
Comparative Label-Free Liquid Chromatography–Mass Spectrometry Milk Proteomic Profiles Highlight Putative Differences between the Autochthon Teramana and Saanen Goat Breeds

 , , ,
, , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Sample Preparation
2.3. Filter-Aided Sample Preparation (FASP)
2.4. Mass Spectrometry for Label-Free LC–MS
2.5. Label-Free LC–MS Quantitative Profiling
2.6. Functional and Protein Network Analyses
3. Results
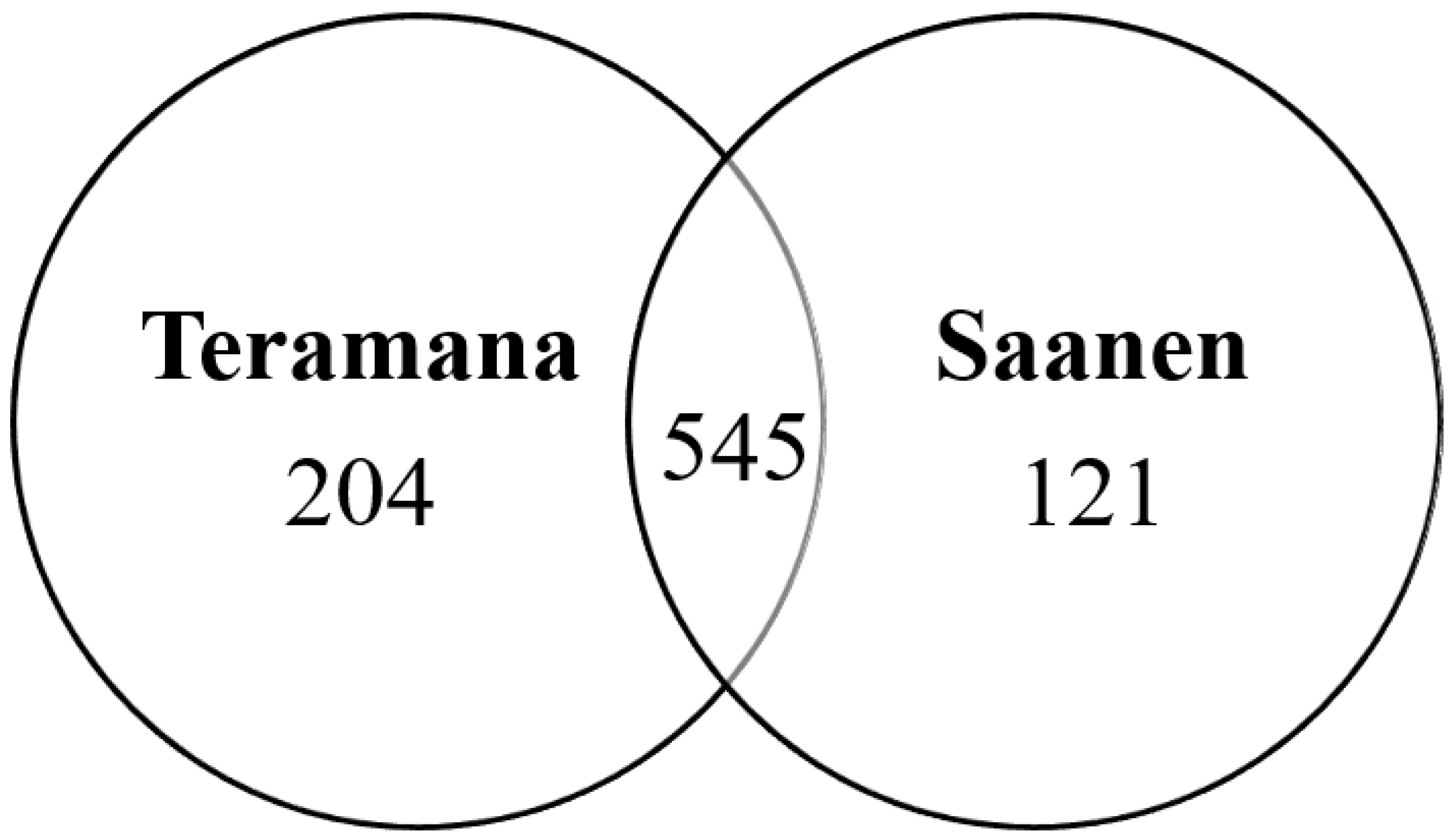
3.1. Identification of Milk Proteins in Goat Breeds
3.2. Label-Free Quantitative Proteomic Analysis of Goat Milk
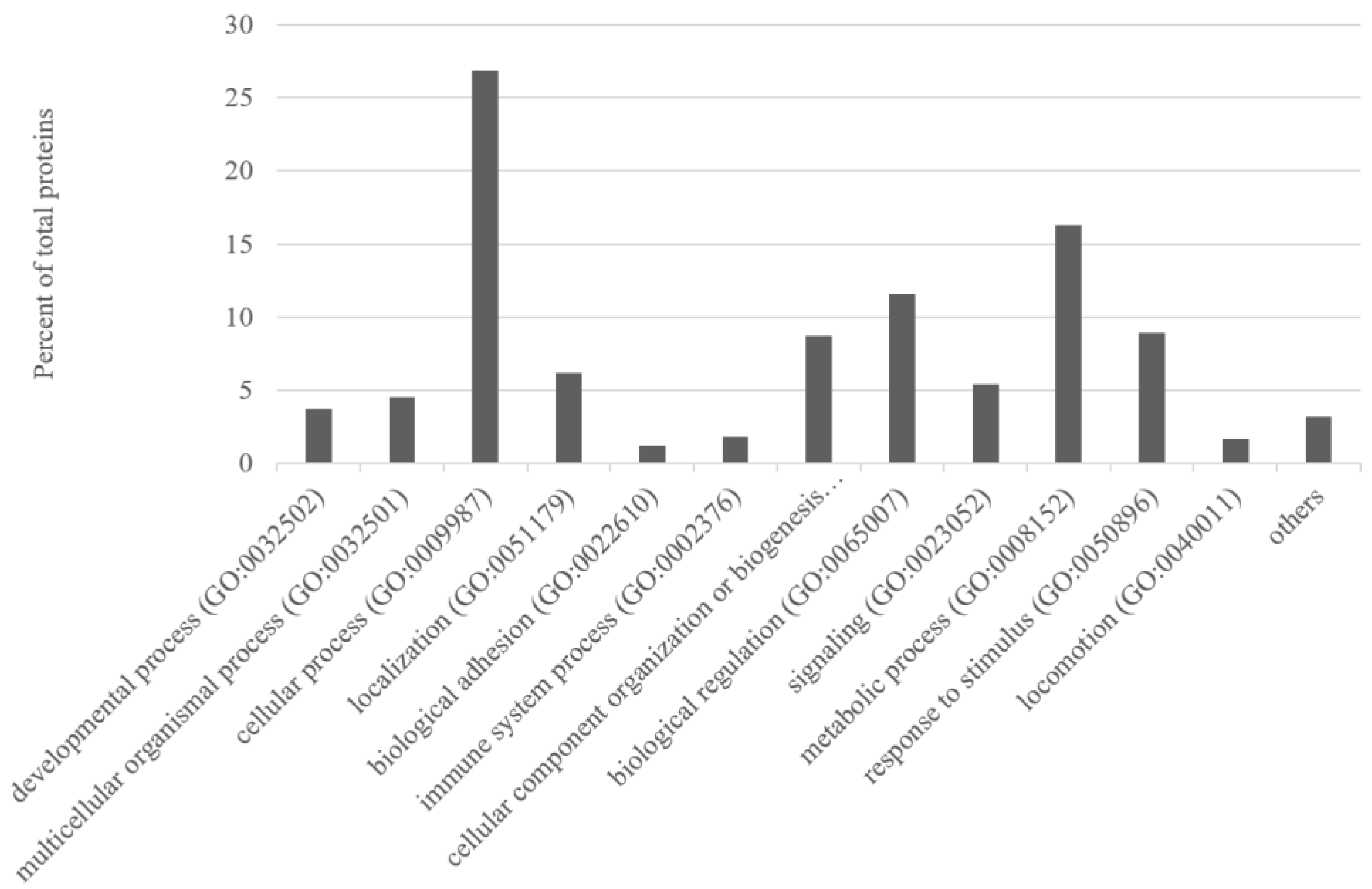
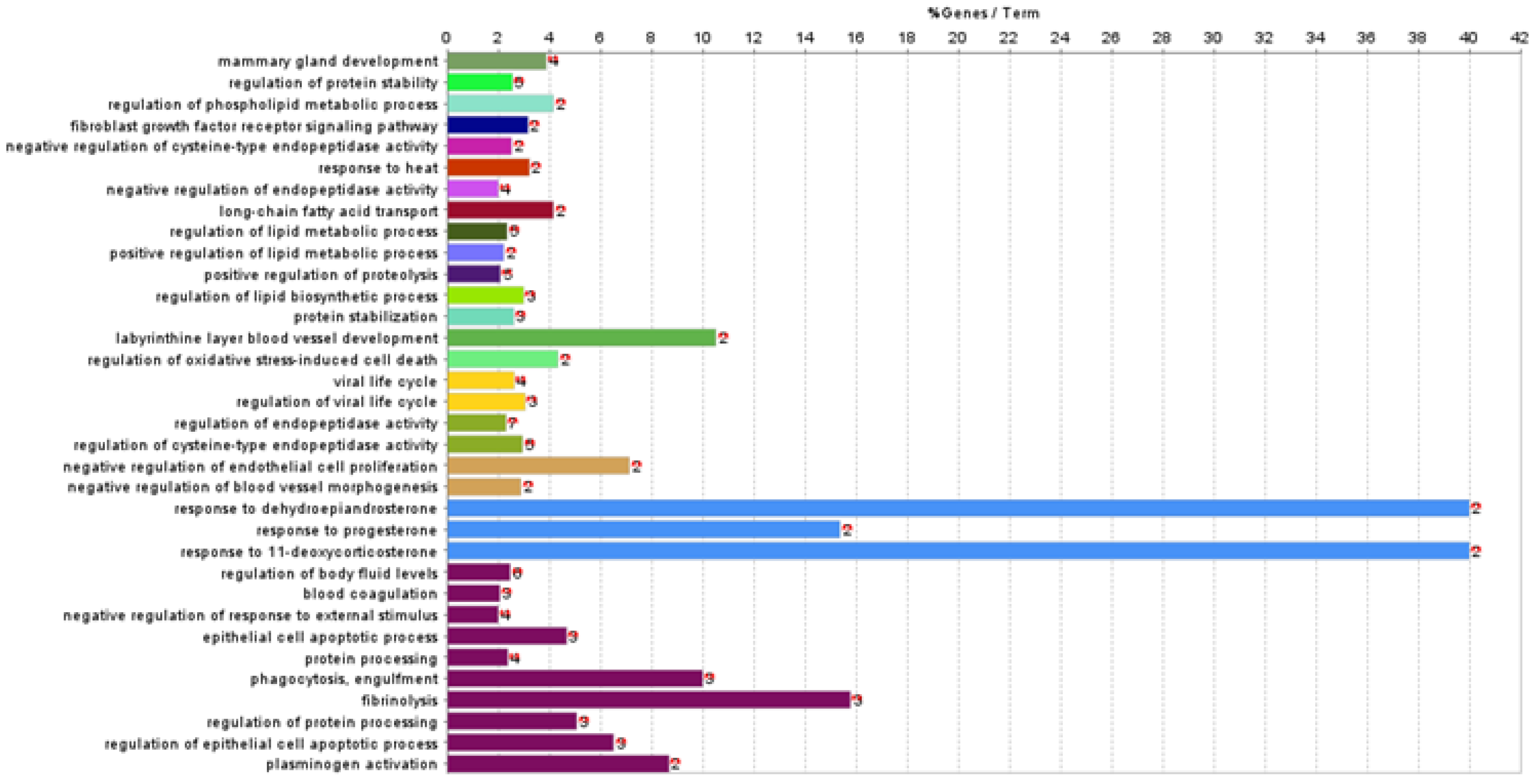
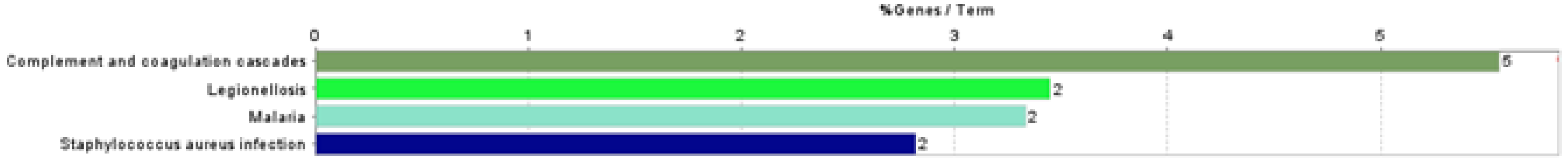
3.3. Functional Association Analysis
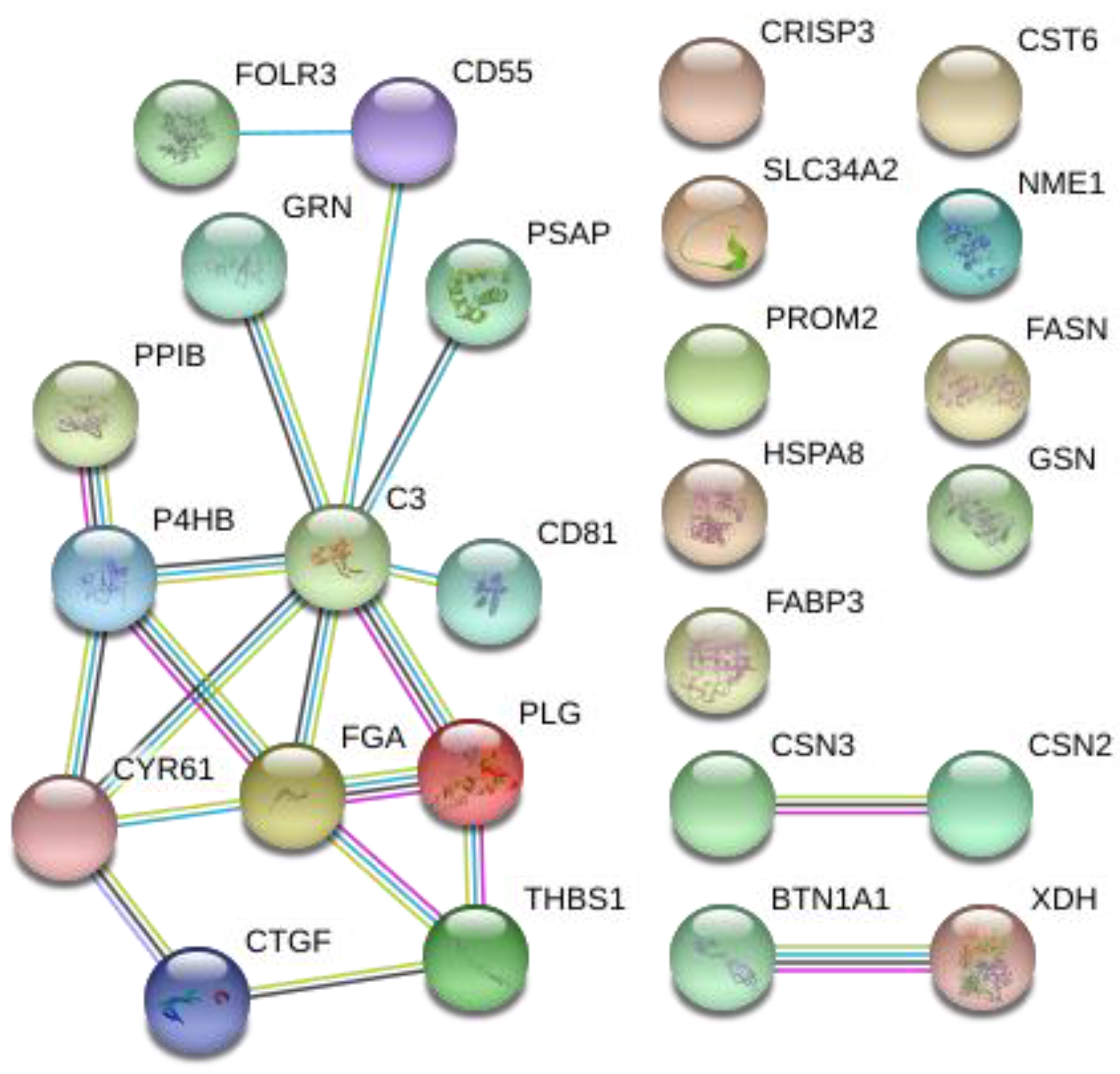
3.4. Protein–Protein Interaction (PPI) Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roncada, P.; Piras, C.; Soggiu, A.; Turk, R.; Urbani, A.; Bonizzi, L. Farm animal milk proteomics. J. Proteom. 2012, 75, 4259–4274. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Li, X.Y.; Zhao, X.; Qin, Y.S.; Zhang, X.X.; Li, J.; Wang, J.M.; Wang, C.F. Proteomics and microstructure profiling of goat milk protein after homogenization. J. Dairy Sci. 2019, 102, 3839–3850. [Google Scholar] [CrossRef] [PubMed]
- Vargas, M.; Cháfer, M.; Albors, A.; Chiralt, A.; González-Martínez, C. Physicochemical and sensory characteristics of yoghurt produced from mixtures of cows’ and goats’ milk. Int. Dairy J. 2008, 18, 1146–1152. [Google Scholar] [CrossRef]
- Ceballos, L.S.; Morales, E.R.; de la Torre Adarve, G.; Castro, J.D.; Martínez, L.P.; Sampelayo, M.R.S. Composition of goat and cow milk produced under similar conditions and analyzed by identical methodology. J. Food Compos. Anal. 2009, 22, 322–329. [Google Scholar] [CrossRef]
- Park, Y.W. Hypo-allergenic and therapeutic significance of goat milk. Small Rumin. Res. 1994, 14, 151–159. [Google Scholar] [CrossRef]
- Lara-Villoslada, F.; Olivares, M.; Jiménez, J.; Boza, J.; Xaus, J. Goat milk is less immunogenic than cow milk in a murine model of atopy. J. Pediatr. Gastroenterol. Nutr. 2004, 39, 354–360. [Google Scholar] [CrossRef]
- Haenlein, G.F.W. Past, present, and future perspectives of small ruminant dairy research. J. Dairy Sci. 2001, 84, 2097–2115. [Google Scholar] [CrossRef]
- Faye, B.; Konuspayeva, G. The sustainability challenge to the dairy sector—The growing importance of non-cattle milk production worldwide. Int. Dairy J. 2012, 24, 50–56. [Google Scholar] [CrossRef]
- Haenlein, G.F.W. Goat milk in human nutrition. In Small Ruminant Research; Elsevier: Amsterdam, The Netherlands, 2004; Volume 51, pp. 155–163. [Google Scholar]
- Di Luca, A.; Ianni, A.; Bennato, F.; Henry, M.; Meleady, P.; Martino, G. A Label-Free Quantitative Analysis for the Search of Proteomic Differences between Goat Breeds. Animals 2022, 12, 3336. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations Domestic Animal Diversity Information System (DAD-IS). Available online: http://www.fao.org/dad-is (accessed on 23 April 2020).
- Auerbach, D.; Thaminy, S.; Hottiger, M.O.; Stagljar, I. The post-genomic era of interactive proteomics: Facts and perspectives. Proteomics 2002, 2, 611. [Google Scholar] [CrossRef]
- Lottspeich, F. Proteome analysis: A pathway to the functional analysis of proteins. Angew. Chemie Int. Ed. 1999, 38, 2476. [Google Scholar] [CrossRef]
- Cebo, C.; Caillat, H.; Bouvier, F.; Martin, P. Major proteins of the goat milk fat globule membrane. J. Dairy Sci. 2010, 93, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Roncada, P.; Gaviraghi, A.; Liberatori, S.; Canas, B.; Bini, L.; Greppi, G.F. Identification of caseins in goat milk. Proteomics 2002, 2, 723–726. [Google Scholar] [CrossRef] [PubMed]
- Di Gerlando, R.; Tolone, M.; Sutera, A.M.; Monteleone, G.; Portolano, B.; Sardina, M.T.; Mastrangelo, S. Variation of proteomic profile during lactation in Girgentana goat milk: A preliminary study. Ital. J. Anim. Sci. 2019, 18, 88–97. [Google Scholar] [CrossRef]
- Neilson, K.A.; Ali, N.A.; Muralidharan, S.; Mirzaei, M.; Mariani, M.; Assadourian, G.; Lee, A.; van Sluyter, S.C.; Haynes, P.A. Less label, more free: Approaches in label-free quantitative mass spectrometry. Proteomics 2011, 11, 535. [Google Scholar] [CrossRef]
- Becker, C.H.; Bern, M. Recent developments in quantitative proteomics. Mutat. Res. 2011, 722, 171. [Google Scholar] [CrossRef]
- Lu, J.; Wang, X.; Zhang, W.; Liu, L.; Pang, X.; Zhang, S.; Lv, J. Comparative proteomics of milk fat globule membrane in different species reveals variations in lactation and nutrition. Food Chem. 2016, 196, 665–672. [Google Scholar] [CrossRef]
- Ginger, M.R.; Grigor, M.R. Comparative aspects of milk caseins. Comp. Biochem. Physiol.—B Biochem. Mol. Biol. 1999, 124, 133–145. [Google Scholar] [CrossRef]
- Fox, P.F.; Kelly, A.L. Developments in the chemistry and technology of milk proteins. 2. Minor milk proteins. Food Australia 2003, 55, 104–108. [Google Scholar]
- Affolter, M.; Grass, L.; Vanrobaeys, F.; Casado, B.; Kussmann, M. Qualitative and quantitative profiling of the bovine milk fat globule membrane proteome. J. Proteom. 2010, 73, 1079–1088. [Google Scholar] [CrossRef]
- Hogarth, C.J.; Fitzpatrick, J.L.; Nolan, A.M.; Young, F.J.; Pitt, A.; Eckersall, P.D. Differential protein composition of bovine whey: A comparison of whey from healthy animals and from those with clinical mastitis. Proteomics 2004, 4, 2094–2100. [Google Scholar] [CrossRef] [PubMed]
- da Costa, W.K.A.; de Souza, E.L.; Beltrão-Filho, E.M.; Vasconcelos, G.K.V.; Santi-Gadelha, T.; de Almeida Gadelha, C.A.; Franco, O.L.; Magnani, M. Comparative Protein Composition Analysis of Goat Milk Produced by the Alpine and Saanen Breeds in Northeastern Brazil and Related Antibacterial Activities. PLoS ONE 2014, 9, e93361. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhang, L.; Wu, Y.; Zhou, P. Changes in milk fat globule membrane proteome after pasteurization in human, bovine and caprine species. Food Chem. 2019, 279, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, C.; Sun, X.; Guo, M. Proteomic analysis of whey proteins in the colostrum and mature milk of Xinong Saanen goats. J. Dairy Sci. 2020, 103, 1164–1174. [Google Scholar] [CrossRef]
- Anagnostopoulos, A.K.; Katsafadou, A.I.; Pierros, V.; Kontopodis, E.; Fthenakis, G.C.; Arsenos, G.; Karkabounas, S.C.; Tzora, A.; Skoufos, I.; Tsangaris, G.T. Milk of Greek sheep and goat breeds; characterization by means of proteomics. J. Proteom. 2016, 147, 76–84. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Wisniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Coleman, O.; Henry, M.; Clynes, M.; Meleady, P. Filter-Aided Sample Preparation (FASP) for Improved Proteome Analysis of Recombinant Chinese Hamster Ovary Cells. Methods Mol. Biol. 2017, 1603, 187–194. [Google Scholar]
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER version 14: More genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2018, 47, D419–D426. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed]
- Bovo, S.; Di Luca, A.; Galimberti, G.; Dall’Olio, S.; Fontanesi, L. A comparative analysis of label-free liquid chromatography-mass spectrometry liver proteomic profiles highlights metabolic differences between pig breeds. PLoS ONE 2018, 13, e0199649. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Liu, L.; Pang, X.; Zhang, S.; Jia, Z.; Ma, C.; Zhao, L.; Lv, J. Comparative proteomics of milk fat globule membrane in goat colostrum and mature milk. Food Chem. 2016, 209, 10–16. [Google Scholar] [CrossRef]
- Chen, D.; Li, X.; Zhao, X.; Qin, Y.; Wang, J.; Wang, C. Comparative proteomics of goat milk during heated processing. Food Chem. 2019, 275, 504–514. [Google Scholar] [CrossRef]
- Cunsolo, V.; Fasoli, E.; Saletti, R.; Muccilli, V.; Gallina, S.; Righetti, P.G.; Foti, S. Zeus, Aesculapius, Amalthea and the proteome of goat milk. J. Proteom. 2015, 128, 69–82. [Google Scholar] [CrossRef]
- Zhao, L.; Du, M.; Gao, J.; Zhan, B.; Mao, X. Label-free quantitative proteomic analysis of milk fat globule membrane proteins of yak and cow and identification of proteins associated with glucose and lipid metabolism. Food Chem. 2019, 275, 59–68. [Google Scholar] [CrossRef]
- Keragala, C.B.; Draxler, D.F.; McQuilten, Z.K.; Medcalf, R.L. Haemostasis and innate immunity—A complementary relationship. Br. J. Haematol. 2018, 180, 782–798. [Google Scholar] [CrossRef]
- Moreno-Indias, I.; Dodds, A.W.; Argüello, A.; Castro, N.; Sim, R.B. The complement system of the goat: Haemolytic assays and isolation of major proteins. BMC Vet. Res. 2012, 8, 91. [Google Scholar] [CrossRef]
- Moreno-Indias, I.; Morales-delaNuez, A.; Hernández-Castellano, L.E.; Sánchez-Macías, D.; Capote, J.; Castro, N.; Argüello, A. Docosahexaenoic acid in the goat kid diet: Effects on immune system and meat quality1. J. Anim. Sci. 2012, 90, 3729–3738. [Google Scholar] [CrossRef]
- Janssen, B.J.C.; Huizinga, E.G.; Raaijmakers, H.C.A.; Roos, A.; Daha, M.R.; Nilsson-Ekdahl, K.; Nilsson, B.; Gros, P. Structures of complement component C3 provide insights into the function and evolution of immunity. Nature 2005, 437, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Rainard, P. The complement in milk and defense of the bovine mammary gland against infections. Vet. Res. 2003, 34, 647–670. [Google Scholar] [CrossRef] [PubMed]
- Tregoat, V.; Montagne, P.; Cuilliere, M.L.; Bene, M.C.; Faure, G.C. Sequential C3 and C4 levels in human milk in relation to prematurity and parity. Clin. Chem. Lab. Med. 2000, 38, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Prunkard, D.; Cottingham, I.; Garner, I.; Bruce, S.; Dalrymple, M. High-level expression of recombinant human fibrinogen in the milk of transgenic mice. Nat. Biotechnol. 1996, 14, 867–871. [Google Scholar] [CrossRef]
- Petersen, H.H.; Nielsen, J.P.; Heegaard, P.M.H. Application of acute phase protein measurements in veterinary clinical chemistry. Vet. Res. 2004, 35, 163–187. [Google Scholar] [CrossRef]
- Vet, T.J.; Sci, A.; Fasulkov, I.; Karadaev, M.; Vasilev, N.; Urumova, V.; Mircheva, T. Determination of plasma fibrinogen and haptoglobin, hematological and blood biochemical changes in Bulgarian local goats with experimentally induced Staphylococcus aureus mastitis. Turkish J. Vet. Anim. Sci. 2014, 38, 439–444. [Google Scholar]
- Nagy, O.; Tóthová, C.; Nagyová, V.; Kováč, G.; Pošivák, J. Changes in the serum protein electrophoretic pattern in lambs during the first month of life. Acta Vet. Brno 2014, 83, 187–193. [Google Scholar] [CrossRef]
- Yang, G.P.; Lau, L.F. Cell Growth & Differentiation Cyr6l, Product of a Growth Factor-inducible Immediate Early Gene, Is Associated with the Extracellular Matrix and the Cell Surface. Cell Growth Differ. 1991, 2, 351–357. [Google Scholar]
- Sherbet, G.V. Connective Tissue Growth Factor. In Growth Factors and Their Receptors in Cell Differentiation, Cancer and Cancer Therapy; Elsevier: Amsterdam, The Netherlands, 2011; pp. 105–110. [Google Scholar]
- Stelwagen, K.; Knight, C.H. Effect of unilateral once or twice daily milking of cows on milk yield and udder characteristics in early and late lactation. J. Dairy Res. 1997, 64, 487–494. [Google Scholar] [CrossRef]
- Littlejohn, M.D.; Walker, C.G.; Ward, H.E.; Lehnert, K.B.; Snell, R.G.; Verkerk, G.A.; Spelman, R.J.; Clark, D.A.; Davis, S.R. Effects of reduced frequency of milk removal on gene expression in the bovine mammary gland. Physiol. Genom. 2010, 41, 21–32. [Google Scholar] [CrossRef]
- Ozen, B.F.; Hayes, K.D.; Mauer, L.J. Measurement of plasminogen concentration and differentiation of plasmin and plasminogen using Fourier-transform infrared spectroscopy. Int. Dairy J. 2003, 13, 441–446. [Google Scholar] [CrossRef]
- Ismail, B.; Nielsen, S.S. Invited review: Plasmin protease in milk: Current knowledge and relevance to dairy industry. J. Dairy Sci. 2010, 93, 4999–5009. [Google Scholar] [CrossRef]
- Theodorou, G.; Kominakis, A.; Rogdakis, E.; Politis, I. Factors affecting the plasmin-plasminogen system in milk obtained from three Greek dairy sheep breeds with major differences in milk production capacity. J. Dairy Sci. 2007, 90, 3263–3269. [Google Scholar] [CrossRef] [PubMed]
- Bastian, E.D.; Brown, R.J. Plasmin in milk and dairy products: An update. Int. Dairy J. 1996, 6, 435–457. [Google Scholar] [CrossRef]
- Battacone, G.; Cannas, E.A.; Mazzette, A.; Dimauro, C.; Enne, G. Why does the increase of plasmin worsen the coagulation properties of milk in dairy sheep? Ital. J. Anim. Sci. 2016, 4, 342–344. [Google Scholar] [CrossRef]
- Ali Khan, H.; Mutus, B. Protein disulfide isomerase a multifunctional protein with multiple physiological roles. Front. Chem. 2014, 2, 70. [Google Scholar] [CrossRef]
- Galligan, J.J.; Petersen, D.R. The human protein disulfide isomerase gene family. Hum. Genom. 2012, 6, 6. [Google Scholar] [CrossRef]
- Pryce, J.E.; Royal, M.D.; Garnsworthy, P.C.; Mao, I.L. Fertility in the high-producing dairy cow. In Livestock Production Science; Elsevier: Amsterdam, The Netherlands, 2004; Volume 86, pp. 125–135. [Google Scholar]
- Royal, M.D.; Darwash, A.O.; Flint, A.P.F.; Webb, R.; Woolliams, J.A.; Lamming, G.E. Declining fertility in dairy cattle: Changes in traditional and endocrine parameters of fertility. Anim. Sci. 2000, 70, 487–501. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniProt * | Gene Name | Description | Peptides § | Score ¥ | ANOVA (p) | Fold Change | Highest Condition ǂ |
|---|---|---|---|---|---|---|---|
| A0A452DX18 | C3 | Complement C3 | 27 | 94.92 | 0.0097 | 4.03 | Teramana |
| A0A452F1G2 | THBS1 | Thrombospondin 1 | 9 | 24.44 | 0.0007 | 24.62 | Teramana |
| A0A452DRX5 | PSAP | Prosaposin | 6 | 18.88 | 0.0031 | 2.63 | Teramana |
| A0A452FHJ7 | GSN | Gelsolin | 6 | 20.57 | 0.0056 | 1.52 | Teramana |
| A0A452E7A0 | PLG | Plasminogen | 6 | 22.23 | 0.0241 | 2.27 | Teramana |
| A0A452FK23 | CSN2 | Beta-casein | 4 | 10.73 | 0.0086 | 2.06 | Teramana |
| A0A452G9D9 | CSN3 | Kappa-casein | 4 | 36.55 | 0.0140 | 2.53 | Teramana |
| A0A452E2Y0 | CLU | Clusterin | 4 | 15.79 | 0.0292 | 4.47 | Teramana |
| D6PX62 | Crisp3 | Cysteine-rich secretory protein 3 | 3 | 9.86 | 0.0026 | 38.63 | Teramana |
| A0A452G7V5 | P4HB | Protein disulfide-isomerase | 3 | 9.50 | 0.0245 | 2.12 | Teramana |
| A0A452EZB5 | CD55 | Complement decay-accelerating factor | 3 | 7.99 | 0.0267 | 3.89 | Teramana |
| A0A452F9Y8 | PPIB | Peptidyl-prolyl cis-trans isomerase | 3 | 10.26 | 0.0341 | 1.98 | Teramana |
| A0A452GBG4 | CYR61 | Cellular communication network factor 1 | 2 | 7.98 | 0.0001 | 23.17 | Teramana |
| A0A452EKN2 | IGHG4 | Immunoglobulin heavy constant gamma 4 | 2 | 6.11 | 0.0006 | 9.56 | Teramana |
| A0A452DZD6 | rig-5 | Ig-like domain-containing protein | 2 | 8.56 | 0.0029 | 1.51 | Teramana |
| A0A452G5W1 | GRN | Granulin precursor | 2 | 6.71 | 0.0034 | 3.02 | Teramana |
| A0A452F4L3 | FGA | Fibrinogen alpha chain | 2 | 5.47 | 0.0194 | 2.46 | Teramana |
| A0A452G077 | CTGF | Cellular communication network factor 2 | 2 | 6.67 | 0.0402 | 6.69 | Teramana |
| A0A452EZW6 | XDH | FAD-binding PCMH-type domain-containing protein | 14 | 38.58 | 0.0089 | 1.96 | Saanen |
| A0A452EYF6 | HSPA8 | Heat shock cognate 71 kDa protein | 5 | 15.53 | 0.0011 | 1.72 | Saanen |
| A0A452ERT5 | SLC34A2 | Solute carrier family 34 member 2 | 4 | 9.43 | 0.0002 | 5.17 | Saanen |
| A0A452E4K0 | CST6 | Cystatin E/M | 4 | 15.21 | 0.0041 | 2.86 | Saanen |
| Q6S4N9 | FABP3 | Fatty acid binding protein 3 | 3 | 7.85 | 0.0002 | 1.75 | Saanen |
| A0A452FN55 | BTN1A1 | Butyrophilin subfamily 1 member A1 | 3 | 8.43 | 0.0058 | 1.76 | Saanen |
| A0A452F4U3 | FASN | Fatty acid synthase | 3 | 10.66 | 0.0104 | 4.12 | Saanen |
| A0A452G9E6 | FOLR3 | Folate_rec domain-containing protein | 3 | 8.19 | 0.0164 | 1.83 | Saanen |
| A0A452F3X9 | PROM2 | Prominin 2 | 2 | 5.84 | 0.0116 | 2.99 | Saanen |
| A0A452E0U0 | CD81 | Tetraspanin | 2 | 13.07 | 0.0211 | 2.28 | Saanen |
| A0A452F230 | NME1 | Nucleoside diphosphate kinase | 2 | 4.55 | 0.0320 | 1.52 | Saanen |
| GOID | Description | Functional Group 1 | p-Value 2 | % of Associated Proteins 3 | No. of Proteins | Upregulated or Downregulated Proteins 4 |
|---|---|---|---|---|---|---|
| GO:0030879 | mammary gland development | G0 | 0.000057 | 3.88 | 4 | CSN2 ↓, CSN3 ↓, FASN ↑, XDH ↑ |
| GO:0031647 | regulation of protein stability | G1 | 0.000053 | 2.56 | 5 | CD81 ↑, CLU ↓, CSN3 ↓, GSN ↓, PPIB ↓, |
| GO:1903725 | regulation of phospholipid metabolic process | G2 | 0.003120 | 4.17 | 2 | CD81 ↑, FABP3 ↑ |
| GO:0008543 | fibroblast growth factor receptor signalling pathway | G3 | 0.004663 | 3.17 | 2 | CTGF ↓, THBS1 ↓ |
| GO:2000117 | negative regulation of cysteine-type endopeptidase activity | G4 | 0.006824 | 2.53 | 2 | CSN2 ↓, THBS1 ↓ |
| GO:0009408 | response to heat | G5 | 0.004820 | 3.23 | 2 | CSN2 ↓, PLG ↓ |
| GO:0010951 | negative regulation of endopeptidase activity | G6 | 0.000418 | 2.00 | 4 | C3 ↓, CSN2 ↓, CST6 ↑, THBS1 ↓ |
| GO:0015909 | long-chain fatty acid transport | G7 | 0.003120 | 4.17 | 2 | FABP3 ↑, THBS1 ↓ |
| GO:0019216 | regulation of lipid metabolic process | G8 | 0.000060 | 2.36 | 5 | C3 ↓, CD81 ↑, CYR61 ↓, FABP3 ↑, PSAP ↓ |
| GO:0045834 | positive regulation of lipid metabolic process | G9 | 0.007858 | 2.22 | 2 | CD81 ↑, CYR61 ↓ |
| GO:0045862 | positive regulation of proteolysis | G10 | 0.000071 | 2.09 | 5 | C3 ↓, CLU ↓, CYR61 ↓, GSN ↓, XDH ↑ |
| GO:0046890 | regulation of lipid biosynthetic process | G11 | 0.000733 | 3.00 | 3 | C3 ↓, CYR61 ↓, FABP3 ↑ |
| GO:0050821 | protein stabilisation | G12 | 0.001011 | 2.61 | 3 | CLU ↓, CSN3 ↓, PPIB ↓ |
| GO:0060716 | labyrinthine layer blood vessel development | G13 | 0.000680 | 10.53 | 2 | CYR61 ↓, PLG ↓ |
| GO:1903201 | regulation of oxidative stress-induced cell death | G14 | 0.003088 | 4.35 | 2 | P4HB ↓, PSAP ↓ |
| GO:0019058 | viral life cycle | G15 | 0.000186 | 2.63 | 4 | CD81 ↑, GSN ↓, P4HB ↓, PPIB ↓ |
| GO:1903900 | regulation of viral life cycle | G15 | 0.000186 | 3.06 | 3 | GSN ↓, P4HB ↓, PPIB ↓ |
| GO:0052548 | regulation of endopeptidase activity | G16 | 0.000004 | 2.31 | 7 | C3 ↓, CSN2 ↓, CST6 ↑, CYR61 ↓, GSN ↓, THBS1 ↓, XDH ↑ |
| GO:2000116 | regulation of cysteine-type endopeptidase activity | G16 | 0.000004 | 2.96 | 5 | CSN2 ↓, CYR61 ↓, GSN ↓, THBS1 ↓, XDH ↑ |
| GO:0001937 | negative regulation of endothelial cell proliferation | G17 | 0.006928 | 7.14 | 2 | THBS1 ↓, XDH ↑ |
| GO:2000181 | negative regulation of blood vessel morphogenesis | G17 | 0.006928 | 2.90 | 2 | THBS1 ↓, XDH ↑ |
| GO:1903494 | response to dehydroepiandrosterone | G18 | 0.000390 | 40.00 | 2 | CSN2 ↓, CSN3 ↓ |
| GO:0032570 | response to progesterone | G18 | 0.000390 | 15.38 | 2 | CSN2 ↓, CSN3 ↓ |
| GO:1903496 | response to 11-deoxycorticosterone | G18 | 0.000390 | 40.00 | 2 | CSN2 ↓, CSN3 ↓ |
| GO:0050878 | regulation of body fluid levels | G19 | 0.000014 | 2.48 | 6 | CSN2 ↓, CSN3 ↓, FGA ↓, PLG ↓, THBS1 ↓, XDH ↑ |
| GO:0007596 | blood coagulation | G19 | 0.000014 | 2.07 | 3 | FGA ↓, PLG ↓, THBS1 ↓ |
| GO:0032102 | negative regulation of response to external stimulus | G19 | 0.000014 | 2.00 | 4 | CSN2 ↓, FGA ↓, PLG ↓, THBS1 ↓ |
| GO:1904019 | epithelial cell apoptotic process | G19 | 0.000014 | 4.69 | 3 | FGA ↓, GSN ↓, THBS1 ↓ |
| GO:0016485 | protein processing | G19 | 0.000014 | 2.38 | 4 | C3 ↓, FGA ↓, GSN ↓, THBS1 ↓ |
| GO:0006911 | phagocytosis, engulfment | G19 | 0.000014 | 10.00 | 3 | C3 ↓, GSN ↓, THBS1 ↓ |
| GO:0042730 | fibrinolysis | G19 | 0.000014 | 15.79 | 3 | FGA ↓, PLG ↓, THBS1 ↓ |
| GO:0070613 | regulation of protein processing | G19 | 0.000014 | 5.08 | 3 | C3 ↓, GSN ↓, THBS1 ↓ |
| GO:1904035 | regulation of epithelial cell apoptotic process | G19 | 0.000014 | 6.52 | 3 | FGA ↓, GSN ↓, THBS1 ↓ |
| GO:0031639 | plasminogen activation | G19 | 0.000014 | 8.70 | 2 | FGA ↓, THBS1 ↓ |
| GOID | Description | Functional Group 1 | p-Value 2 | % of Associated Proteins 3 | No. of Proteins | Upregulated or Downregulated Proteins 4 |
|---|---|---|---|---|---|---|
| KEGG:04610 | Complement and coagulation cascades | G0 | 0.000004 | 5.56 | 5 | C3 ↓, CD55 ↓, CLU ↓, FGA ↓, PLG ↓ |
| KEGG:05134 | Legionellosis | G1 | 0.013192 | 3.45 | 2 | C3 ↓, HSPA8 ↑ |
| KEGG:05144 | Malaria | G2 | 0.009393 | 3.33 | 2 | CD81 ↑, THBS1 ↓ |
| KEGG:05150 | Staphylococcus aureus infection | G3 | 0.009755 | 2.82 | 2 | C3 ↓, PLG ↓ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Luca, A.; Bennato, F.; Ianni, A.; Grotta, L.; Henry, M.; Meleady, P.; Martino, G. Comparative Label-Free Liquid Chromatography–Mass Spectrometry Milk Proteomic Profiles Highlight Putative Differences between the Autochthon Teramana and Saanen Goat Breeds. Animals 2023, 13, 2263. https://doi.org/10.3390/ani13142263
Di Luca A, Bennato F, Ianni A, Grotta L, Henry M, Meleady P, Martino G. Comparative Label-Free Liquid Chromatography–Mass Spectrometry Milk Proteomic Profiles Highlight Putative Differences between the Autochthon Teramana and Saanen Goat Breeds. Animals. 2023; 13(14):2263. https://doi.org/10.3390/ani13142263
Chicago/Turabian StyleDi Luca, Alessio, Francesca Bennato, Andrea Ianni, Lisa Grotta, Michael Henry, Paula Meleady, and Giuseppe Martino. 2023. "Comparative Label-Free Liquid Chromatography–Mass Spectrometry Milk Proteomic Profiles Highlight Putative Differences between the Autochthon Teramana and Saanen Goat Breeds" Animals 13, no. 14: 2263. https://doi.org/10.3390/ani13142263
APA StyleDi Luca, A., Bennato, F., Ianni, A., Grotta, L., Henry, M., Meleady, P., & Martino, G. (2023). Comparative Label-Free Liquid Chromatography–Mass Spectrometry Milk Proteomic Profiles Highlight Putative Differences between the Autochthon Teramana and Saanen Goat Breeds. Animals, 13(14), 2263. https://doi.org/10.3390/ani13142263