Nesting ecology of Ferruginous Duck Aythya nyroca in north-eastern Algeria. Afr. Zool. 2015, 50, 299–305. [Google Scholar] [CrossRef]
- Palmer, A.G.; Nordmeyer, D.L.; Roby, D.D. Factors influencing nest attendance and time-activity budgets of Peregrine Falcons in interior Alaska. Arctic 2001, 54, 105–114. [Google Scholar] [CrossRef]
- Schmidt, K.A.; Whelan, C.J. Quantifying male wood thrush nest-attendance and its relationship to nest success. Condor 2005, 107, 138–144. [Google Scholar] [CrossRef]
- Stien, J.; Ims, R.A. Absence from the nest due to human disturbance induces higher nest predation risk than natural recesses in Common eiders Somateria mollissima. IBIS 2016, 158, 249–260. [Google Scholar] [CrossRef]
- Djelailia, A.; Baaziz, N.; Samraoui, F.; Alfarhan, A.H.; Samraoui, B. Distribution and breedingecology of the Ferruginous Duck Aythya nyroca in Algeria. Ostrich 2017, 89, 5–12. [Google Scholar] [CrossRef]
- Flegeltaub, M.; Biro, P.A.; Beckmann, C. Avian nest abandonment prior to laying—A strategy to minimize predation risk? J. Ornithol. 2017, 158, 1091–1098. [Google Scholar] [CrossRef]
- Verboven, N.; Tinbergen, J.M. Nest desertion: A trade-off between current and future reproduction. Anim. Behav. 1971, 63, 951–958. [Google Scholar] [CrossRef]
- Soldatini, C.; Albores-Barajas, Y.V.; Tagliavia, M.; Massa, B.; Fusani, L.; Canoine, V. Effects of human disturbance on cave-nesting seabirds: The case of the storm petrel. Conserv. Physiol. 2015, 3, cov041. [Google Scholar] [CrossRef]
- Doherty, T.S.; Hays, G.C.; Driscoll, D.A. Human disturbance causes widespread disruption of animal movement. Nat. Ecol. Evol. 2021, 5, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Burger, J. Effects of human disturbance on colonial species, particularly gulls. Col. Waterbirds 1981, 4, 28–36. [Google Scholar] [CrossRef]
- Watson, H.; Bolton, M.; Monaghan, P. Out of sight but not out of harm’s way: Human disturbance reduces reproductive success of a cavity-nesting seabird. Biol. Conserv. 2014, 174, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, B.; Chown, S.L. A meta-analysis of human disturbance impacts on Antarctic wildlife: Human disturbance impacts on Antarctic wildlife. Biol. Rev. 2016, 91, 578–596. [Google Scholar] [CrossRef]
- Trivers, R.L. Parent-offspring conflict. Am. Zool. 1974, 14, 249–264. [Google Scholar] [CrossRef]
- Wade, M.; Shuster, S.M. The evolution of parental care in the context of sexual selection: A critical reassessment of parental investment theory. Am. Nat. 2002, 160, 285–292. [Google Scholar] [CrossRef][Green Version]
- Montgomerie, R.D.; Weatherhead, P.J. Risks and rewards of nest defence by parent birds. Q. Rev. Biol. 1988, 63, 167–187. [Google Scholar] [CrossRef]
- Zbyryt, A.; Dylewski, L.; Morelli, F.; Sparks, T.H.; Tryjanowski, P. Behavioural responses of adult and young White Storks Ciconia ciconia in nests to an unmanned aerial vehicle. Acta Ornithol. 2021, 55, 243–251. [Google Scholar] [CrossRef]
- Belabed, B.-E.; Athamnia, M.; Touati, L.; Samraoui, F.; Boucheker, A.; Samraoui, B. The early bird catches the worm: Age-specific arrival time influences reproductive performance in the White Stork Ciconia ciconia. Bird Study 2019, 66, 121–129. [Google Scholar] [CrossRef]
- Athamnia, M.; Belabed, B.-E.; Samraoui, K.R.; Boucheker, A.; Touati, L.; Samraoui, F.; El-Serehy, H.A.; Samraoui, B. Variability in arrival tile of White Storks (Ciconia ciconia L.): Impact of age, interindividual variation, and global change. Front. Ecol. Evol. 2022, 10, 841744. [Google Scholar] [CrossRef]
- R Development Core Team. R a Language and Environment for Statistical Computing; R Development Core Team: Vienna, Austria, 2022. [Google Scholar]
- Dale, S.; Gustavsen, R.; Slagsvold, T. Risk taking during parental care: A test of three hypotheses applied to the pied flycatcher. Behav. Ecol. Sociobiol. 1996, 39, 31–42. [Google Scholar] [CrossRef]
- Fulin, M.; Jerzak, L.; Sparks, T.H.; Tryjanowski, P. Relationship between arrival date, hatching date and breeding success of the white stork (Ciconia ciconia) in Slovakia. Biologia 2009, 64, 361–364. [Google Scholar] [CrossRef][Green Version]
- Collins, S.A.; Giffin, G.J.; Strong, W.T. Using flight initiation distance to evaluate responses of colonial-nesting Great Egrets to the approach of an unmanned aerial vehicle. J. Field Ornithol. 2019, 90, 382–390. [Google Scholar] [CrossRef]
- Redmond, L.J.; Eroh, C.; Haq, J.I.; Varra, D. The effect of offspring value on variation of nest defense intensity in gray catbirds (Dumetella carolinensis). Behav. Ecol. Sociobiol. 2020, 74, 3. [Google Scholar] [CrossRef]
- Onnebrink, H.; Curio, E. Brood defense and age of young: A test of the vulnerability hypothesis. Behav. Ecol. Sociobiol. 1991, 29, 61–68. [Google Scholar] [CrossRef]
- Redondo, T. Avian nest defence: Theoretical models and evidence. Behaviour 1989, 111, 161–195. [Google Scholar]
- Pugesek, B.H. The relationship between parental age and reproductive effort in the California gull (Larus californicus). Behav. Ecol. Sociobiol. 1983, 13, 161–171. [Google Scholar] [CrossRef]
- Santos, E.S.A.; Nakagawa, S. The costs of parental care: A meta-analysis of the trade-off between parental effort and survival in birds. J. Evol. Biol. 2012, 25, 1911–1917. [Google Scholar] [CrossRef]
- Møller, A.P.; Nielsen, J.T. Parental defense of offspring and life history of a long-lived raptor. Behav. Ecol. 2014, 25, 1505–1512. [Google Scholar] [CrossRef]
- Arnqvist, G.; Rowe, L. Sexual Conflict; Princeton University Press: Princeton, NJ, USA, 2005. [Google Scholar] [CrossRef]
- Burger, J. Pattern, mechanism, and adaptive significance of territoriality in Herring Gulls (Larus argentatus). Ornithol. Monogr. 1984, 34, 1–92. [Google Scholar]
- Kilpi, M. Do Herring Gulls (Larus argentatus) invest more in offspring defence as the breeding season advances? Ornis Fenn. 1987, 64, 16–20. [Google Scholar]
- Harrison, F.; Barta, Z.; Cuthill, I.; Székely, T. How is sexual conflict over parental care resolved? A meta-analysis. J. Evol. Biol. 2009, 22, 1800–1812. [Google Scholar] [CrossRef] [PubMed]
- Székely, T. Sexual conflict between parents: Offspring desertion and asymmetrical parental care. Cold Spring Harb. Perspect. Med. 2014, 6, a017665. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, S.; Barba, E. Nestling growth is impaired by heat stress: An experimental study in a Mediterranean Great Tit population. Zool. Stud. 2016, 55, e40. [Google Scholar] [CrossRef]
- Bohler, M.W.; Chowdury, V.S.; Cline, M.A.; Gilbert, E.R. Heat stress responses in birds: A review of the neural components. Biology 2021, 10, 1095. [Google Scholar] [CrossRef] [PubMed]
- Gillett, W.H.; Hayward, J.L., Jr.; Stout, J.F. Effects of human activity on egg and chick mortality in a Glaucous-winged Gull colony. Condor 1975, 77, 492–495. [Google Scholar] [CrossRef]
- Tremblay, J.; Ellison, L.N. Effects of human disturbance on breeding of Black-crowned Night Herons. The Auk 1979, 96, 364–369. [Google Scholar]
- Cairns, D. Nesting density, habitat structure and human disturbance as factors in Black Guillemot reproduction. Wilson Bull. 1980, 92, 352–361. [Google Scholar]
- Gotanda, K.M. Human influences on antipredator behavior in Darwin’s finches. Anim. Ecol. 2019, 89, 614–622. [Google Scholar] [CrossRef]
- Møller, A.P.; Ibañez-Álamo, J.D. Escape behaviour of birds provides evidence of predation being involved in urbanization. Anim. Behav. 2012, 84, 341–348. [Google Scholar] [CrossRef]
- Møller, A.P.; Grim, T.; Ibáñez-Alamo, J.D.; Markó, G.; Tryjanowski, P. Change in flight initiation distance between urban and rural habitats following a cold winter. Behav. Ecol. 2013, 24, 1211–1217. [Google Scholar] [CrossRef]
- Beaugeard, E.; Brischoux, F.; Henry, P.-Y.; Parenteau, C.; Trouvé, C.; Angelier, F. Does urbanization cause stress in wild birds during development? Insights from feather corticosterone levels in juvenile house sparrows (Passer domesticus). Ecol. Evol. 2019, 9, 640–652. [Google Scholar] [CrossRef] [PubMed]
- Weaver, D.B.; Lawton, L.J. Twenty years on: The state of contemporary ecotourism research. Tour. Manag. 2007, 28, 1168–1179. [Google Scholar] [CrossRef]
- Wheeller, B. Tourism’s troubled times: Responsible tourism is not the answer. Tour. Manag. 1991, 12, 91–96. [Google Scholar] [CrossRef]
- Blumstein, D.T.; Anthony, L.; Harcourt, R.; Ross, G.A. Testing a key assumption of wildlife buffer zones: Is flight initiation distance a species-specific trait? Biol. Conserv. 2003, 110, 97–100. [Google Scholar] [CrossRef]
- Whitfield, P.; Rae, R. Human disturbance of breeding Wood Sandpipers Tringaglareola: Implications for “alert distances” in prescribing protective buffer zones. Ornis Fenn. 2014, 91, 57–66. [Google Scholar]
- Braimoh, B.; Iwajomo, S.; Wilson, M.; Chaskda, A.; Ajang, A.; Cresswell, W. Managing human disturbance: Factors influencing flight-initiation distance of birds in a West African nature reserve. Ostrich 2018, 89, 59–69. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Estimate | SE | t-Value | p-Value |
|---|---|---|---|---|
| Flight Initiation Distance | ||||
| Parametric Coefficients | ||||
| Intercept | 12.26 | 3.4 | 3.61 | 0.0004 |
| Age | −1.36 | 0.6 | −2.26 | 0.0245 |
| No. of Individuals | 8.27 | 1.3 | 6.36 | 8.10 × 10−10 |
| edf | Ref.df | Chi.sq | p-Value | |
| Smooth Terms | ||||
| Date | 4.64 | 4.64 | 24.56 | <2× 10−16 |
| Leave Duration | ||||
| Parametric Coefficients | ||||
| Intercept | 4.88 | 2.32 | 2.11 | 0.0362 |
| Age | −0.53 | 0.37 | −1.43 | 0.1527 |
| No. of Individuals | 3.62 | 1.05 | 3.45 | 0.0007 |
| edf | Ref.df | Chi.sq | p-Value | |
| Smooth Terms | ||||
| Date | 5.49 | 5.49 | 26.47 | <2× 10−16 |
| Predictor | Odds Ratio | 95% CI Lower Bound | 95% CI Upper Bound | Std. Error | z-Value | Pr |
|---|---|---|---|---|---|---|
| Intercept | 0.014 | 0.005 | 0.035 | 0.518 | −8.217 | <2 × 10−16 |
| FID | 1.097 | 1.05 | 1.148 | 0.018 | 5.191 | 2.09 × 10−7 |
| Parameter | Estimate | Se | t-Value | p-Value |
|---|---|---|---|---|
| FID | ||||
| Parametric Coefficients | ||||
| Intercept | 29.27 | 2.99 | 9.78 | <2 × 10−16 |
| Egg-Laying and Incubation | −5.28 | 1.33 | −3.98 | 7.80 × 10−5 |
| Chick Brooding | −7.35 | 1.63 | −4.51 | 8.20 × 10−6 |
| Post-Brooding | 7.2 | 1.62 | 4.45 | 1.06 × 10−5 |
| Age | −2.31 | 0.55 | −4.18 | 3.45 × 10−5 |
| No. of Individuals | 5.46 | 1.18 | 4.61 | 5.10 × 10−6 |
| Nest Recess | ||||
| Intercept | 11.41 | 1.78 | 6.39 | 3.65 × 10−10 |
| Egg-Laying and Incubation | −7.83 | 1.09 | −7.15 | 2.97 × 10−12 |
| Chick Brooding | −7.84 | 1.34 | −5.86 | 8.18 × 10−9 |
| Post-Brooding | −1.26 | 1.32 | −0.96 | 0.339 |
| Age | −0.18 | 0.28 | −0.63 | 0.532 |
| No. of Individuals | 3.82 | 0.94 | 4.05 | 5.97 × 10−5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Touati, L.; Athamnia, M.; Boucheker, A.; Belabed, B.-E.; Samraoui, F.; Alfarhan, A.H.; Møller, A.P.; Samraoui, B. To Flee or Not to Flee: How Age, Reproductive Phase, and Mate Presence Affect White Stork Flight Decisions. Animals 2023, 13, 2920. https://doi.org/10.3390/ani13182920
Touati L, Athamnia M, Boucheker A, Belabed B-E, Samraoui F, Alfarhan AH, Møller AP, Samraoui B. To Flee or Not to Flee: How Age, Reproductive Phase, and Mate Presence Affect White Stork Flight Decisions. Animals. 2023; 13(18):2920. https://doi.org/10.3390/ani13182920
Chicago/Turabian StyleTouati, Laïd, Mohamed Athamnia, Abdennour Boucheker, Bourhane-Edinne Belabed, Farrah Samraoui, Ahmed H. Alfarhan, Anders P. Møller, and Boudjéma Samraoui. 2023. "To Flee or Not to Flee: How Age, Reproductive Phase, and Mate Presence Affect White Stork Flight Decisions" Animals 13, no. 18: 2920. https://doi.org/10.3390/ani13182920
APA StyleTouati, L., Athamnia, M., Boucheker, A., Belabed, B.-E., Samraoui, F., Alfarhan, A. H., Møller, A. P., & Samraoui, B. (2023). To Flee or Not to Flee: How Age, Reproductive Phase, and Mate Presence Affect White Stork Flight Decisions. Animals, 13(18), 2920. https://doi.org/10.3390/ani13182920