
Impact of Heat Stress on Bovine Sperm Quality and Competence



{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction—The Climate Change Scenario
2. Adaptive Mechanisms to Heat Stress in Cattle
3. Cell Response to Heat Stress
Heat Shock Proteins (HSPs)
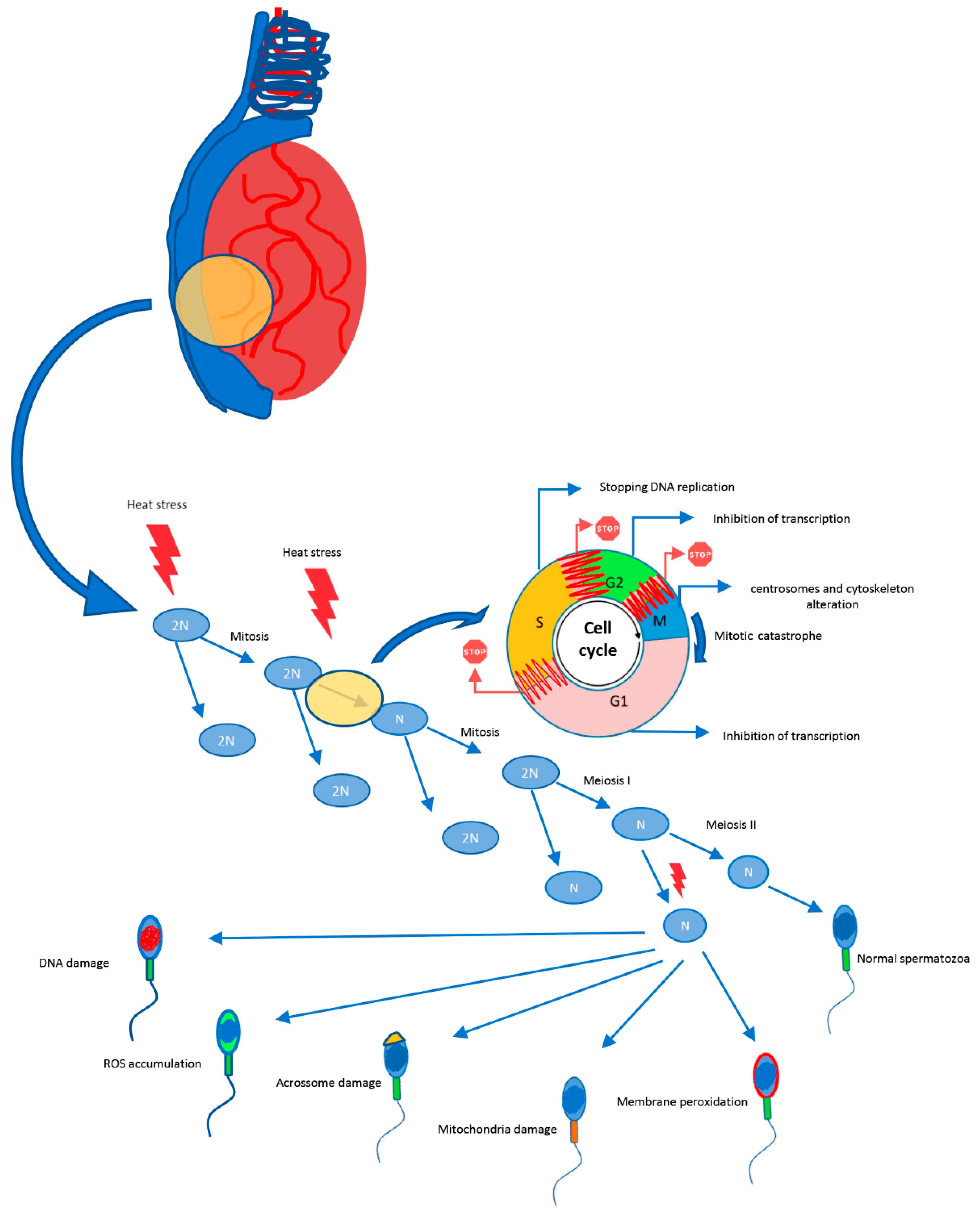
4. Impact of Heat Stress on Spermatogenesis
4.1. Oxidative Stress
4.2. Apoptosis
4.3. DNA Damage
5. Heat Stress Effects on Spermatozoa Capacitation and Fertilization Ability
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- IPCC. Summary for Policymakers. In Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Shukla, P.R., Skea, J., Buendia, E.C., Masson-Delmotte, V., Pörtner, H.-O., Roberts, D.C., Zhai, P., Slade, R., Connors, S., van Diemen, R., et al., Eds.; IPCC: Geneva, Switzerland, 2019; in press. [Google Scholar]
- Rashamol, V.P.; Sejian, V. Climate resilient livestock production: Way forward. Dairy Vet. Sci. 2018, 5, 5556673. [Google Scholar] [CrossRef]
- NASA. Available online: https://www.nasa.gov/press-release/2021-tied-for-6th-warmest-year-in-continued-trend-nasa-analysis-shows (accessed on 20 February 2021).
- Rockström, J.; Steffen, W.; Noone, K.; Persson, A.; Chapin, F.S., 3rd; Lambin, E.; Lenton, T.M.; Scheffer, M.; Folke, C.; Schellnhuber, H.J.; et al. A safe operating space for humanity. Nature 2009, 461, 472–475. [Google Scholar] [CrossRef] [PubMed]
- Umberto, B. Climate change: Impact on livestock and how can we adapt. Anim. Front. 2019, 9, 3–5. [Google Scholar] [CrossRef]
- Ravagnolo, O.; Misztal, I. Genetic Component of Heat Stress in Dairy Cattle, Parameter Estimation. J. Dairy Sci. 2000, 83, 2126–2130. [Google Scholar] [CrossRef]
- McManus, C.M.; Paiva, S.R.; Faria, D. Genomics and climate change. Rev. Sci. Tech. 2020, 39, 481–490. [Google Scholar] [CrossRef]
- Hansen, P.J. Physiological and cellular adaptations of zebu cattle to thermal stress. Anim. Reprod. Sci. 2004, 82–83, 349–360. [Google Scholar] [CrossRef]
- Aitken, R.J. The male is significantly implicated as the cause of unexplained infertility. Semin. Reprod. Med. 2020, 38, 3–20. [Google Scholar] [CrossRef]
- Kaushik, K.; Kaushal, N.; Mittal, P.K.; Kalla, N.R. Heat induced differential pattern of DNA fragmentation in male germ cells of rats. J. Biol. 2019, 84, 351–356. [Google Scholar] [CrossRef]
- Xavier, M.J.; Nixon, B.; Roman, S.D.; Scott, R.J.; Drevet, J.R.; Aitken, R.J. Paternal impacts on development: Identification of genomic regions vulnerable to oxidative DNA damage in human spermatozoa. Hum. Reprod. 2019, 34, 1876–1890. [Google Scholar] [CrossRef]
- Rizzoto, G.; Boe-Hansen, G.; Klein, C.; Thundathil, J.C.; Kastelic, J.P. Acute mild heat stress alters gene expression in testes and reduces sperm quality in mice. Theriogenology 2020, 158, 375–381. [Google Scholar] [CrossRef]
- Bligh, J.; Johnson, K.G. Glossary of terms for thermal physiology. J. Appl. Phys. 1973, 35, 941–961. [Google Scholar] [CrossRef] [PubMed]
- Abdelnour, S.A.; El-Hack, M.E.A.; Khafaga, A.F.; Arif, M.; Taha, A.E.; Noreldin, A.E. Stress biomarkers and proteomics alteration to thermal stress in ruminants: A review. J. Therm. Biol. 2019, 79, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Silanikove, N.; Maltz, E.; Halevi, A.; Shinder, D. Metabolism of water, sodium, potassium and chlorine by high yieldingdairy cows at the onset of lactation. J. Dairy Sci. 1997, 80, 949–956. [Google Scholar] [CrossRef]
- Winchester, C.F.; Morris, M.J. Water intake rates of cattle. J. Anim. Sci. 1956, 15, 722–740. [Google Scholar] [CrossRef]
- Beede, D.K.; Collier, R.J. Potential nutritional strategies for intensively managed cattle during heat stress. J. Anim. Sci. 1986, 62, 543–550. [Google Scholar] [CrossRef]
- Nacional Research Council. Nutrient Requirements of Dairy Cattle, 7th ed.; The National Academies Press: Washington, DC, USA, 2001. [Google Scholar]
- Magdub, A.B.; Johnson, H.D.; Belyea, R.L. Effect of environment heat and dietary fiber on thyroid physiology of the lactating cows. J. Dairy Sci. 1982, 65, 2323–2331. [Google Scholar] [CrossRef]
- Silanikove, N. Effects of heat stress on the welfare of extensively managed domestic ruminants. Livest. Prod. Sci. 2000, 67, 1–18. [Google Scholar] [CrossRef]
- Morea, P.; Basirico, L.; Hosoda, K.; Bernabucci, U. Chronic heat stress upregulates leptin and adiponectin secretion and expression and improves leptin, adipoctin and insulin sensitivity in mice. J. Mol. Endocrinol. 2012, 48, 129–138. [Google Scholar] [CrossRef]
- Igono, M.O.; Bjotvet, G.; Sanford-Crane, H.T. Environmental profile and critical temperature effects on milk production of Holstein cows in desert climate. Int. J. Biometeorol. 1992, 36, 77–87. [Google Scholar] [CrossRef]
- Hahn, G.L. Dynamic responses of cattle to thermal heat loads. J. Anim. Sci. 1999, 77 (Suppl. 2), 10–20. [Google Scholar] [CrossRef]
- Johnson, H.D.; Vanjonack, W.J. Effects of environmental and other stressors on blood patterns in lactating animals. J. Dairy Sci. 1976, 59, 1603–1617. [Google Scholar] [CrossRef]
- Bohmanova, J.; Misztal, I.; Cole, J.B. Temperature-Humidity Indices as Indicators of Milk Production Losses due to Heat Stress. J. Dairy Sci. 2007, 90, 1947–1956. [Google Scholar] [CrossRef] [PubMed]
- Brügemann, K.; Gernand, E.; Borstel, U.K.; König, S. Defining and evaluating heat stress thresholds in different dairy cow production systems. Arch. Anim. Breed. 2012, 55, 13–24. [Google Scholar] [CrossRef]
- Menegassi, S.R.O.; Pereira, G.R.; Dias, E.A.; Rocha, M.K.; Carvalho, H.R.; Koetz, C., Jr.; Oberst, E.R.; Barcellos, J.O.J. Infrared thermography as a noninvasive method to assess scrotal insulation on sperm production in beef bulls. Andrologia 2018, 50, e12904. [Google Scholar] [CrossRef]
- McDowell, R.E.; Hooven, N.W.; Camoens, J.K. Effects of climate on performance of Holsteins in first lactation. J. Dairy Sci. 1976, 59, 965–971. [Google Scholar] [CrossRef]
- Lemerle, C.; Goddard, M.E. Assessment of heat stress in dairy cattle in Papua New Guinea. Trop. Anim. Health Prod. 1986, 18, 232–242. [Google Scholar] [CrossRef]
- Allen, T.E.; Pan, Y.S.; Hayman, R.H. The effect of feeding on evaporative heat loss and body temperature in Zebu and Jersey heifers. Aust. J Agric. Res. 1963, 14, 580–593. [Google Scholar] [CrossRef]
- Marti, S.; Wilde, R.E.; Moya, D.; Heuston, C.E.M.; Brown, F.; Schwartzkopf-Genswein, K.S. Effect of rest stop duration during long-distance transport on welfare indicators in recently weaned beef calves. J. Anim. Sci. 2016, 95, 636–644. [Google Scholar] [CrossRef]
- González, L.A.; Schwartzkopf-Genwein, K.S.; Caulkett, N.A.; Janzen, E.; Fierheller, E.; Mcallister, T.A.; Schaefer., A.L.; Haley, D.B.; Stookey, J.M.; Hendrick, S. Pain mitigation following band castration of beef calves and its effects on performance, behavior, E. coli, and salivary cortisol. J. Anim. Sci. 2009, 88, 802–810. [Google Scholar] [CrossRef]
- Petrie, N.; Mellor, D.J.; Stafford, K.J.; Bruce, R.A.; Ward, R.N. Cortisol resposnses of calves to two methods of disbudding used with or without local anaesthetic. N. Z. Vet. J. 1996, 44, 9–14. [Google Scholar] [CrossRef]
- Cook, C.J.; Mellor, D.J.; Harris, P.J.; Ingram, J.R.; Matthews, L.R. Hands-on and Hands-off measurement of stress. In The Biology of Animals Stress; Moberg, G.P., Mench, J.A., Eds.; CAB Internantional: Wallinford, UK, 2000; pp. 123–147. [Google Scholar]
- Moya, D.; Schwartzkopf-Genswein, K.S.; Veira, D.M. Standardization of a non-invasive methodology to measure cortisol in hair of beef cattle. Livest. Sci. 2013, 158, 138–144. [Google Scholar] [CrossRef]
- Uetake, K.; Morita, S.; Sakagami, N.; Yamamoto, K.; Hashimura, S.; Tanaka, T. Hair cortisol levels of lactating dairy cows in cold- and warm-temperate regions in Japan. Anim. Sci. J. 2018, 89, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Fisher, D.D.; Wilson, L.L.; Leach, R.M.; Scholz, R.W. Switch hair as an indicator of magnesium and copper status. Am. J. Vet. Res. 1985, 46, 2235–2240. [Google Scholar]
- Garcia-Oliveros, L.N.; de Arruda, R.P.; Batissaco, L.; Gonzaga, V.H.G.; Nogueira, V.J.M.; Florez-Rodriguez, S.A.; Almeida, F.S.; Alves, M.B.R.; Costa Pinto, S.C.; Nichi, M.; et al. Heat stress effects on bovine sperm cells: A chronological approach to early findings. Int. J. Biometeorol. 2020, 64, 1367–1378. [Google Scholar] [CrossRef]
- Widlak, W.; Vydra, N. The Role of Heat Shock Factors in Mammalian Spermatogenesis. In The Role of Heat Shock Proteins in Reproductive System Development and Function; MacPhee, D., Ed.; Springer: Cham, Switzerland, 2017; Volume 222, pp. 45–65. [Google Scholar] [CrossRef]
- Durairajanayagam, D.; Agarwal, A.; Ong, C. Causes, effects and molecular mechanisms of testicular heat stress. Reprod. BioMed. Online 2015, 30, 14–27. [Google Scholar] [CrossRef]
- Brito, L.F.; Silva, A.E.; Barbosa, R.T.; Kastelic, J.P. Testicular thermoregulation in Bos indicus, crossbred and Bos taurus bulls: Relationship with scrotal, testicular vascular cone and testicular morphology, and effects on semen quality and sperm production. Theriogenology 2004, 61, 511–528. [Google Scholar] [CrossRef]
- Morrell, J.M. Heat stress and bull fertility. Theriogenology 2020, 153, 62–67. [Google Scholar] [CrossRef]
- Cameron, R.D.A.; Blackshaw, A.W. The effect of elevated ambient temperature on spermatogenesis in the boar. J. Reprod. Fert. 1980, 53, 173–179. [Google Scholar] [CrossRef]
- England, K.; O’Driscoll, C.; Cotter, T.G. Carbonylation of glycolytic proteins is a key response to drug-induced oxidative stress and apoptosis. Cell Death Differ. 2004, 11, 252–260. [Google Scholar] [CrossRef]
- Slimen, I.B.; Najar, T.; Ghram, A.; Abdrrabba, M. Heat stress effects on livestock: Molecular, cellular and metabolic aspects, a review. J. Anim. Physiol. Nutr. 2016, 100, 401–412. [Google Scholar] [CrossRef]
- Harmon, B.V.; Corder, A.M.; Collins, R.J.; Gobé, G.C.; Allen, J.; Allan, D.J.; Kerr, J.F. Cell death induced in a murine mastocytoma by 42–47 degrees C heating in vitro: Evidence that the form of death changes from apoptosis to necrosis above a critical heat load. Int. J. Radiat. Biol. 1990, 58, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Sorokina, I.V.; Denisenko, T.V.; Imreh, G.; Tyurin-Kuzmin, P.A.; Kaminskyy, V.O.; Gogvadze, V.; Zhivotovsky, B. Involvement of autophagy in the outcome of mitotic catastrophe. Sci. Rep. 2017, 6, 14571. [Google Scholar] [CrossRef] [PubMed]
- Thompson, S.M.; Callstrom, M.R.; Butters, K.A.; Knudsen, B.; Grande, J.P.; Roberts, L.R.; Woodrum, D.A. Heat stress induced cell death mechanisms in hepatocytes and hepatocellular carcinoma: In vitro and in vivo study. Lasers Surg. Med. 2014, 46, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Petrova, N.V.; Velichkoa, A.K.; Razin, S.V.; Kantidze, O.L. Early S-phase cell hypersensitivity to heat stress. Cell Cycle 2016, 15, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Kühl, N.M.; Rensing, L. Heat shock effects on cell cycle progression. Cell. Mol. Life Sci. 2000, 57, 450–463. [Google Scholar] [CrossRef]
- Rappa, F.; Farina, F.; Zummo, G.; David, S.; Campanella, C.; Carini, F.; Tomasello, G.; Damiani, P.; Cappello, F.; De Macario, E.C.; et al. HSP-molecular chaperones in cancer biogenesis and tumor therapy: An overview. Anticancer Res. 2012, 32, 5139–5150. [Google Scholar]
- Bukau, B.; Weissman, J.; Horwich, A. Molecular chaperones and protein quality control. Cell 2016, 125, 443–451. [Google Scholar] [CrossRef]
- Rosenzweig, R.; Nillegoda, N.; Mayer, M.; Bukau, B. The Hsp70 chaperone network. Nat. Rev. Mol. Cell Biol. 2019, 20, 665–680. [Google Scholar] [CrossRef]
- Wang, Y.; Jia, C.; Li, Q.S.; Xie, C.Y.; Zhang, N.; Qu, Y. BAG-1L Protects SH-SY5Y neuroblastoma cells against hypoxia/re-oxygenation through up-regulating HSP70 and activating PI3K/AKT signaling pathway. Neurochem. Res. 2017, 42, 2861–2868. [Google Scholar] [CrossRef]
- Quintana, F.J.; Cohen, I.R. The Hsp60 immune system network. Trends Immunol. 2013, 32, 89–95. [Google Scholar] [CrossRef]
- Shevtsov, M.; Multhoff, G. Heat shock protein–peptide and HSP-based immunotherapies for the treatment of cancer. Front. Immunol. 2016, 29, 171. [Google Scholar] [CrossRef] [PubMed]
- Garrido, C.; Solary, E. A role of HSPs in apoptosis through “protein triage”? Cell Death Differ. 2003, 10, 619–620. [Google Scholar] [CrossRef] [PubMed]
- Velichko, A.K.; Markova, E.N.; Petrova, N.V.; Razin, S.V.; Kantidze, O.L. Mechanisms of heat shock response in mammals. Cell. Mol. Life Sci. 2013, 70, 4229–4241. [Google Scholar] [CrossRef] [PubMed]
- Parshina, E.Y.; Yusipovich, A.I.; Brazhe, A.R.; Silicheva, M.A.; Maksimov, G.V. Heat damage of cytoskeleton in erythrocytes increases membrane roughness and cell rigidity. J. Biol. Phys. 2019, 45, 367–377. [Google Scholar] [CrossRef]
- Grad, I.; Cederroth, C.R.; Walicki, J.; Grey, C.; Barluenga, S.; Winssinger, N.; De Massy, B.; Nef, S.; Picard, D. The molecular chaperone Hsp90alpha is required for meiotic progression of spermatocytes beyond pachytene in the mouse. PLoS ONE 2010, 5, e15770. [Google Scholar] [CrossRef]
- Kajiwara, C.; Kondo, S.; Uda, S.; Dai, L.; Ichiyanagi, T.; Chiba, T.; Ishido, S.; Koji, T.; Udono, H. Spermatogenesis arrest caused by conditional deletion of Hsp90a in adult mice. Biol. Open 2012, 1, 977–982. [Google Scholar] [CrossRef]
- Zhang, X.G.; Hu, S.; Han, C.; Zhu, Q.C.; Yan, G.J.; Hu, J.H. Association of heat shock protein 90 with motility of post-thawed sperm in bulls. Cryobiology 2015, 70, 164–169. [Google Scholar] [CrossRef]
- Ramon, M.; Salces-Ortiz, J.; Gonzalez, C.; Perez-Guzman, M.D.; Garde, J.J.; García-Álvarez, O.; Maroto-Morales, A.; Calvo, J.H.; Serrano, M.M. Influence of the temperature and the genotype of the HSP90AA1 gene over sperm chromatin stability in Manchega rams. PLoS ONE 2014, 9, e86107. [Google Scholar] [CrossRef]
- Paul, C.; Murray, A.A.; Spears, N.; Saunders, P.T.K. A single, mild, transient scrotal heat stress causes DNA damage, subfertility and impairs formation of blastocysts in mice. Reproduction 2008, 136, 73–84. [Google Scholar] [CrossRef]
- Rahman, M.B.; Vandaelea, L.; Rijsselaerea, T.; Maesa, D.; Hoogewijsa, M.; Frijtersb, A.; Noordmanb, J.; Granadosc, A.; Dernellec, E.; Shamsuddin, M. Scrotal insulation and its relationship to abnormal morphology, chromatin protamination and nuclear shape of spermatozoa in Holstein-Friesian and Belgian Blue bulls. Theriogenology 2011, 76, 1246–1257. [Google Scholar] [CrossRef]
- Pérez-Crespo, M.; Pintado, B.; Gutiérrez-Adán, A. Scrotal Heat Stress Effects on Sperm Viability, Sperm DNA Integrity, and the Offspring Sex Ratio in Mice. Mol. Reprod. Dev. 2008, 75, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Sabés-Alsina, M.; Lundeheim, N.; Johannisson, A.; López-Béjar, M.; Morrell, J.M. Relationships between climate and sperm quality in dairy bull semen: A retrospective analysis. J. Dairy Sci. 2019, 102, 5623–5633. [Google Scholar] [CrossRef]
- Luceño, N.L.; Angrimani, D.S.R.; Bicudo, L.C.; Szymanska, K.J.; Van Poucke, M.; Demeyere, K.; Meyer, E.; Peelman, L.; Mullaart, E.; Broekhuijse, M.L.W.J.; et al. Exposing dairy bulls to high temperature-humidity index during spermatogenesis compromises subsequent embryo development in vitro. Theriogenology 2020, 141, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.B.; Schellander, K.; Luceño, N.L.; Soom, A.V. Heat stress responses in spermatozoa: Mechanisms and consequences for cattle fertility. Theriogenology 2018, 113, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Paul, C.; Teng, S.; Saunders, P.T.K. A Single, Mild, Transient Scrotal Heat Stress Causes Hypoxia and Oxidative Stress in Mouse Testes, Which Induces Germ Cell Death. Biol. Reprod. 2009, 80, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Rizzoto, G.; Kastelic, J.P. A new paradigm regarding testicular thermoregulation in ruminants? Theriogenology 2020, 147, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Gibb, Z.; Baker, M.A.; Drevet, J.; Gharagozloo, P. Causes and consequences of oxidative stress in spermatozoa. Reprod. Fertil. Dev. 2016, 28, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Park, K.; Rhee, K. Heat stress response of male germ cells. Cell. Mol. Life Sci. 2013, 70, 2623–2636. [Google Scholar] [CrossRef]
- Houston, B.J.; Nixon, B.; Martin, J.H.; De Luliis, G.N.; Trigg, N.A.; Bromfield, E.G.; McEwan, K.E.; Aitken, R.J. Heat exposure induces oxidative stress and DNA damage in the male germ line. Biol. Reprod. 2018, 98, 593–606. [Google Scholar] [CrossRef]
- Leite, R.F.; Losano, J.D.A.; Kawai, G.K.V.; Rui, B.R.; Nagai, K.K.; Castiglioni, V.C.; Siqueira, A.F.P.; D’Avila Assumpção, M.E.O.; Baruselli, P.S.; Nichi, M. Sperm function and oxidative status: Effect on fertility in Bos taurus and Bos indicus bulls when semen is used for fixed-time artificial insemination. Anim. Reprod. Sci. 2022, 237, 106922. [Google Scholar] [CrossRef]
- de Castro, L.S.; de Assis, P.M.; Siqueira, A.F.P.; Hamilton, T.R.S.; Mendes, C.M.; Losano, J.D.A.; Nichi, M.; Visintin, J.A.; Assumpção, M.E.O.A. Sperm Oxidative Stress Is Detrimental to Embryo Development: A Dose-Dependent Study Model and a New and More Sensitive Oxidative Status Evaluation. Oxid. Med. Cell. Longev. 2016, 2016, 8213071. [Google Scholar] [CrossRef] [PubMed]
- Garret, L.J.A.; Revell, S.G.; Leese, H.J. Adenosine Triphosphate Production by Bovine Spermatozoa and Its Relationship to Semen Fertilizing Ability. J. Androl. 2008, 29, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Sugimura, S.; Matoba, S.; Hashiyada, Y.; Aikawa, Y.; Ohtake, M.; Matsuda, H.; Kobayashi, S.; Konishi, K.; Imai, K. Oxidative phosphorylation-linked respiration in individual bovine oocytes. J. Reprod. Dev. 2012, 58, 636–641. [Google Scholar] [CrossRef] [PubMed]
- Christen, F.; Desrosiers, V.; Dupont-Cyr, B.A.; Vandenberg, G.W.; Le François, N.R.; Tardif, J.; Dufresne, F.; Lamarre, S.G.; Blier, P.U. Thermal tolerance and thermal sensitivity of heart mitochondria: Mitochondrial integrity and ROS production. Free Radic. Biol. Med. 2018, 116, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Lucio, A.C.; Alves, B.G.; Alves, K.A.; Martins, M.C.; Braga, L.S.; Miglio, L.; Alves, B.G.; Silva, T.H.; Jacomini, J.O.; Beletti, M.E. Selected sperm traits are simultaneouslyaltered after scrotal heat stress and play specific roles in in vitrofertilization and embryonic development. Theriogenology 2016, 86, 924–933. [Google Scholar] [CrossRef]
- De, K.; Kumar, D.; Balaganur, K.; Saxena, V.K.; Thirumurugan, P.; Naqvi, S.M.K. Effect of thermal exposure on physiological adaptability and seminal attributes of rams under semi-arid environment. J. Biol. 2017, 65, 113–118. [Google Scholar] [CrossRef]
- Teixeira, J.; Deus, C.M.; Borges, F.; Oliveira, P.J. Mitochondria: Targeting mitochondrial reactive oxygen species with mitochondriotropic polyphenolic-based antioxidants. Int. J. Biochem. Cell Biol. 2018, 97, 98–103. [Google Scholar] [CrossRef]
- Teixeira, C.; Marques, C.C.; Baptista, M.C.; Pimenta, J.; Teixeira, J.; Cagide, F.; Borges, F.; Montezinho, L.; Oliveira, P.; Pereira, R.M.L.N. Effect of a new mitochondriotropic antioxidant on oocyte maturation and embryo production. In Book of Abstracts of the 71st Annual Meeting of the European Federation of Animal Science Virtual Meeting—1 to 4 December; Scientific Committee: Rome, Italy, 2020; p. 672. [Google Scholar] [CrossRef]
- Montezinho, L.C.P.; Santos, J.; Marques, C.; Baptista, M.C.; Pimenta, J.; Teixeira, J.; Cagide, F.; Borges, F.; Oliveira, P.J.; Pereira, R.M. Impacts of a novel mitochondrial-directed antioxidant on bovine sperm function and in vitro embryo development. Eur. J. Soc. Clin. Investig. 2021, 51, 37. [Google Scholar]
- Vera, Y.; Diaz-Romero, M.; Rodriguez, S.; Lue, Y.; Wang, C.; Swerdloff, R.S.; Hikim, A.P.S. Mitochondria-Dependent Pathway Is Involved in Heat-Induced Male Germ Cell Death: Lessons from Mutant Mice. Biol. Reprod. 2004, 70, 1534–1540. [Google Scholar] [CrossRef]
- Santos, J.; Marques, C.; Baptista, M.C.; Pimenta, J.; Teixeira, J.; Montezinho, L.; Cagide, F.; Borges, F.; Oliveira, P.J.; Pereira, R.M.L. Effect of a novel hydroxybenzoic acid-based mitochondria directed antioxidant molecule on bovine sperm function and embryo production. Animals 2022, 12, 804. [Google Scholar] [CrossRef]
- Hikim, A.P.S.; Lue, Y.; Diaz-Romero, M.; Yen, P.H.; Wang, C.; Swerdloff, R.S. Deciphering the pathways of germ cell apoptosis in the testis. J. Steroid Biochem. Mol. Biol. 2003, 85, 175–182. [Google Scholar] [CrossRef]
- Hikim, A.P.S.; Lue, Y.; Yamamoto, C.M.; Vera, Y.; Rodriguez, S.; Yen, P.H.; Soeng, K.; Wang, C.; Swerdloff, R.S. Key Apoptotic Pathways for Heat-Induced Programmed Germ Cell Death in the Testis. Endocrinology 2003, 144, 3167–3175. [Google Scholar] [CrossRef] [PubMed]
- Green, D.R. Apoptotic Pathways: Paper Wraps Stone Blunts Scissors. Cell 2000, 102, 1–4. [Google Scholar] [CrossRef]
- Guo, X.; Chi, S.; Cong, X.; Li, H.; Jiang, Z.; Cao, R.; Tian, W. Baicalin protects sertoli cells from heat stress-induced apoptosis via activation of the Fas/FasL pathway and Hsp72 expression. Reprod. Toxicol. 2015, 57, 196–203. [Google Scholar] [CrossRef]
- Wang, M.; Su, P. The role of the Fas/FasL signaling pathway in environmental toxicant-induced testicular cell apoptosis: An update. Syst. Biol. Reprod. Med. 2018, 64, 93–102. [Google Scholar] [CrossRef]
- Bălan, I.; Nicolae, R.; Vladimir, B.; Nicolae, F.; Alexandru, D.; Irina, B.; Roman, C. The biological role of apoptosis in spermatogenesis: A review. Numărul 2020, 2, 137–145. [Google Scholar]
- Xavier, M.J.; Nixon, B.; Roman, S.D.; Aitken, R.J. Improved methods of DNA extraction from human spermatozoa that mitigate experimentally-induced oxidative DNA damage. PLoS ONE 2018, 13, e0195003. [Google Scholar] [CrossRef]
- Heninger, N.L.; Staub, C.; Blanchard, T.; Johnson, L.; Varner, D.D.; Forrest, D. Germ cell apoptosis in the testes of normal stallions. Theriogenology 2004, 62, 283–297. [Google Scholar] [CrossRef]
- Rockett, J.C.; Mapp, F.L.; Garges, J.B.; Luft, J.C.; Mori, C.; Dix, D.J. Effects of Hyperthermia on Spermatogenesis, Apoptosis, Gene Expression, and Fertility in Adult Male Mice. Biol. Reprod. 2001, 65, 229–239. [Google Scholar] [CrossRef]
- Sharma, R.; Iovine, C.; Agarwal, A.; Henkel, R. Tunel assay- Standardized method for testing sperm DNA fragmentation. Andrologia 2021, 53, e13738. [Google Scholar] [CrossRef]
- Peña, F.J.; O’Flaherty, C.; Ortiz Rodríguez, J.M.; Cano, F.E.M.; Gaitskell-Phillips, G.L.; Gil, M.C.; Ferrusola, C.O. Redox Regulation and Oxidative Stress: The Particular Case of the Stallion Spermatozoa. Antioxidants 2019, 8, 567. [Google Scholar] [CrossRef] [PubMed]
- Evenson, D.; Jost, L. Sperm chromatin structure assay is useful for fertility assessment. Methods Cell Sci. 2000, 22, 169–189. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Setchell, B.P. Effects of paternal heat stress on the in vivo development of preimplantation embryos in the mouse. Reprod. Nutr. Dev. 2004, 44, 617–629. [Google Scholar] [CrossRef][Green Version]
- Rahman, M.B.; Kamal, M.M.; Rijsselaere, T.; Vandaele, L.; Shamsuddin, M.; Van Soom, A. Altered chromatin condensation of heat-stressed spermatozoa perturbs the dynamics of DNA methylation reprogramming in the paternal genome after in vitro fertilisation in cattle. Reprod. Fertil. Dev. 2014, 26, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Kiefer, H.; Sellem, E.; Bonnet-Garnier, A.; Pannetier, M.; Costes, V.; Schibler, L.; Jammes, H. The epigenome of male germ cells and the programming of phenotypes in cattle. Anim. Front. 2021, 11, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Drevet, J.R. The Importance of Oxidative Stress in Determining the Functionality of Mammalian Spermatozoa: A Two-Edged Sword. Antioxidants 2020, 9, 111. [Google Scholar] [CrossRef]
- Rivlin, J.; Mendel, J.; Rubinstein, S.; Etkovitz, N.; Breitbart, H. Role of hydrogen peroxide in sperm capacitation and acrosome reaction. Biol. Reprod. 2004, 70, 518–522. [Google Scholar] [CrossRef]
- Koppers, A.J.; Reddy, T.; O’Bryan, M.K. The role of cysteine-rich secretory proteins in male fertility. Asian J. 2011, 13, 111–117. [Google Scholar] [CrossRef]
- Balić, I.M.; Milinković-Tur, S.; Samardžija, M.; Vince, S. Effect of age and environmental factors on semen quality, glutathione peroxidase activity and oxidative parameters in Simmental bulls. Theriogenology 2012, 78, 423–431. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, W.; Yu, H.; Xu, L.; Qu, M.; Li, Y. Improved antioxidant activity and rumen fermentation in beef cattle under heat stress by dietary supplementation with creatine pyruvate. J. Anim. Sci. 2020, 91, e13486. [Google Scholar] [CrossRef]
- Colombo, E.A.; Cooke, R.F.; Millican, A.A.; Schubach, K.M.; Scatolin, G.N.; Rett, B.; Brandão, A.P. Supplementing an immunomodulatory feed ingredient to improve thermoregulation and performance of finishing beef cattle under heat stress conditions. J. Anim. Sci. 2019, 97, 4085–4092. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capela, L.; Leites, I.; Romão, R.; Lopes-da-Costa, L.; Pereira, R.M.L.N. Impact of Heat Stress on Bovine Sperm Quality and Competence. Animals 2022, 12, 975. https://doi.org/10.3390/ani12080975
Capela L, Leites I, Romão R, Lopes-da-Costa L, Pereira RMLN. Impact of Heat Stress on Bovine Sperm Quality and Competence. Animals. 2022; 12(8):975. https://doi.org/10.3390/ani12080975
Chicago/Turabian StyleCapela, Luís, Inês Leites, Ricardo Romão, Luís Lopes-da-Costa, and Rosa M. Lino Neto Pereira. 2022. "Impact of Heat Stress on Bovine Sperm Quality and Competence" Animals 12, no. 8: 975. https://doi.org/10.3390/ani12080975
APA StyleCapela, L., Leites, I., Romão, R., Lopes-da-Costa, L., & Pereira, R. M. L. N. (2022). Impact of Heat Stress on Bovine Sperm Quality and Competence. Animals, 12(8), 975. https://doi.org/10.3390/ani12080975