
Identification of Candidate Genes for Pigmentation in Camels Using Genotyping-by-Sequencing

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
- Y: coat color
- α: SNP effects
- U: kinship background effects
- e: residual effects
- X and Z: incidence matrix relating the individuals to fixed marker effects α and random group effects u, respectively.
3. Results
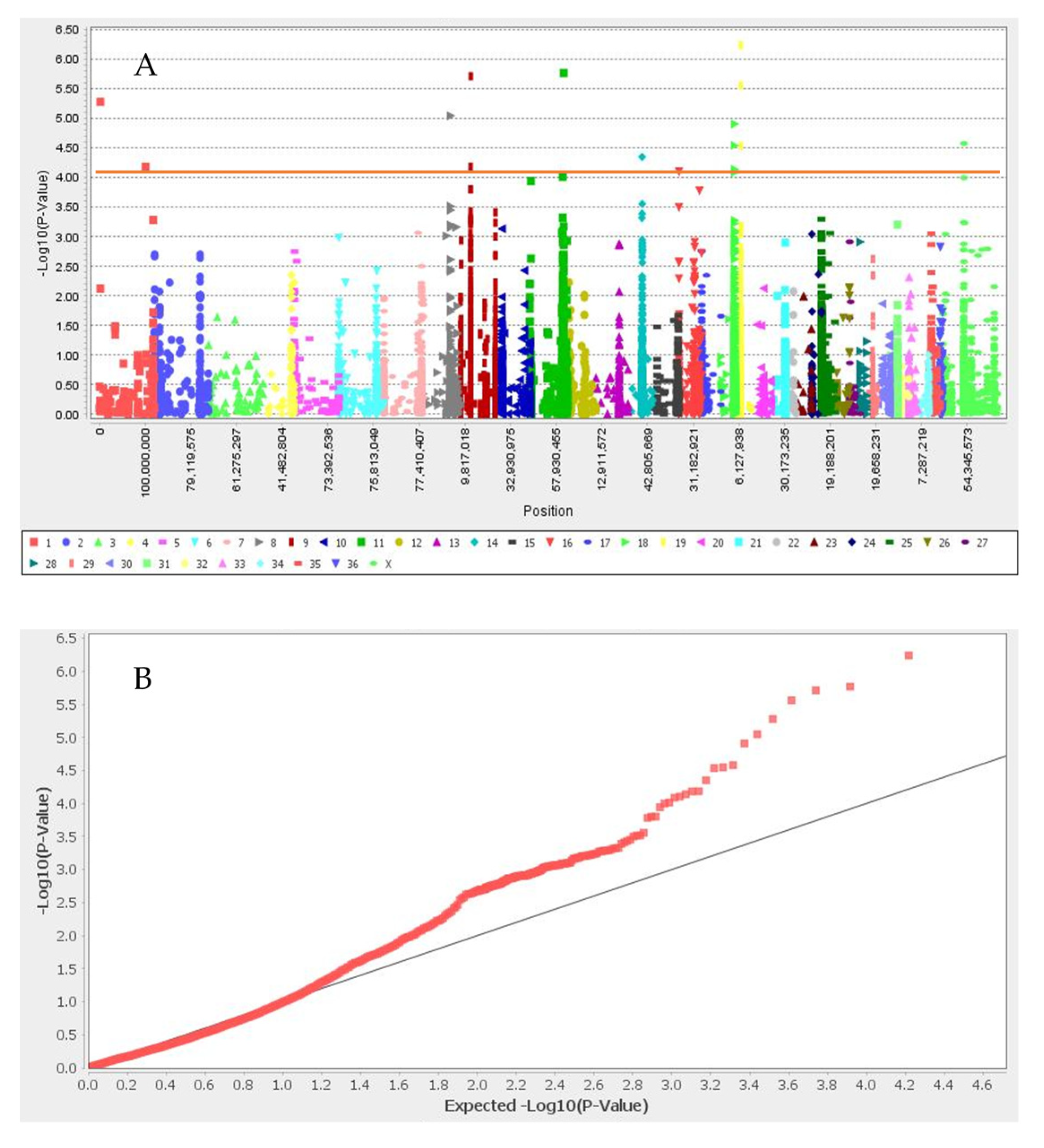
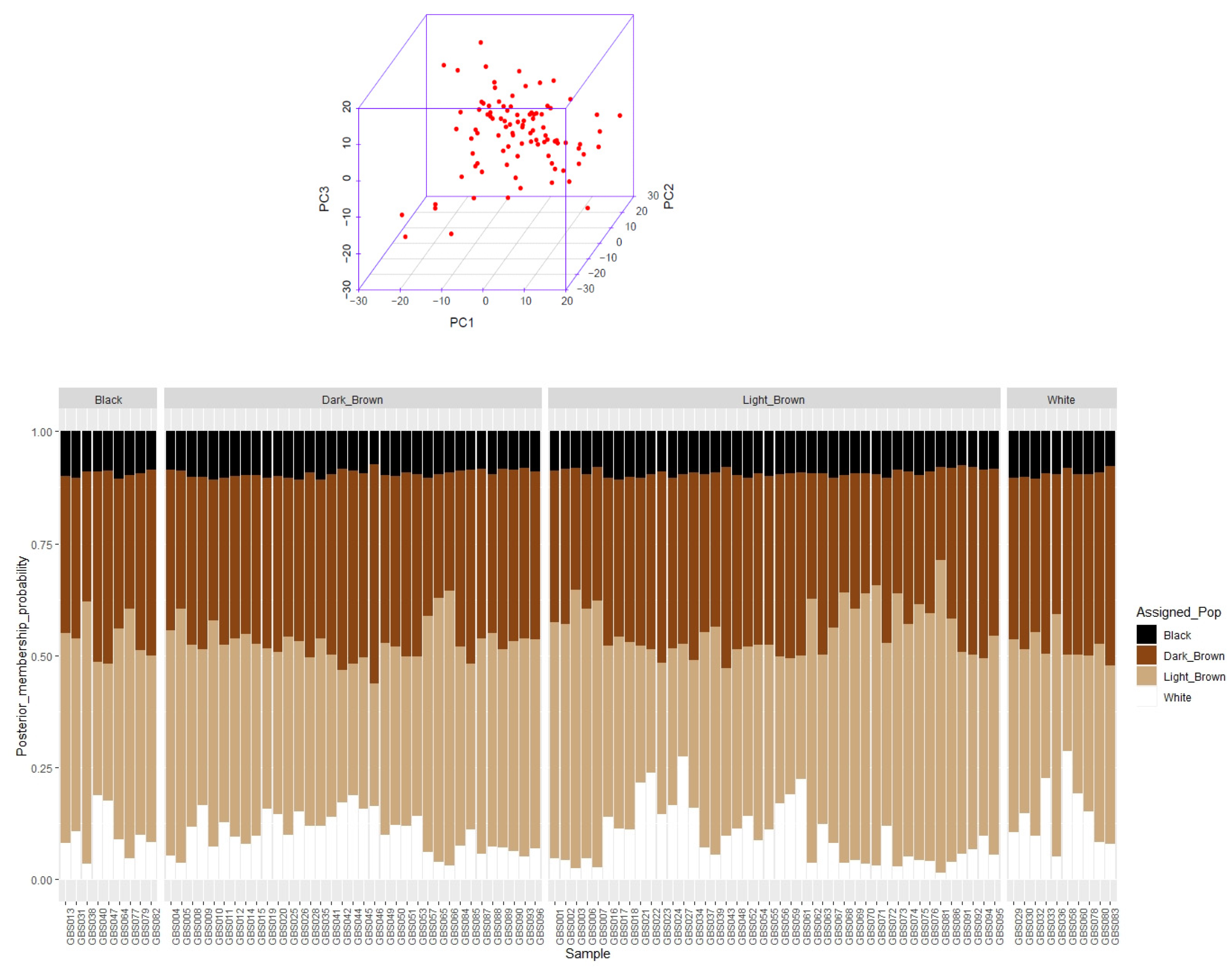
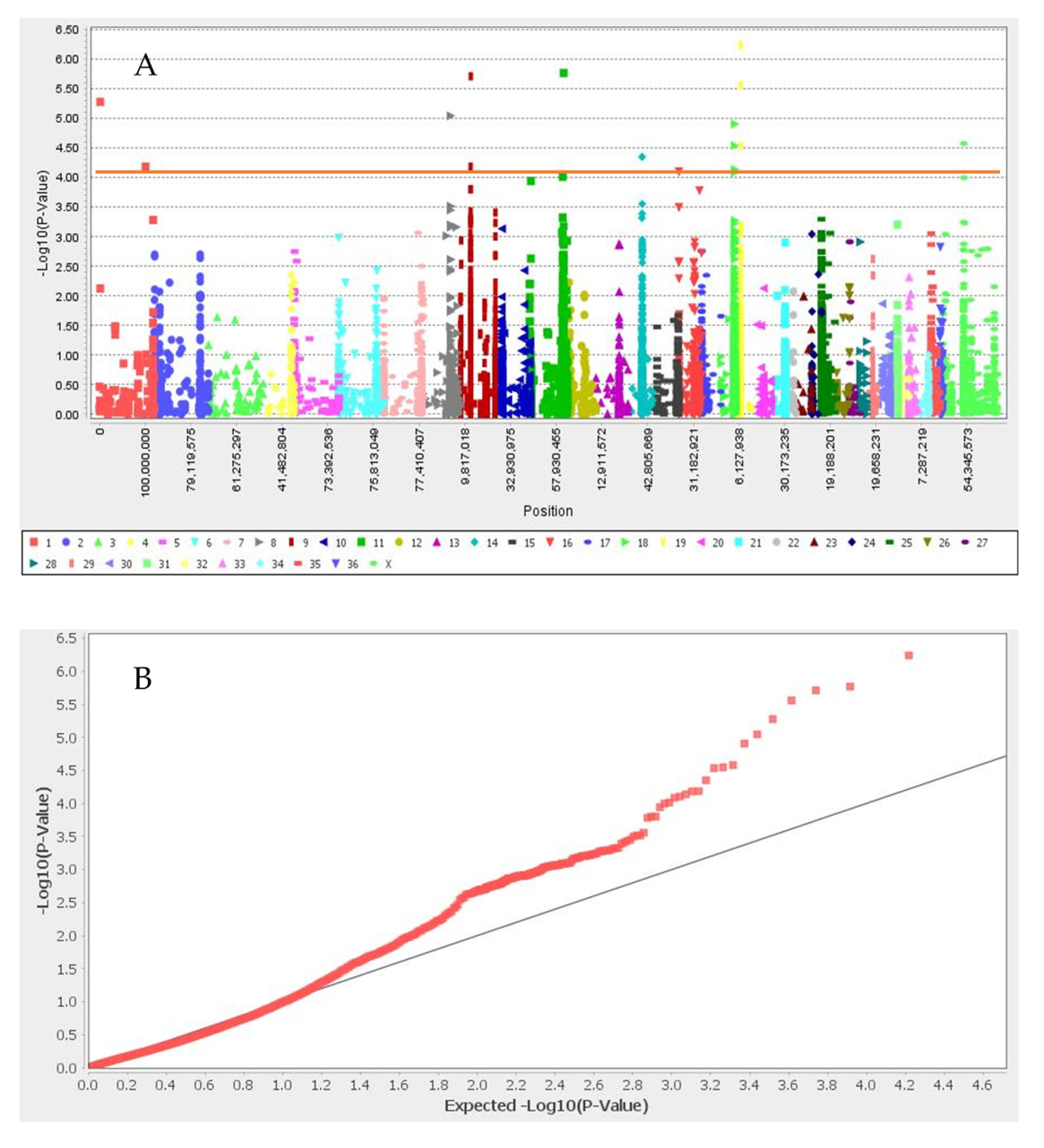
3.1. Summary Statistic of SNPs, Linkage Disequilibrium, Population Structure, and Kinship Analyses
3.2. Genome-Wide Association Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Almathen, F.; Elbir, H.; Bahbahani, H.; Mwacharo, J.; Hanotte, O. Polymorphisms in MC1R and ASIP Genes are Associated with Coat Color Variation in the Arabian Camel. J. Hered. 2018, 109, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Peters, K.J.; Horst, P.; Kleinheisterkamp, H.H. The importance of coat colour and coat type as indicators of productive adaptability of beef cattle in a subtropical environment. Trop. Anim. Prod. 1982, 7, 296–304. [Google Scholar]
- Giantsis, I.A.; Antonopoulou, D.; Dekolis, N.; Zaralis, K.; Avdi, M. Origin, demographics, inbreeding, phylogenetics, and phenogenetics of Karamaniko breed, a major common ancestor of the autochthonous Greek sheep. Trop. Anim. Health Prod. 2022, 54, 73. [Google Scholar] [CrossRef] [PubMed]
- Finch, V.A.; Bennett, I.L.; Holmes, C.R. Coat colour in cattle: Effect on thermal balance, behaviour and growth, and relationship with coat type. J. Agric. Sci. 1984, 102, 141–147. [Google Scholar] [CrossRef]
- Ward, J.M.; Blount, J.D.; Ruxton, G.D.; Houston, D.C. The adaptive significance of dark plumage for birds in desert environments. Ardea 2002, 90, 311–323. [Google Scholar]
- Traoré, B.; Moula, N.; Toure, A.; Ouologuem, B.; Leroy, P.; Antoine-Moussiaux, N. Characterisation of camel breeding practices in the Ansongo Region, Mali. Trop. Anim. Health Prod. 2014, 46, 1303–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Swailem, A.M.; Al-Busadah, K.A.; Shehata, M.M.; Al-Anazi, I.O.; Askari, E. Classification of Saudi Arabian camel (Camelus dromedarius) subtypes based on RAPD technique. J. Food Agric. Environ. 2007, 5, 143–148. [Google Scholar]
- Hussein, Y.A.; Al-Eknah, M.M.; Al-Shami, S.A.; Mandour, M.A.; Fouda, T.A. Coat color breed variation in blood constituents among indigenous Saudi Arabia camel strains. Mansoura Vet. Med. J. 2012, 14, 191–204. [Google Scholar]
- Sturm, A.R.; Teasdale, R.D.; Box, N.F. Human pigmentation genes: Identification, structure and consequences of polymorphic variation. Gene 2001, 277, 49–62. [Google Scholar] [CrossRef]
- Cieslak, M.; Reissmann, M.; Hofreiter, M.; Ludwig, A. Colours of domestication. Biol. Rev. 2011, 86, 885–899. [Google Scholar] [CrossRef]
- Geissler, E.N.; Ryan, M.A.; Housman, D.E. The dominant-white spotting (W) locus of the mouse encodes the c-kit proto-oncogene. Cell 1988, 55, 185–192. [Google Scholar] [CrossRef]
- Marklund, S.; Kijas, J.; Rodriguez-Martinez, H.; Rönnstrand, L.; Funa, K.; Moller, M.; Lange, D.; Edfors-Lilja, I.; Andersson, L. Molecular Basis for the Dominant White Phenotype in the Domestic Pig. Genome Res. 1998, 8, 826–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pielberg, G.; Olsson, C.; Syvanen, A.C.; Andersson, L. Unexpectedly high allelic diversity at the KIT locus causing dominant white color in the domestic pig. Genetics 2002, 160, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Haase, B.; Brooks, S.A.; Tozaki, T.; Burger, D.; Poncet, P.A.; Rieder, S.; Hasegawa, T.; Penedo, C.; Leeb, T. Seven novel KIT mutations in horses with white coat colour phenotypes. Anim. Genet. 2009, 40, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Haase, B.; Jagannathan, V.; Rieder, S.; Leeb, T. A novel KIT variant in an Icelandic horse with white-spotted coat colour. Anim. Genet. 2015, 46, 466. [Google Scholar] [CrossRef] [PubMed]
- Fontanesi, L.; Beretti, F.; Riggio, V.; Dall’Olio, S.; González, E.G.; Finocchiaro, R.; Davoli, R.; Russo, V.; Portolano, B. Missense and nonsense mutations in melanocortin 1 receptor (MC1R) gene of different goat breeds: Association with red and black coat colour phenotypes but with unexpected evidences. BMC Genet. 2009, 10, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatis, C.; Giannoulis, T.; Galliopoulou, E.; Billinis, C.; Mamuris, Z. Genetic analysis of melanocortin 1 receptor gene in endangered Greek sheep breeds. Small Rumin. Res. 2017, 157, 54–56. [Google Scholar] [CrossRef]
- Fontanesi, L.; D’Alessandro, E.; Scotti, E.; Liotta, L.; Crovetti, A.; Chiofalo, V.; Russo, V. Genetic heterogeneity and selection signature at the KIT gene in pigs showing different coat colours and patterns. Anim. Genet. 2010, 41, 478–492. [Google Scholar] [CrossRef]
- Wong, A.K.; Ruhe, A.L.; Robertson, K.R.; Loew, E.R.; Williams, D.C.; Neff, M.W. Ade novomutation inKITcauses white spotting in a subpopulation of German Shepherd dogs. Anim. Genet. 2012, 44, 305–310. [Google Scholar] [CrossRef]
- David, V.A.; Menotti-Raymond, M.; Wallace, A.C.; Roelke, M.; Kehler, J.; Leighty, R.; Eizirik, E.; Hannah, S.S.; Nelson, G.; Schäffer, A.A.; et al. Endogenous retrovirus insertion in the KIT oncogene determines white and white spotting in domestic cats. G3: Genes Genomes Genet. 2014, 4, 1881–1891. [Google Scholar] [CrossRef] [Green Version]
- Dürig, N.; Jude, R.; Holl, H.; Brooks, S.A.; Lafayette, C.; Jagannathan, V.; Leeb, T. Whole genome sequencing reveals a novel deletion variant in the KIT gene in horses with white spotted coat colour phenotypes. Anim. Genet. 2017, 48, 483–485. [Google Scholar] [CrossRef] [PubMed]
- Holl, H.; Isaza, R.; Mohamoud, Y.; Ahmed, A.; Almathen, F.; Youcef, C.; Gaouar, S.; Antczak, D.F.; Brooks, S. A Frameshift Mutation in KIT is Associated with White Spotting in the Arabian Camel. Genes. 2017, 8, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Searl, A.G. Comparative Genetics of Coat Colour in Mammals; Logos: London, UK, 1968. [Google Scholar]
- Qureshi, A.S.; Shah, M.G.; Reiβmann, M.; Schwartz, H.J. Differentiation of six Pakistani camel breeds by molecular genetics analysis. In Proceedings of the ICAR Satellite Meeting on Camelid Reproduction, Budapest, Hungary, 12–13 July 2008; pp. 61–65. [Google Scholar]
- Cransberg, R.; Munyard, K.A. Polymorphisms detected in the tyrosinase and matp (slc45a2) genes did not explain coat colour dilution in a sample of Alpaca (Vicugna pacos). Small Rumin. Res. 2011, 95, 92–96. [Google Scholar] [CrossRef]
- Chandramohan, B.; Renieri, C.; La Manna, V.; La Terza, A. The alpaca agouti gene: Genomic locus, transcripts and causative mutations of eumelanic and pheomelanic coat color. Gene 2013, 521, 303–310. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. 2019. Available online: http://faostat3.fao.org/download/Q/QC/E (accessed on 21 January 2015).
- Javanrouh, A.; Banabazi, M.H.; Esmaeilkhanian, S.; Amirinia, C.; Seyedabadi, H.R.; Emrani, H. Optimization on salting out method for DNA extraction from animal and poultry blood cells. In Proceedings of the 57th Annual Meeting Of The European Association For Animal Production, Antalya, Turkey, 17–20 September 2006. [Google Scholar]
- Elbers, J.P.; Rogers, M.F.; Perelman, P.L.; Proskuryakova, A.A.; Serdyukova, N.A.; Johnson, W.E.; Horin, P.; Corander, J.; Murphy, D.; Burger, P.A. Improving Illumina assemblies with Hi-C and long reads: An example with the North African dromedary. Mol. Ecol. Resour. 2019, 19, 1015–1026. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows–Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pembleton, L.W.; Cogan, N.O.I.; Forster, J.W. StAMPP: An R package for calculation of genetic differentiation and structure of mixed-ploidy level populations. Mol. Ecol. Resour. 2013, 13, 946–952. [Google Scholar] [CrossRef]
- Knaus, B.J.; Grünwald, N.J. vcfr: A package to manipulate and visualize variant call format data in R. Mol. Ecol. Resour. 2017, 17, 44–53. [Google Scholar] [CrossRef]
- Kamvar, Z.; Tabima, J.; Grunwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of Phylogenetics and Evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuwirth, E.; Neuwirth, M.E. Package ‘RColorBrewer’; CRAN 2011-06-17 08:34:00. Apache License 2.0; R Core Team: Vienna, Austria, 2011. [Google Scholar]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Mering, C.V.; Huynen, M.; Jaeggi, D.; Schmidt, S.; Bork, P.; Snel, B. STRING: A database of predicted functional associations between proteins. Nucleic Acids Res. 2003, 31, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Alshanbari, F.; Castaneda, C.; Juras, R.; Hillhouse, A.; Mendoza, M.N.; Gutiérrez, G.A.; De León, F.A.P.; Raudsepp, T. Comparative FISH-Mapping of MC1R, ASIP, and TYRP1 in New and Old World Camelids and Association Analysis With Coat Color Phenotypes in the Dromedary (Camelus dromedarius). Front. Genet. 2019, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Klungland, H.; Våge, D.I.; Gomez-Raya, L.; Adalsteinsson, S.; Lien, S. The role of melanocyte-stimulating hormone (MSH) receptor in bovine coat color determination. Mamm. Genome 1995, 6, 636–639. [Google Scholar] [CrossRef] [PubMed]
- Joerg, H.; Fries, H.R.; Meijerink, E.; Stranzinger, G.F. Red coat color in Holstein cattle is associated with a deletion in the MSHR gene. Mamm. Genome 1996, 7, 317–318. [Google Scholar] [CrossRef]
- Våge, D.I.; Klungland, H.; Lu, D.; Cone, R.D. Molecular and pharmacological characterization of dominant black coat color in sheep. Mamm. Genome 1999, 10, 39–43. [Google Scholar] [CrossRef]
- Switonski, M.; Mankowska, M.; Salamon, S. Family of melanocortin receptor (MCR) genes in mammals—mutations, polymorphisms and phenotypic effects. J. Appl. Genet. 2013, 54, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Adalsteinsson, S.; Sponenberg, D.P.; Alexieva, S.; Russel, A.J. Inheritance of goat coat colors. J. Hered. 1994, 85, 267–272. [Google Scholar] [CrossRef]
- Martin, P.M.; Palhière, I.; Ricard, A.; Tosser-Klopp, G.; Rupp, R. Genome Wide Association Study Identifies New Loci Associated with Undesired Coat Color Phenotypes in Saanen Goats. PLoS ONE 2016, 11, e0152426. [Google Scholar] [CrossRef]
- Norris, B.J.; Whan, V.A. A gene duplication affecting expression of the ovine ASIP gene is responsible for white and black sheep. Genome Res. 2008, 18, 1282–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, M.G.; Reissmann, M.; Qureshi, A.S.; Schwartz, H.J. Sequencing and mutation screening in exon 1 of camel tyrosinase gene. The global food & product chain-dynamics, innovations, conflicts, strategies; Deutscher Tropentag: Hohenheim, Germany, 11–13 October 2005. [Google Scholar]
- Schmidtz, B.H.; Buchanan, F.C.; Plante, Y.; Schmutz, S.M. Linkage mapping of Tyrosinase gene to bovine chromosome 26. Anim. Genet. 2001, 32, 119–120. [Google Scholar] [CrossRef] [PubMed]
- Bitaraf Sani, M.; Harofte, J.Z.; Bitaraf, A.; Esmaeilkhanian, S.; Banabazi, M.H.; Salim, N.; Teimoori, A.; Naderi, A.S.; Faghihi, M.A.; Burger, P.A.; et al. Genome-Wide Diversity, Population Structure and Demographic History of Dromedaries in the Central Desert of Iran. Genes 2020, 11, 599. [Google Scholar] [CrossRef] [PubMed]
- Morante, R.; Goyache, F.; Burgos, A.; Cervantes, I.; Pérez-Cabal, M.; Gutiérrez, J. Genetic improvement for alpaca fibre production in the Peruvian Altiplano: The Pacomarca experience. Anim. Genet. Resour. Inf. 2009, 45, 37–43. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Coat Color | SNP | Chr | Pos | −log (p-Value) | Candidate Genes ± 100 kb |
|---|---|---|---|---|---|
| White | Chr25_73462 | 25 | 734,62 | 5.84 | - |
| Chr35_5648539 | 35 | 5,648,539 | 5.39 | ANKRD26, GNB1 | |
| Chr18_29916170 | 18 | 29,916,170 | 4.78 | TSPYL4, TEKT5, DEXI, CIITA, TVP23B, CLEC16A | |
| Chr33_12297059 | 33 | 12,297,059 | 4.71 | TMPRSS13, FXYD6, MPZL3, ANKRD26 | |
| Chr35_5696438 | 35 | 5,696,438 | 4.40 | - | |
| Chr9_1807083 | 9 | 1,807,083 | 4.37 | HFM1, CDC7, TGFBR3 | |
| Chr10_74907708 | 10 | 74,907,708 | 4.19 | - | |
| Chr8_31826040 | 8 | 31,826,040 | 4.17 | HACE1 | |
| Chr25_505194 | 25 | 505,194 | 4.16 | - | |
| Black | Chr19_10157184 | 19 | 10,157,184 | 6.24 | UBE2V1, TMEM189, SNAI1 |
| Chr11_74286851 | 11 | 74,286,851 | 5.77 | SEC23IP, MCMBP, INPP5F | |
| Chr9_22746017 | 9 | 22,746,017 | 5.71 | trpS | |
| Chr19_10612243 | 19 | 10,612,243 | 5.56 | CREB3L1, DGKZ, MDK, CHRM4 | |
| Chr1_2037209 | 1 | 2,037,209 | 5.28 | COL4A4, COL6A6, CAPN7, EAF1 | |
| Chr8_59919441 | 8 | 59,919,441 | 5.05 | ||
| Chr18_29898490 | 18 | 29,898,490 | 4.91 | TSPYL4, TEKT5, DEXI, CIITA, TVP23B, CLEC16A | |
| Chrx_46816486 | X | 46,816,486 | 4.58 | HDLBP | |
| Chr18_29898527 | 18 | 29,898,527 | 4.55 | TSPYL4, TEKT5, DEXI, CIITA, TVP23B, CLEC16A | |
| Chr19_10612244 | 19 | 10,612,244 | 4.54 | CREB3L1, DGKZ, MDK, CHRM4 | |
| Chr14_30853969 | 14 | 30,853,969 | 4.35 | - | |
| Chr9_22614201 | 9 | 22,614,201 | 4.19 | TBX15, trpS | |
| Chr1_101414918 | 1 | 1.01 × 108 | 4.18 | - | |
| Light and dark brown | No associated SNP | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bitaraf Sani, M.; Zare Harofte, J.; Banabazi, M.H.; Faraz, A.; Esmaeilkhanian, S.; Naderi, A.S.; Salim, N.; Teimoori, A.; Bitaraf, A.; Zadehrahmani, M.; et al. Identification of Candidate Genes for Pigmentation in Camels Using Genotyping-by-Sequencing. Animals 2022, 12, 1095. https://doi.org/10.3390/ani12091095
Bitaraf Sani M, Zare Harofte J, Banabazi MH, Faraz A, Esmaeilkhanian S, Naderi AS, Salim N, Teimoori A, Bitaraf A, Zadehrahmani M, et al. Identification of Candidate Genes for Pigmentation in Camels Using Genotyping-by-Sequencing. Animals. 2022; 12(9):1095. https://doi.org/10.3390/ani12091095
Chicago/Turabian StyleBitaraf Sani, Morteza, Javad Zare Harofte, Mohammad Hossein Banabazi, Asim Faraz, Saeid Esmaeilkhanian, Ali Shafei Naderi, Nader Salim, Abbas Teimoori, Ahmad Bitaraf, Mohammad Zadehrahmani, and et al. 2022. "Identification of Candidate Genes for Pigmentation in Camels Using Genotyping-by-Sequencing" Animals 12, no. 9: 1095. https://doi.org/10.3390/ani12091095
APA StyleBitaraf Sani, M., Zare Harofte, J., Banabazi, M. H., Faraz, A., Esmaeilkhanian, S., Naderi, A. S., Salim, N., Teimoori, A., Bitaraf, A., Zadehrahmani, M., Burger, P. A., Asadzadeh, N., Silawi, M., Taghipour Sheshdeh, A., Mohammad Nazari, B., Faghihi, M. A., & Roudbari, Z. (2022). Identification of Candidate Genes for Pigmentation in Camels Using Genotyping-by-Sequencing. Animals, 12(9), 1095. https://doi.org/10.3390/ani12091095