
Antibiotic Resistance of Enterococcus Species in Ornamental Animal Feed

 , ,
, ,  and
and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolates
2.2. Total Microbial Level
2.3. Antimicrobial Susceptibility Testing
2.4. Resistance and Virulence Genes
2.5. Statistical Analysis
3. Results
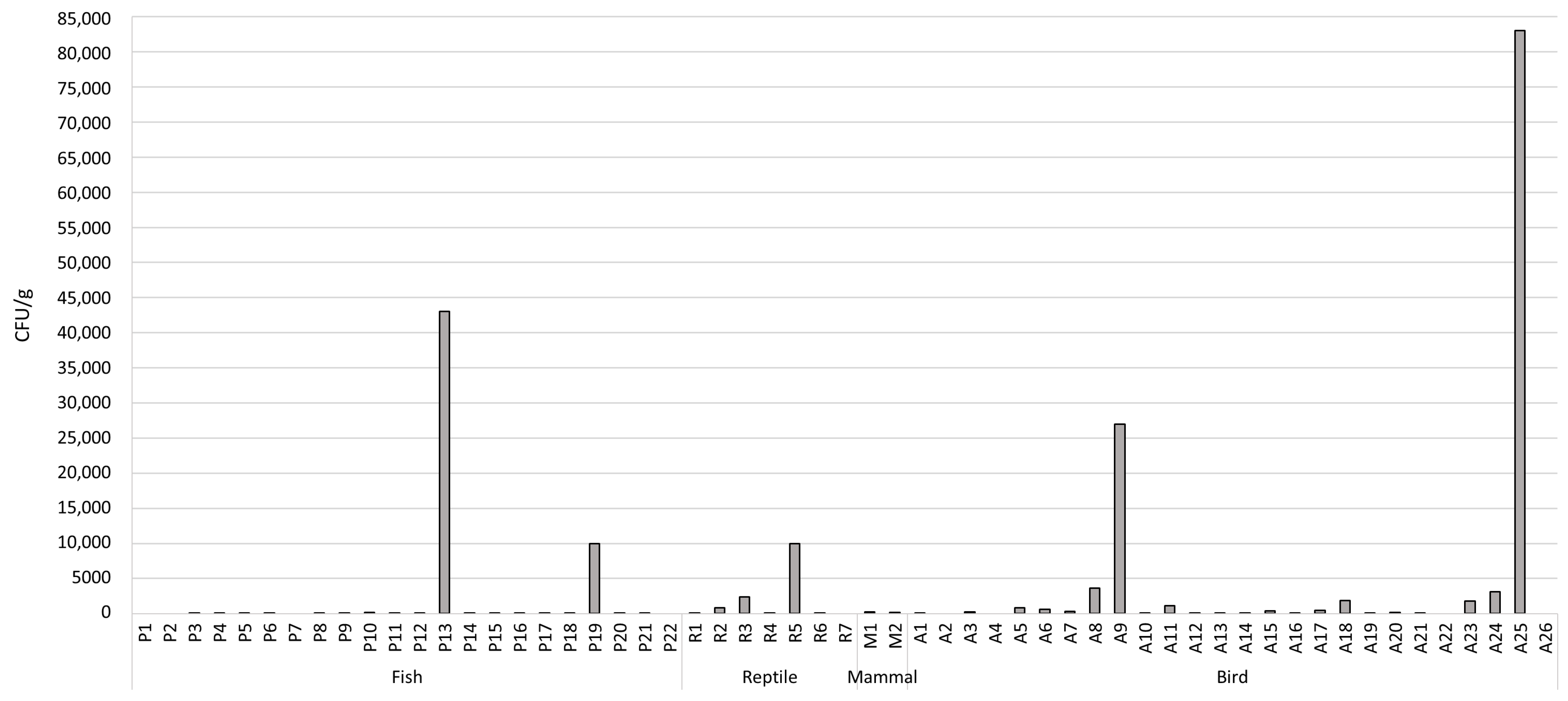
3.1. Total Microbial Level
3.2. Enterococcus Isolates/Species Identification
3.3. Antimicrobial Susceptibility Characterization
3.4. Genotypic of Antibiotic Resistance and Virulence Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guerrero-Ramos, E.; Cordero, J.; Molina-González, D.; Poeta, P.; Igrejas, G.; Alonso-Calleja, C.; Capita, R. Antimicrobial resistance and virulence genes in enterococci from wild game meat in Spain. Food Microbiol. 2016, 53, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Strateva, T.; Atanasova, D.; Savov, E.; Petrova, G.; Mitov, I. Incidence of virulence determinants in clinical Enterococcus faecalis and Enterococcus faecium isolates collected in Bulgaria. Braz. J. Infect. Diseas. 2016, 20, 127–133. [Google Scholar] [CrossRef] [PubMed]
- García-Solache, M.; Rice, L.B. The Enterococcus: A model of adaptability to its environment. Clin. Microbiol. Rev. 2019, 32, e00058-18. [Google Scholar] [CrossRef]
- Abamecha, A.; Wondafrash, B.; Abdissa, A. Antimicrobial resistance profile of Enterococcus species isolated from intestinal tracts of hospitalized patients in Jimma, Ethiopia Microbiology. BMC Res. Not. 2015, 8, 1–7. [Google Scholar]
- Martzy, R.; Kolm, C.; Brunner, K.; Mach, R.L.; Krska, R.; Sinkovec, H.; Sommer, R.; Farnleitner, A.H.; Reischer, G.H. A loop-mediated isothermal amplification (LAMP) assay for the rapid detection of Enterococcus spp. in water. Water Res. 2017, 122, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.C.; Souza, N.R.; Silva, P.H.C.; Santana, A.P. Resistência antimicrobiana em Enterococcus faecalis e Enterococcus faecium isolados de carcaças de frango. Pesq. Vet. Bras. 2013, 33, 575–580. [Google Scholar] [CrossRef]
- Thurlow, L.R.; Thomas, V.C.; Narayanan, S.; Olson, S.; Fleming, S.D.; Hancock, L.E. Gelatinase contributes to the pathogenesis of endocarditis caused by Enterococcus faecalis. Infect. Immun. 2010, 78, 4936–4943. [Google Scholar] [CrossRef]
- Zhang, Y.; Du, M.; Chang, Y.; Chen, L.; Zhang, Q. Incidence, clinical characteristics, and outcomes of nosocomial Enterococcus spp. bloodstream infections in a tertiary-care hospital in Beijing, China: A four-year retrospective study. Antimicrob. Resist. Infect. Control 2017, 6, 1–11. [Google Scholar] [CrossRef]
- Markwart, R.; Willrich, N.; Haller, S.; Noll, I.; Koppe, U.; Werner, G.; Eckmanns, T.; Reuss, A. The rise in vancomycin-resistant Enterococcus faecium in Germany: Data from the german antimicrobial resistance surveillance (ARS). Antimicrob. Resist. Infect. Control 2019, 8, 1–11. [Google Scholar] [CrossRef]
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and acquired resistance mechanisms in Enterococcus. Virul 2012, 3, 421–433. [Google Scholar] [CrossRef]
- Asmah, N. Molecular aspects of Enterococcus faecalis virulence. J. Syiah Kual Dentis. 2020, 5, 89–94. [Google Scholar] [CrossRef]
- Li, B.; Webster, T.J. Bacteria antibiotic resistance: New challenges and opportunities for implant-associated orthopedic infections. J. Orthop. Res. 2018, 36, 22–32. [Google Scholar] [CrossRef]
- Radhouani, H.; Silva, N.; Poeta, P.; Torres, C.; Correia, S.; Igrejas, G. Potential impact of antimicrobial resistance in wildlife, environment, and human health. Front. Microbiol. 2014, 5, 23. [Google Scholar] [CrossRef]
- Loureiro, R.J.; Roque, F.; Rodrigues, A.T.; Herdeiro, M.T.; Ramalheira, E. Use of antibiotics and bacterial resistances: Brief notes on its evolution. Rev. Port Saud Public 2016, 34, 77–84. [Google Scholar]
- Ventola, C.L.M.S. The antibiotic resistance crisis. Pharm Ther. 2015, 40, 277–283. [Google Scholar]
- Lee, J.H.; Shin, D.; Lee, B.; Lee, H.; Lee, I.; Jeong, D.W. Genetic Diversity and Antibiotic Resistance of Enterococcus faecalis Isolates from Traditional Korean Fermented Soybean Foods. J. Microbiol. Biotechnol. 2017, 27, 916–924. [Google Scholar] [CrossRef]
- Golob, M.; Pate, M.; Kušar, D.; Dermota, U.; Avberšek, J.; Papic, B.; Zdovc, I. Antimicrobial Resistance and Virulence Genes in Enterococcus faecium and Enterococcus faecalis from Humans and Retail Red Meat. BioMed. Res. Intern. 2019, 2019, 14–16. [Google Scholar] [CrossRef]
- Elghaieb, H.; Tedim, A.P.; Abbassi, M.S.; Novais, C.; Duarte, B.; Hassen, A.; Peixe, L.; Freitas, A.R. From farm to fork: Identical clones and Tn6674-like elements in linezolid-resistant Enterococcus faecalis from food-producing animals and retail meat. J. Antimicrob. Chemoth. 2020, 75, 30–35. [Google Scholar] [CrossRef]
- Ahmed, H.A.; Awad, N.F.S.; El-Hamid, M.I.A.; Shaker, A.; Mohamed, R.E.; Elsohaby, I. Pet birds as potential reservoirs of virulent and antibiotic resistant zoonotic bacteria. Comp. Immun. Microbiol. Infect. Diseas 2021, 75, 101606. [Google Scholar] [CrossRef]
- Carvalho, I.; Campo, R.D.; Sousa, M.; Silva, N.; Carrola, J.; Marinho, C.; Santos, T.; Carvalho, S.; Nóvoa, M.; Quaresma, M.; et al. Antimicrobial-resistant Escherichia coli and Enterococcus spp. isolated from Miranda donkey (Equus asinus): An old problem from a new source with a different approach. J. Med. Microbiol. 2017, 66, 191–202. [Google Scholar] [CrossRef]
- Dutka-Malen, S.; Evers, S.; Courvalin, P. Detection of Glycopeptide Resistance Genotypes and Identification to the Species Level of Clinically Relevant Enterococci by PCR. J. Clin. Microbiol. 1995, 33, 24–27. [Google Scholar] [CrossRef]
- Arias, C.A.; Robredo, B.; Singh, K.V.; Torres, C.; Panesso, D.; Murray, B.E. Rapid identification of Enterococcus hirae and Enterococcus durans by PCR and detection of a homologue of the E. hirae mur-2 Gene in E. durans. J. Clin. Microbiol. 2006, 44, 1567–1570. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Disk Susceptibility Tests, 13th ed.; CLSI standard M02; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Sutcliffe, J.; Grebe, T.; Tait-Kamradt, A.; Wondrack, L. Detection of erythromycin-resistant determinants by PCR. Antimicrob. Agents Chemother. 1996, 40, 2562–2566. [Google Scholar] [CrossRef]
- Aarestrup, F.M.; Agerso, Y.; Gerner-Smidt, P.; Madsen, M.; Jensen, L.B. Comparison of antimicrobial resistance phenotypes and resistance genes in Enterococcus faecalis and Enterococcus faecium from humans in the community, broilers, and pigs in Denmark. Diagn Microbiol. Infect. Dis. 2000, 37, 127–137. [Google Scholar] [CrossRef]
- Robredo, B.; Singh, K.V.; Baquero, F.; Murray, B.E.; Torres, C. Vancomycin-resistant Enterococci isolated from animals and food. Int. J. Food Microbiol. 2000, 54, 197–204. [Google Scholar] [CrossRef]
- Del Campo, R.; Tenorio, C.; Rubio, C.; Castillo, J.; Torres, C.; Gómez-Lus, R. Aminoglycoside-modifying enzymes in high-level streptomycin and gentamicin resistant Enterococcus spp. in Spain. Int. J. Antimicrob. Agents 2000, 15, 221–226. [Google Scholar] [CrossRef]
- Eaton, T.J.; Gasson, M.J. Molecular screening of Enterococcus virulence determinants and potential for genetic exchange between food and medical isolates. Appl. Environ. Microbiol. 2001, 67, 1628–1635. [Google Scholar] [CrossRef]
- Mannu, L.; Paba, A.; Daga, E.; Comunian, R.; Zanetti, S.; Duprè, I.; Sechi, L.A. Comparison of the incidence of virulence determinants and antibiotic resistance between Enterococcus faecium strains of dairy, animal and clinical origin. Int. J. Food Microbiol. 2003, 88, 291–304. [Google Scholar] [CrossRef]
- Miranda, C.; Silva, V.; Igrejas, G.; Poeta, P. Impact of European pet antibiotic use on enterococci and staphylococci antimicrobial resistance and human health. Future Microbiol. 2021, 16, 185–201. [Google Scholar] [CrossRef]
- Dolka, B.; Czopowicz, M.; Chrobak-Chmiel, D.; Ledwoń, A.; Szeleszczuk, P. Prevalence, antibiotic susceptibility and virulence factors of Enterococcus species in racing pigeons (Columba livia f. domestica). BMC Vet. Res. 2020, 16, 1–14. [Google Scholar] [CrossRef]
- Marinho, C. Resistência a antibióticos em Enterococcus spp. e Escherichia coli de equinodermes: Um problema ambiental e de saúde pública. Master’s Thesis, Dissertação de Mestrado em Genética Molecular Comparativa e Tecnológica, University of Trás-os-Montes and Alto Douro, Vila Real, Portugal, 2013; p. 109. Available online: http://hdl.handle.net/10348/5305 (accessed on 26 September 2022).
- Trościańczyk, A.; Nowakiewicz, A.; Gnat, S.; Łagowski, D.; Osińska, M. Are dogs and cats a reservoir of resistant and virulent Enterococcus faecalis strains and a potential threat to public health? J. Appl. Microbiol. 2021, 131, 2061–2071. [Google Scholar] [CrossRef]
- Osman, K.M.; Al-Maary, K.S.; Mubarak, A.S.; Dawoud, T.M.; Moussa, I.M.I.; Ibrahim, M.D.S.; Hessain, A.M.; Orabi, A.; Fawzy, N.M. Characterization and susceptibility of streptococci and enterococci isolated from Nile tilapia (Oreochromis niloticus) showing septicaemia in aquaculture and wild sites in Egypt. BMC Vet. Res. 2017, 13, 1–10. [Google Scholar] [CrossRef]
- Freitas, A.R.; Finisterra, L.; Tedim, A.P.; Duarte, B.; Novais, C.; Peixe, L. Linezolid- And multidrug-resistant enterococci in raw commercial dog food, Europe, 2019–2020. Emerg. Infect. Diseas 2021, 27, 2221–2224. [Google Scholar] [CrossRef]
- Santos, T. Proteogenómica de isolados de Enterococcus spp. e Escherichia coli com a utilização do MALDI-TOF MS. Master’s Thesis, Dissertação de Mestrado em Genética Molecular Comparativa e Tecnológica, University of Trás-os-Montes and Alto Douro, Vila Real, Portugal, 2014. Available online: http://hdl.handle.net/10348/5482 (accessed on 26 September 2022).
- Iweriebor, B.C.; Obi, L.C.; Okoh, A.I. Virulence and antimicrobial resistance factors of Enterococcus spp. isolated from fecal samples from piggery farms in Eastern Cape, South Africa. BMC Microbiol. 2015, 15, 136. [Google Scholar] [CrossRef]
- Andrade, S.D.E. Caracterização fenotípica e genotípica de amostras clínicas e indígenas de Enterococcus isoladas de seres humanos: Diversidade, virulência e resistência a drogas antimicrobianas. Tese de Doutoramento em Microbiologia. Ph.D. Thesis, Instituto de Ciências biológicas, Universidade Federal de Minas Gerais, Belo Horizonte, Brazil, 2018. Available online: http://hdl.handle.net/1843/BUOS-B76JMH (accessed on 26 September 2022).
- Van Tyne, D.; Martin, M.J.; Gilmore, M.S. Structure, function, and biology of the 85 Enterococcus faecalis cytolysin. Toxins 2013, 5, 895–911. [Google Scholar] [CrossRef]


{kind=link}
| Antibiotic Agent | Antimicrobial Class | Origin of Feed Sample | Total (%) | ||
|---|---|---|---|---|---|
| Bird (n = 58) | Fish (n = 33) | Reptile (n = 12) | |||
| Ampicilin | Penicilins | 3 | 0 | 0 | 3 (2.9) |
| Vancomycin | Glycopeptides | 1 | 2 | 0 | 3 (2.9) |
| Teicoplanin | Glycopeptides | 1 | 2 | 0 | 3 (2.9) |
| Tetracycline | Tetracyclines | 16 | 10 | 1 | 27 (26.2) |
| Erythromycin | Macrolides | 29 | 18 | 3 | 50 (48.5) |
| Ciprofloxacin | Fluoroquinolones | 24 | 8 | 7 | 39 (37.9) |
| Chloramphenicol | Phenicols | 3 | 1 | 0 | 4 (3.9) |
| Quinupristin/ Dalfopristin * | Streptogramins | 58 | 33 | 12 | 103 (100) |
| Nitrofurantoin | Nitrofurantoins | 11 | 5 | 3 | 19 (18.4) |
| Rifampicin | Ansamycins | 48 | 24 | 8 | 80 (77.7) |
| Fosfomycin | Fosfoycins | 5 | 2 | 0 | 7 (6.8) |
| Gentamicin | Aminoglicosides | 0 | 0 | 0 | 0 (0.0) |
| Streptomycin | Aminoglicosides | 0 | 0 | 0 | 0 (0.0) |
| Linezolid | Oxazolidinones | 15 | 3 | 2 | 20 (19.4) |
| Antibiotic Gene | E. faecalis (n = 22) | E. faecium (n = 19) | E. gallinarum (n = 15) | E. durans (n = 4) | Total (%) |
|---|---|---|---|---|---|
| ermA ermB ermC | 0 4 3 | 3 13 4 | 1 13 0 | 0 4 0 | 4 (6.7) 34 (56.7) 7 (11.7) |
| tetK tetM tetL | 5 8 4 | 5 0 13 | 7 10 10 | 2 4 4 | 19 (31,7) 22 (36.7) 31 (51.7) |
| aac(6′′)-aph(2′′) | 0 | 0 | 2 | 1 | 3 (5.0) |
| catA | 1 | 0 | 9 | 2 | 12 (20.0) |
| vatD vatE | 1 4 | 0 2 | 1 4 | 2 1 | 4 (6.7) 11 (18.3) |
| vanA vanB | 5 6 | 6 4 | 10 2 | 2 0 | 23 (38.3) 12 (20.0) |
| ant(6)-Ia | 3 | 4 | 10 | 4 | 21 (35.0) |
| Virulence Gene | E. faecalis (n = 22) | E. faecium (n = 19) | E. gallinarum (n = 15) | E. durans (n = 4) | Total (%) |
|---|---|---|---|---|---|
| esp | 10 | 0 | 12 | 2 | 24 (40.0) |
| ace | 0 | 0 | 0 | 0 | 0 (0.0) |
| gelE | 4 | 0 | 10 | 3 | 17 (28.3) |
| agg | 6 | 2 | 7 | 3 | 18 (30.0) |
| fsr | 0 | 0 | 0 | 0 | 0 (0.0) |
| cpd | 3 | 0 | 8 | 1 | 12 (20.0) |
| cylA | 1 | 0 | 4 | 2 | 7 (11.7) |
| cylB | 2 | 0 | 1 | 0 | 3 (5.0) |
| cylM | 8 | 0 | 3 | 3 | 14 (23.3) |
| cylL | 8 | 6 | 13 | 4 | 31 (51.7) |
| E. faecalis | E. faecium | E. gallinarum | χ2 | p | ||
|---|---|---|---|---|---|---|
| Antibiotic resistance genes | ||||||
| ermB | Positive Negative | 4 18 | 13 6 | 13 2 | 19.37 | 0.0001 |
| tetM | Positive Negative | 8 14 | 0 19 | 10 5 | 17.38 | 0.0002 |
| tetL | Positive Negative | 4 18 | 13 6 | 10 5 | 13.10 | 0.0014 |
| vanA | Positive Negative | 5 17 | 6 13 | 10 5 | 7.78 | 0.0205 |
| ant6-Ia | Positive Negative | 3 19 | 4 15 | 10 5 | 13.04 | 0.0015 |
| Virulence factors genes | ||||||
| cylL | Positive Negative | 8 14 | 6 13 | 13 2 | 12.27 | 0.0022 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soares, R.; Miranda, C.; Cunha, S.; Ferreira, L.; Martins, Â.; Igrejas, G.; Poeta, P. Antibiotic Resistance of Enterococcus Species in Ornamental Animal Feed. Animals 2023, 13, 1761. https://doi.org/10.3390/ani13111761
Soares R, Miranda C, Cunha S, Ferreira L, Martins Â, Igrejas G, Poeta P. Antibiotic Resistance of Enterococcus Species in Ornamental Animal Feed. Animals. 2023; 13(11):1761. https://doi.org/10.3390/ani13111761
Chicago/Turabian StyleSoares, Rúben, Carla Miranda, Sandra Cunha, Luís Ferreira, Ângela Martins, Gilberto Igrejas, and Patrícia Poeta. 2023. "Antibiotic Resistance of Enterococcus Species in Ornamental Animal Feed" Animals 13, no. 11: 1761. https://doi.org/10.3390/ani13111761
APA StyleSoares, R., Miranda, C., Cunha, S., Ferreira, L., Martins, Â., Igrejas, G., & Poeta, P. (2023). Antibiotic Resistance of Enterococcus Species in Ornamental Animal Feed. Animals, 13(11), 1761. https://doi.org/10.3390/ani13111761