
Loci Associated with Negative Heterosis for Viability and Meat Productivity in Interspecific Sheep Hybrids
 ,
,  , ,
, ,  ,
,
Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Genotype Data
2.3. Phenotype Data
2.4. Genome-Wide Association Study (GWAS)
2.4.1. Viability GWAS
2.4.2. GWAS of the Four Indices and Mass
2.5. Functional Annotation
2.6. Estimation of Heritability
2.7. Replication Analysis
3. Results
3.1. Viability GWAS
3.2. GWAS of Four Indices and Mass
3.3. Functional Annotation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jueken, A.; Hubdar, A.K.; Yiming, S.; Li, Q.F.; Xie, Z. Comparative Analysis on the Performance of the Hybrid Offspring of Wild Argali and Bashibay Sheep. Afr. J. Biotechnol. 2011, 10, 5539–5544. [Google Scholar]
- Li, X.; He, S.-G.; Li, W.-R.; Luo, L.-Y.; Yan, Z.; Mo, D.-X.; Wan, X.; Lv, F.-H.; Yang, J.; Xu, Y.-X.; et al. Genomic analyses of Pamir argali, Tibetan sheep, and their hybrids provide insights into chromosome evolution, phenotypic variation, and germplasm innovation. Genome Res. 2022, 32, 1669–1684. [Google Scholar] [CrossRef] [PubMed]
- Mavrogenis, A.P.; Herzogi, A. Interspecific Hybridization of the Cyprus Mouflon (Agrinon) with Domestic Sheep. Anim. Genet. Resour. Inf. 1990, 7, 23–27. [Google Scholar] [CrossRef]
- Subramaniam, R.; Shanthalingam, S.; Bavananthasivam, J.; Kugadas, A.; Raghavan, B.; Batra, S.A.; Herndon, C.N.; Rodriguez, J.; Tibary, A.; Nelson, D.; et al. Bighorn Sheep × Domestic Sheep Hybrids Survive Mannheimia Haemolytica Challenge in the Absence of Vaccination. Vet. Microbiol. 2014, 170, 278–283. [Google Scholar] [CrossRef]
- Ferreira, V.C.; Rosa, G.J.M.; Berger, Y.M.; Thomas, D.L. Survival in Crossbred Lambs: Breed and Heterosis Effects. J. Anim. Sci. 2015, 93, 912–919. [Google Scholar] [CrossRef]
- Lippman, Z.B.; Zamir, D. Heterosis: Revisiting the Magic. Trends Genet. 2007, 23, 60–66. [Google Scholar] [CrossRef]
- Jones, D.F. Dominance of Linked Factors as a Means of Accounting for Heterosis. Proc. Natl. Acad. Sci. USA 1917, 3, 310–312. [Google Scholar] [CrossRef]
- Li, L.; Lu, K.; Chen, Z.; Mu, T.; Hu, Z.; Li, X. Dominance, Overdominance and Epistasis Condition the Heterosis in Two Heterotic Rice Hybrids. Genetics 2008, 180, 1725–1742. [Google Scholar] [CrossRef]
- Zhou, G.; Chen, Y.; Yao, W.; Zhang, C.; Xie, W.; Hua, J.; Xing, Y.; Xiao, J.; Zhang, Q. Genetic Composition of Yield Heterosis in an Elite Rice Hybrid. Proc. Natl. Acad. Sci. USA 2012, 109, 15847–15852. [Google Scholar] [CrossRef]
- Yu, S.B.; Li, J.X.; Xu, C.G.; Tan, Y.F.; Gao, Y.J.; Li, X.H.; Zhang, Q.; Maroof, M.A.S. Importance of epistasis as the genetic basis of heterosis in an elite rice hybrid. Proc. Natl. Acad. Sci. USA 1997, 94, 9226–9231. [Google Scholar] [CrossRef]
- Schnell, F.W.; Cockerham, C.C. Multiplicative vs. Arbitrary Gene Action in Heterosis. Genetics 1992, 131, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Melchinger, A.E.; Utz, H.F.; Piepho, H.P.; Zeng, Z.B.; Schön, C.C. The Role of Epistasis in the Manifestation of Heterosis: A Systems-Oriented Approach. Genetics 2007, 177, 1815–1825. [Google Scholar] [CrossRef] [PubMed]
- Boeven, P.H.G.; Zhao, Y.; Thorwarth, P.; Liu, F.; Maurer, H.P.; Gils, M.; Schachschneider, R.; Schacht, J.; Ebmeyer, E.; Kazman, E.; et al. Negative Dominance and Dominance-by-Dominance Epistatic Effects Reduce Grain-Yield Heterosis in Wide Crosses in Wheat. Sci. Adv. 2020, 6, eaay4897. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.J.; Li, Z.K.; Mei, H.W.; Shu, Q.Y.; Tabien, R.; Zhong, D.B.; Ying, C.S.; Stansel, J.W.; Khush, G.S.; Paterson, A.H. Overdominant Epistatic Loci Are the Primary Genetic Basis of Inbreeding Depression and Heterosis in Rice. II. Grain Yield Components. Genetics 2001, 158, 1755–1771. [Google Scholar] [CrossRef]
- Li, Z.K.; Luo, L.J.; Mei, H.W.; Wang, D.L.; Shu, Q.Y.; Tabien, R.; Zhong, D.B.; Ying, C.S.; Stansel, J.W.; Khush, G.S.; et al. Overdominant Epistatic Loci Are the Primary Genetic Basis of Inbreeding Depression and Heterosis in Rice. I. Biomass and Grain Yield. Genetics 2001, 158, 1737–1753. [Google Scholar] [CrossRef]
- Malinovskaya, L.P.; Tishakova, K.V.; Bikchurina, T.I.; Slobodchikova, A.Y.; Torgunakov, N.Y.; Torgasheva, A.A.; Tsepilov, Y.A.; Volkova, N.A.; Borodin, P.M. Negative Heterosis for Meiotic Recombination rate in Spermatocytes of the Domestic Chicken Gallus Gallus. Vavilov J. Genet. Breed. 2021, 25, 661–668. [Google Scholar] [CrossRef]
- Mai, C.; Wen, C.; Xu, Z.; Xu, G.; Chen, S.; Zheng, J.; Sun, C.; Yang, N. Genetic Basis of Negative Heterosis for Growth Traits in Chickens Revealed by Genome-Wide Gene Expression Pattern Analysis. J. Anim. Sci. Biotechnol. 2021, 12, 52. [Google Scholar] [CrossRef]
- Arthur, P.F.; Makarechian, M.; Price, M.A.; Berg, R.T. Heterosis, Maternal and Direct Effects in Double-Muscled and Normal Cattle: I. Reproduction and Growth Traits. J. Anim. Sci. 1989, 67, 902–910. [Google Scholar] [CrossRef]
- Clasen, J.B.; Norberg, E.; Madsen, P.; Pedersen, J.; Kargo, M. Estimation of Genetic Parameters and Heterosis for Longevity in Crossbred Danish Dairy Cattle. J. Dairy Sci. 2017, 100, 6337–6342. [Google Scholar] [CrossRef]
- Iolchiev, B.S.; Volkova, N.A.; Bagirov, V.A.; Zinovieva, N.A. Identification of Interspecific Hybrids Argali (Ovis Ammon) and Domestic Sheep (Ovis Aries) of Different Generations by Exterior Indicators. Sel’skokhozyaistvennaya Biol. 2020, 55, 1139–1147. [Google Scholar] [CrossRef]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.A.M.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-Generation PLINK: Rising to the Challenge of Larger and Richer Datasets. Gigascience 2015, 4, 7. [Google Scholar] [CrossRef]
- Aulchenko, Y.S.; Ripke, S.; Isaacs, A.; van Duijn, C.M. GenABEL: An R Library for Genome-Wide Association Analysis. Bioinformatics 2007, 23, 1294–1296. [Google Scholar] [CrossRef] [PubMed]
- Alderson, G.L.H. The Development of a System of Linear Measurements to Provide an Assessment of Type and Function of Beef Cattle. Anim. Genet. Resour. Inf. 1999, 25, 45–55. [Google Scholar] [CrossRef]
- Salako, A.E. Application of Morphological Indices in the Assessment of Type and Function in Sheep. Int. J. Morphol. 2006, 24, 13–18. [Google Scholar] [CrossRef]
- Dahl, A.; Iotchkova, V.; Baud, A.; Johansson, A.; Gyllensten, U.; Soranzo, N.; Mott, R.; Kranis, A.; Marchini, J. A Multiple-Phenotype Imputation Method for Genetic Studies. Nat. Genet. 2016, 48, 466–472. [Google Scholar] [CrossRef]
- Svishcheva, G.R.; Axenovich, T.I.; Belonogova, N.M.; van Duijn, C.M.; Aulchenko, Y.S. Rapid Variance Components—Based Method for Whole-Genome Association Analysis. Nat. Genet. 2012, 44, 1166–1170. [Google Scholar] [CrossRef]
- Zlobin, A.S.; Volkova, N.A.; Borodin, P.M.; Aksenovich, T.I.; Tsepilov, Y.A. Recent Advances in Understanding Genetic Variants Associated with Growth, Carcass and Meat Productivity Traits in Sheep (Ovis Aries): An Update. Arch. Anim. Breed. 2019, 62, 579–583. [Google Scholar] [CrossRef]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef]
- Yang, J.; Lee, S.H.; Goddard, M.E.; Visscher, P.M. GCTA: A Tool for Genome-Wide Complex Trait Analysis. Am. J. Hum. Genet. 2011, 88, 76–82. [Google Scholar] [CrossRef]
- Zlobin, A.S.; Nikulin, P.S.; Volkova, N.A.; Zinovieva, N.A.; Iolchiev, B.S.; Bagirov, V.A.; Borodin, P.M.; Aksenovich, T.I.; Tsepilov, Y.A. Multivariate Analysis Identifies Eight Novel Loci Associated with Meat Productivity Traits in Sheep. Genes 2021, 12, 367. [Google Scholar] [CrossRef]
- Chen, P.W.; Jian, X.; Yoon, H.Y.; Randazzo, P.A. ARAP2 Signals through Arf6 and Rac1 to Control Focal Adhesion Morphology. J. Biol. Chem. 2013, 288, 5849–5860. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.W.; Luo, R.; Jian, X.; Randazzo, P.A. The Arf6 GTPase-Activating Proteins ARAP2 and ACAP1 Define Distinct Endosomal Compartments That Regulate Integrin A5β1 Traffic. J. Biol. Chem. 2014, 289, 30237–30248. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, A.; Håversen, L.; Mobini, R.; Andersson, L.; Ståhlman, M.; Lu, E.; Rutberg, M.; Fogelstrand, P.; Ekroos, K.; Mardinoglu, A.; et al. ARAP2 Promotes GLUT1-Mediated Basal Glucose Uptake through Regulation of Sphingolipid Metabolism. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2016, 1861, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- De Souza, T.C.; da Cruz, V.A.R.; Mourão, G.B.; Pedrosa, V.B.; Rovadoscki, G.A.; Coutinho, L.L.; de Camargo, G.M.F.; Costa, R.B.; de Carvalho, G.G.P.; Pinto, L.F.B. Estimates of Heritability and Candidate Genes for Primal Cuts and Dressing Percentage in Santa Ines Sheep. Livest. Sci. 2022, 264, 105048. [Google Scholar] [CrossRef]
- Lu, X.; Abdalla, I.M.; Nazar, M.; Fan, Y.; Zhang, Z.; Wu, X.; Xu, T.; Yang, Z. Genome-Wide Association Study on Reproduction-Related Body-Shape Traits of Chinese Holstein Cows. Animals 2021, 11, 1927. [Google Scholar] [CrossRef]
- Lee, S.H.; van der Werf, J.; Lee, S.H.; Lim, D.J.; Park, E.W.; Gondro, C.; Yoon, D.; Oh, S.J.; Kim, O.H.; Gibson, J.; et al. Genome wide QTL mapping to identify candidate genes for carcass traits in Hanwoo (Korean Cattle). Genes Genom. 2012, 34, 43–49. [Google Scholar] [CrossRef]
- Salpietro, V.; Perez-Dueñas, B.; Nakashima, K.; San Antonio-Arce, V.; Manole, A.; Efthymiou, S.; Vandrovcova, J.; Bettencourt, C.; Mencacci, N.E.; Klein, C.; et al. A Homozygous Loss-of-function Mutation in PDE2A Associated to Early-onset Hereditary Chorea. Mov. Disord. 2018, 33, 482–488. [Google Scholar] [CrossRef]
- Doummar, D.; Dentel, C.; Lyautey, R.; Metreau, J.; Keren, B.; Drouot, N.; Malherbe, L.; Bouilleret, V.; Courraud, J.; Valenti-Hirsch, M.P.; et al. Biallelic PDE2A Variants: A New Cause of Syndromic Paroxysmal Dyskinesia. Eur. J. Hum. Genet. 2020, 28, 1403–1413. [Google Scholar] [CrossRef]
- Miura, K.; Jacques, K.M.; Stauffer, S.; Kubosaki, A.; Zhu, K.; Hirsch, D.S.; Resau, J.; Zheng, Y.; Randazzo, P.A. ARAP1: A Point of Convergence for Arf and Rho Signaling. Mol. Cell 2002, 9, 109–119. [Google Scholar] [CrossRef]
- Kulzer, J.R.; Stitzel, M.L.; Morken, M.A.; Huyghe, J.R.; Fuchsberger, C.; Kuusisto, J.; Laakso, M.; Boehnke, M.; Collins, F.S.; Mohlke, K.L. A Common Functional Regulatory Variant at a Type 2 Diabetes Locus Upregulates ARAP1 Expression in the Pancreatic Beta Cell. Am. J. Hum. Genet. 2014, 94, 186–197. [Google Scholar] [CrossRef]
- Ahmed, Z.M.; Riazuddin, S.; Bernstein, S.L.; Ahmed, Z.; Khan, S.; Griffith, A.J.; Morell, R.J.; Friedman, T.B.; Riazuddin, S.; Wilcox, E.R. Mutations of the Protocadherin Gene PCDH15 Cause Usher Syndrome Type 1F. Am. J. Hum. Genet. 2001, 69, 25–34. [Google Scholar] [CrossRef]
- Brown, D.; Swan, A.A.; Gill, J.S.; Ball, A.J.; Banks, R.G. Genetic Parameters for Liveweight, Wool and Worm Resistance Traits in Multi-Breed Australian Meat Sheep. 1. Description of Traits, Fixed Effects, Variance Components and Their Ratios. Anim. Prod. Sci. 2015, 56, 1442–1448. [Google Scholar] [CrossRef]
- Vargas Jurado, N.; Notter, D.R.; Taylor, J.B.; Brown, D.J.; Mousel, M.R.; Lewis, R.M. Model Definition for Genetic Evaluation of Purebred and Crossbred Lambs Including Heterosis. J. Anim. Sci. 2022, 100, skac188. [Google Scholar] [CrossRef] [PubMed]
- Cloete, S.W.P.; Thutwa, K.; Scholtz, A.J.; Cloete, J.J.E.; Dzama, K.; Gilmour, A.R.; van Wyk, J.B. Breed Effects and Heterosis for Weight Traits and Tick Count in a Cross between an Indigenous Fat-Tailed and a Commercial Sheep Breed. Trop. Anim. Health Prod. 2021, 53, 165. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Days after Birth | Number of Alive Animals |
|---|---|
| 6 | 87 |
| 42 | 83 |
| 90 | 58 |
| 180 | 7 |
| SNP | CHR | POS | p | A1/A2 | AF | P.11 | P.12 | P.22 | AF Romanovskaya | AF Argali | Nearest Gene | Trait |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
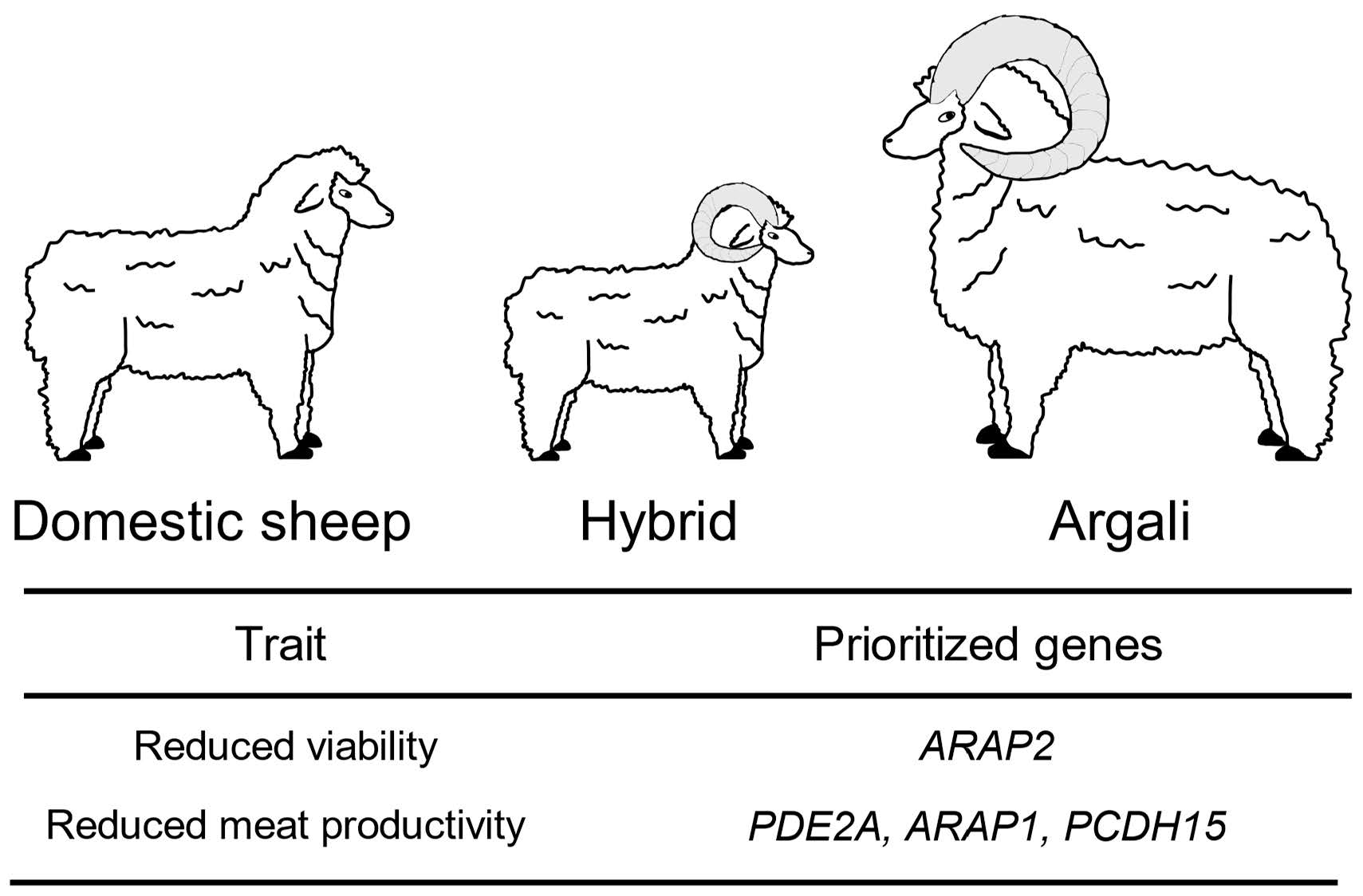
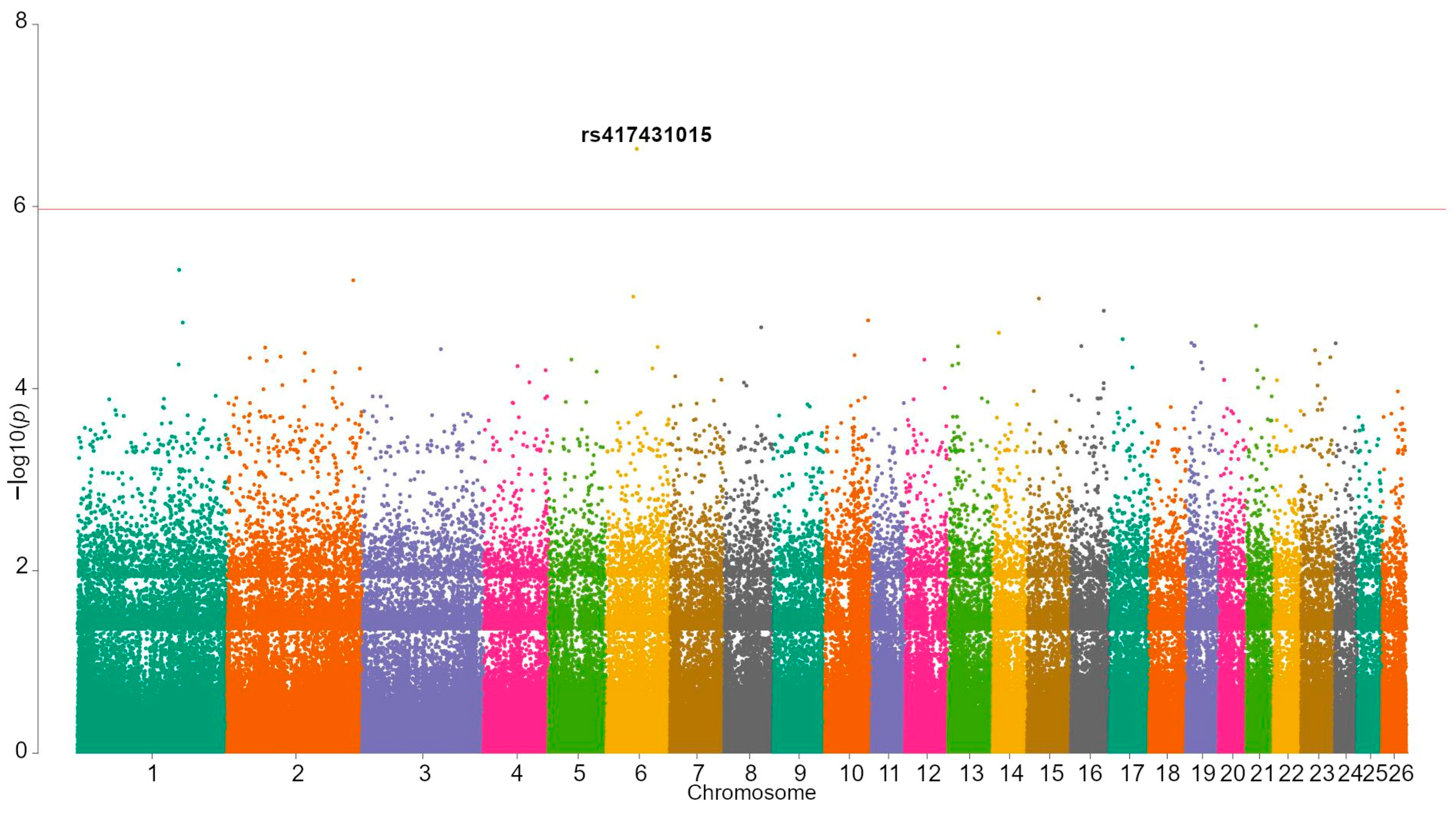
| rs417431015 | 6 | 55,587,508 | 2.33 × 10−7 | G/A | 0.368 | 28 | 54 | 5 | 0.5 | 0.6 | ARAP2 | Viability |
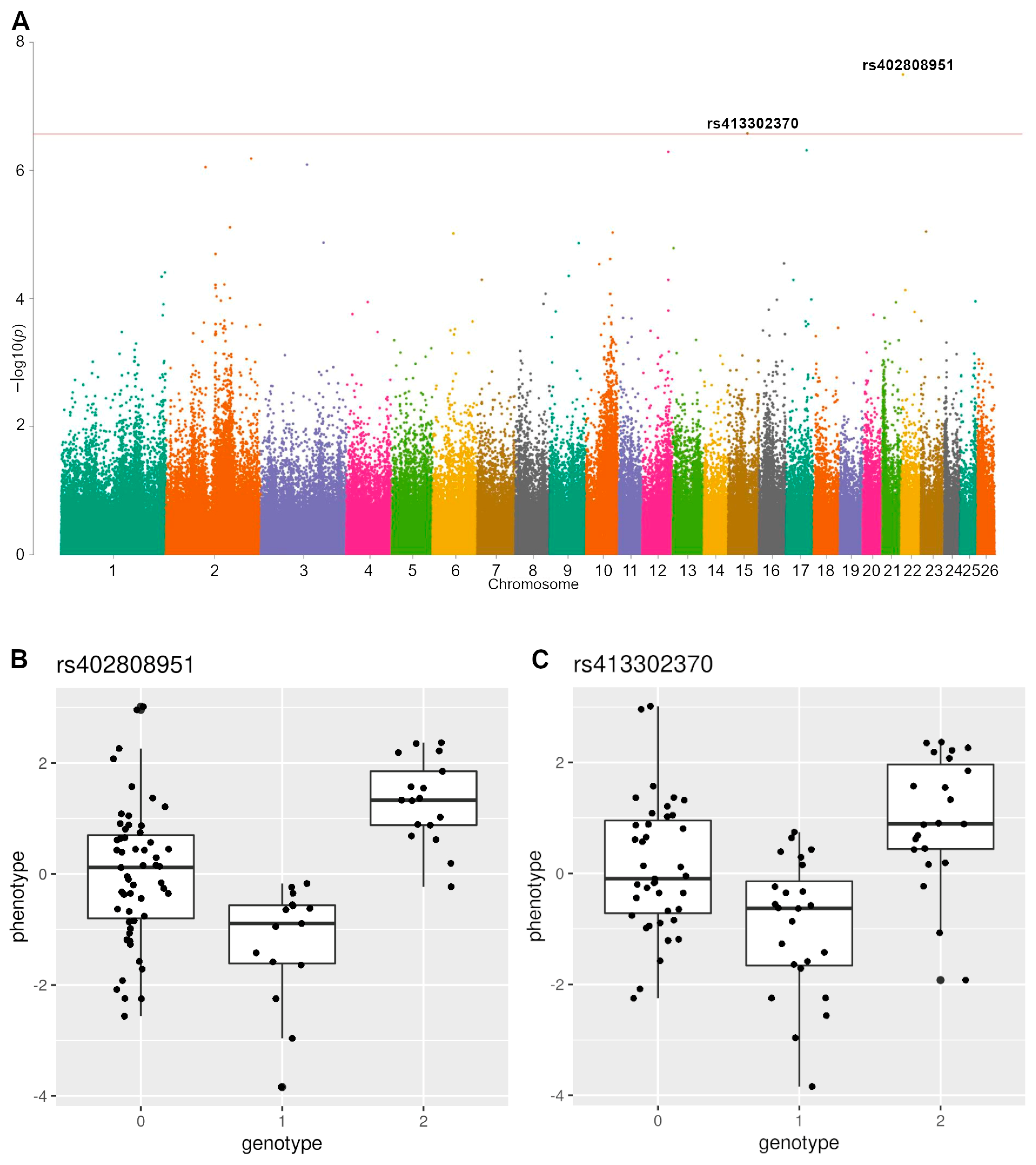
| rs413302370 | 15 | 50,414,913 | 2.65 × 10−7 | A/G | 0.407 | 39 | 24 | 23 | 0.667 | 1 | PDE2A ARAP1 | Compactness index |
| rs402808951 | 22 | 4,457,667 | 3.18 × 10−8 | A/G | 0.282 | 55 | 15 | 17 | 0.278 | 1 | PCDH15 | Compactness index |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zlobin, A.S.; Volkova, N.A.; Zinovieva, N.A.; Iolchiev, B.S.; Bagirov, V.A.; Borodin, P.M.; Axenovich, T.I.; Tsepilov, Y.A. Loci Associated with Negative Heterosis for Viability and Meat Productivity in Interspecific Sheep Hybrids. Animals 2023, 13, 184. https://doi.org/10.3390/ani13010184
Zlobin AS, Volkova NA, Zinovieva NA, Iolchiev BS, Bagirov VA, Borodin PM, Axenovich TI, Tsepilov YA. Loci Associated with Negative Heterosis for Viability and Meat Productivity in Interspecific Sheep Hybrids. Animals. 2023; 13(1):184. https://doi.org/10.3390/ani13010184
Chicago/Turabian StyleZlobin, Alexander S., Natalia A. Volkova, Natalia A. Zinovieva, Baylar S. Iolchiev, Vugar A. Bagirov, Pavel M. Borodin, Tatiana I. Axenovich, and Yakov A. Tsepilov. 2023. "Loci Associated with Negative Heterosis for Viability and Meat Productivity in Interspecific Sheep Hybrids" Animals 13, no. 1: 184. https://doi.org/10.3390/ani13010184
APA StyleZlobin, A. S., Volkova, N. A., Zinovieva, N. A., Iolchiev, B. S., Bagirov, V. A., Borodin, P. M., Axenovich, T. I., & Tsepilov, Y. A. (2023). Loci Associated with Negative Heterosis for Viability and Meat Productivity in Interspecific Sheep Hybrids. Animals, 13(1), 184. https://doi.org/10.3390/ani13010184