
Does Age, Residency, or Feeding Guild Coupled with a Drought Index Predict Avian Health during Fall Migration?


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Site Location
2.2. Data Collection
2.3. Statistical Analyses
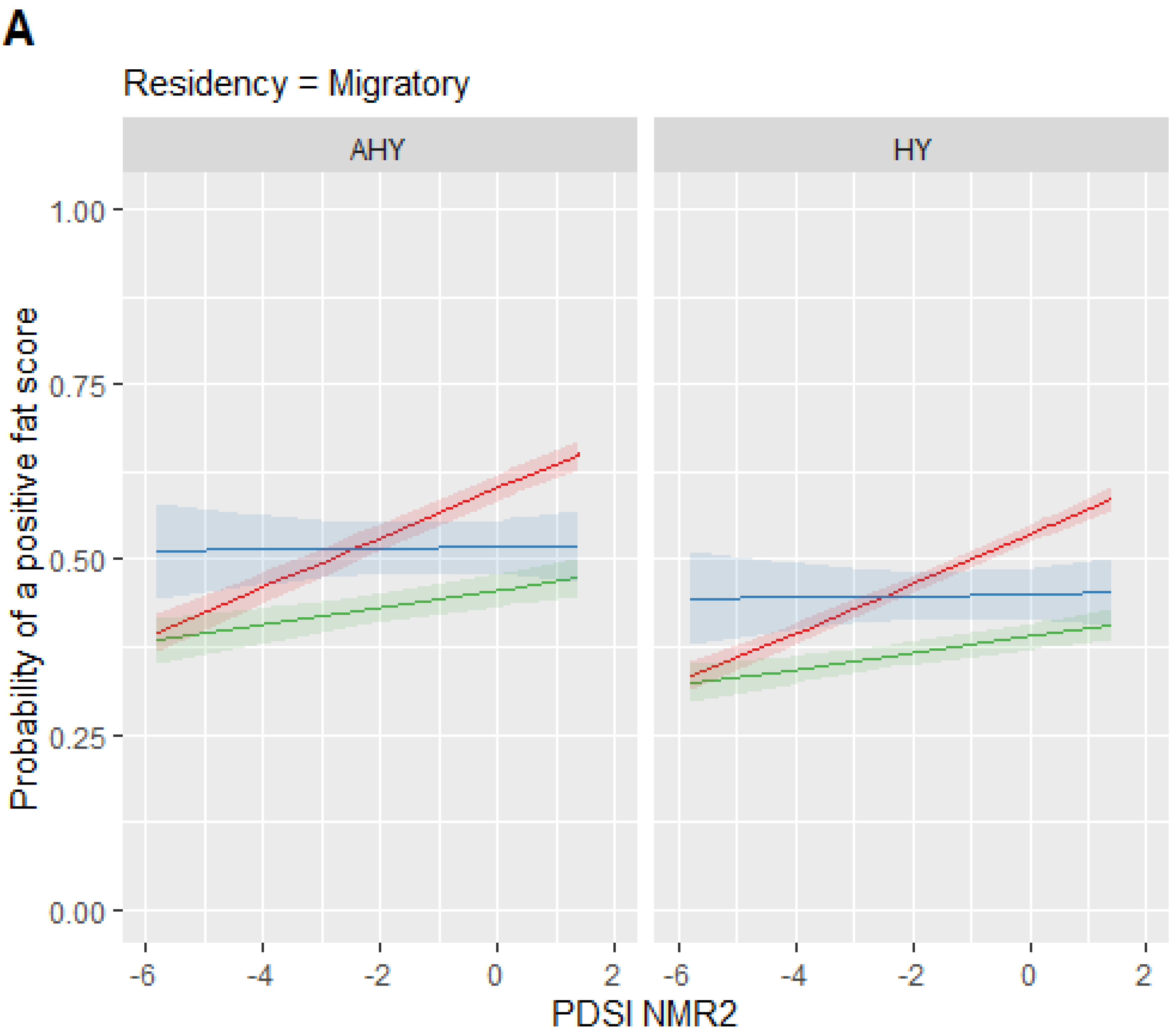
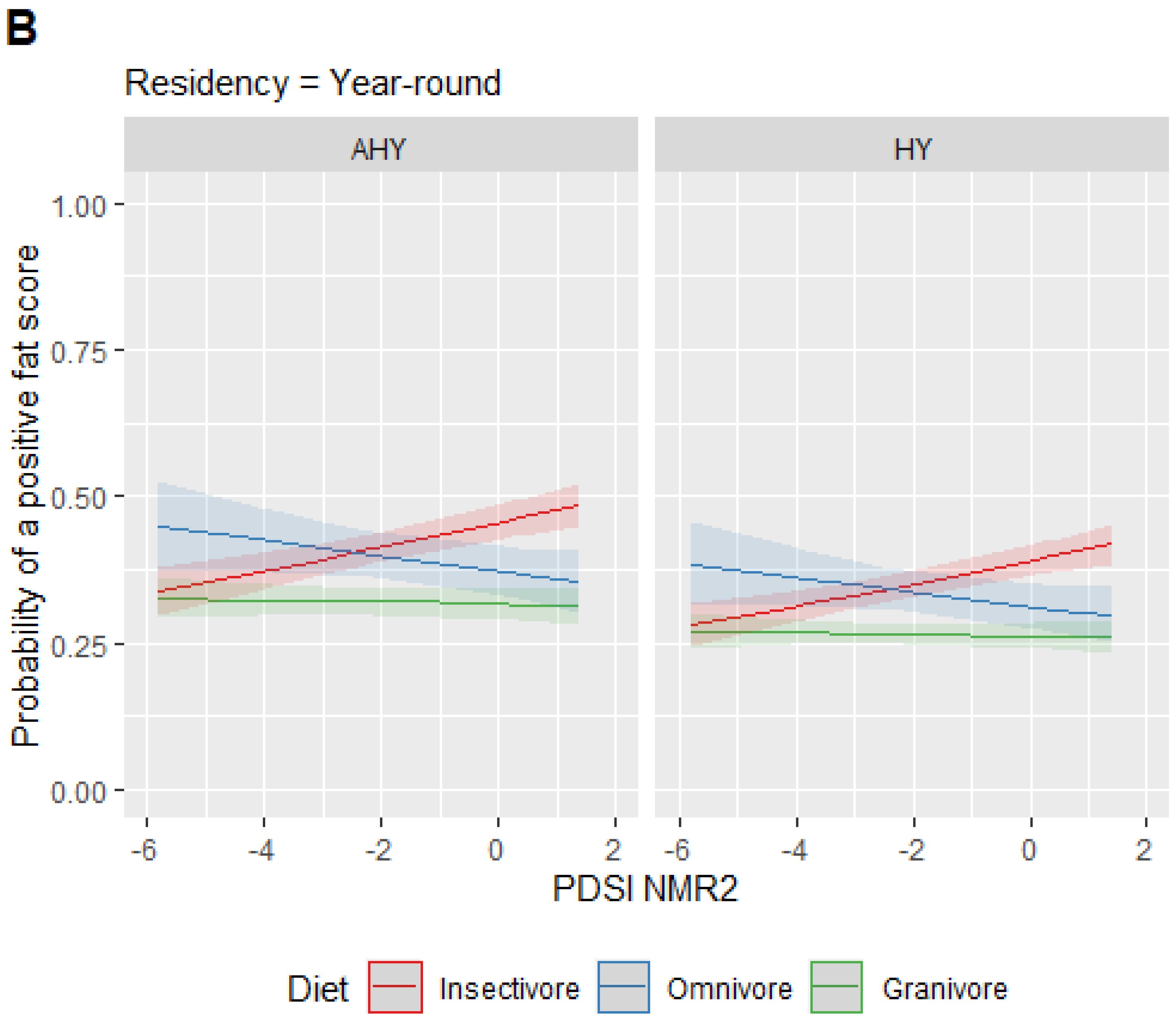
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ziolkowski, D., Jr.; Pardieck, K.; Sauer, J.R. On the road again for a bird survey that counts. Banding 2010, 42, 32–39. [Google Scholar]
- DeSante, D.F.; Burton, K.M.; Kaschube, D.R.; Velez, P.; Froehlich, D.; Albert, S. MAPS Manual: 2021 Protocol; The Institute for Bird Populations: Petaluma, CA, USA, 2021. [Google Scholar]
- DeSante, D.F.; Kaschube, D.R.; Saracco, J.F. Vital Rates of North American Landbirds. 2015. Available online: https://www.vitalratesofnorthamericanlandbirds.org (accessed on 15 December 2021).
- Ellwood, E.R.; Gallinat, A.; Primack, R.B.; Loyd-Evans, T.L. Autumn migration of North American landbirds. In Phenological Synchrony and Bird Migration: Changing Climate and Seasonal Resources in North America. Studies in Avian Biology (No. 47); Wood, E.M., Kellermann, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 193–205. [Google Scholar]
- McWilliams, S.R.; Guglielmo, C.; Pierce, B.; Klaassen, M. Flying, fasting, and feeding in birds during migration: A nutritional and physiological ecology perspective. J. Avian Biol. 2004, 35, 377–393. [Google Scholar] [CrossRef] [Green Version]
- Klaassen, R.H.G.; Hake, M.; Strandberg, R.; Koks, B.J.; Trierweiler, C.; Exo, K.; Bairlein, F.; Alerstam, T. When and where does mortality occur in migratory birds? Direct evidence from long-term satellite tracking of raptors. J. Anim. Ecol. 2014, 83, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Sergio, F.; Tavecchia, G.; Tanferna, A.; Blas, J.; Blanco, G.; Hiraldo, F. When and where mortality occurs throughout the annual cycle changes with age in a migratory bird: Individual versus population implications. Sci. Rep. 2019, 9, 17352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussell, D.J.T.; Ralph, C.J. Recommended methods for monitoring change in bird populations by counting and capture of migrants. N. Am. Bird Bander 2005, 30, 6–20. [Google Scholar]
- Kamm, M.D.; Lloyd-Evans, T.L.; Handmaker, M.; Reed, J.M. A half-century of changes in migratory landbird numbers along coastal Massachusetts. PLoS ONE 2019, 14, e0222232. [Google Scholar] [CrossRef] [PubMed]
- Mills, A.M. Changes in the timing of spring and autumn migration in North American migrant passerines during a period of global warming: Passerine migration and global warming. IBIS 2005, 147, 259–269. [Google Scholar] [CrossRef]
- Richardson, W.J. Timing and amount of bird migration in relation to weather: A review. Oikos 1978, 30, 224–272. [Google Scholar] [CrossRef]
- Jones, T.; Cresswell, W. The phenology mismatch hypothesis: Are declines of migrant birds linked to uneven global climate change? J. Anim. Ecol. 2010, 79, 98–108. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, E.A.; Macdonald, C.M.; Gilchrist, H.G.; Love, O.P. Spring and fall migration phenology of an Arctic-breeding passerine. J. Ornithol. 2016, 157, 681–693. [Google Scholar] [CrossRef]
- Rousseau, J.S.; Alexander, J.D.; Betts, M.G. Using continental-scale bird banding data to estimate demographic migratory patterns for Rufous Hummingbird (Selasphorus rufus). Avian Conserv. Ecol. 2020, 15, 2. [Google Scholar] [CrossRef]
- Deppe, J.L.; Ward, M.P.; Bolus, R.T.; Diehl, R.H.; Celis-Murillo, A.; Zenzal, T.J., Jr.; Moore, F.R.; Benson, T.J.; Smolinsky, J.A.; Schofield, L.N.; et al. Fat, weather, and date affect migratory songbirds’ departure decisions, routes, and time it takes to cross the Gulf of Mexico. Proc. Natl. Acad. Sci. USA 2015, 112, E6331–E6338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sjöberga, S.; Alerstama, T.; Åkessona, S.; Schulz, A.; Weidauer, A.; Coppack, T.; Muheima, R. Weather and fuel reserves determine departure and flight decisions in passerines migrating across the Baltic Sea. Anim. Behav. 2015, 104, 59–68. [Google Scholar] [CrossRef]
- Pilastro, A.; Magnani, A. Weather conditions and fat accumulation dynamics in pre-migratory roosting Barn Swallows Hirundo rustica. J. Avian Biol. 1997, 28, 338–344. [Google Scholar] [CrossRef]
- Cornelius, J.M.; Hahn, T.P.; Robart, A.R.; Vernasco, B.J.; Zahor, D.L.; Glynn, K.J.; Navis, C.J.; Watts, H.E. Seasonal patterns of fat deposits in relation to migratory strategy in facultative migrants. Front. Ecol. Evol. 2021, 9, 691808. [Google Scholar] [CrossRef]
- Van de Pol, M.; Ens, B.J.; Heg, D.; Brouwer, L.; Krol, J.; Maier, M.; Exo, K.; Oosterbeek, K.; Lok, T.; Eising, C.M.; et al. Do changes in the frequency, magnitude and timing of extreme climatic events threaten the population viability of coastal birds? J. Appl. Ecol. 2010, 47, 720–730. [Google Scholar] [CrossRef]
- Gimon, E. Lessons from the Big Texas Freeze. 2021. Available online: https://energyinnovation.org/wp-content/uploads/2021/05/Lessons-from-the-Texas-Big-Freeze.pdf (accessed on 15 December 2021).
- New Mexico Department of Game and Fish. Starvation, Unexpected Weather to Blame in Mass Migratory Songbird Mortality. 2020. Available online: https://content.govdelivery.com/accounts/NMDGF/bulletins/2afbc3e?reqfrom=share (accessed on 15 December 2021).
- Johnson, K. The Southwest Is Facing an ‘Unprecedented’ Migratory Bird Die-Off. 2020. Audubon Magazine. Available online: https://www.audubon.org/news/the-southwest-facing-unprecedented-migratory-bird-die (accessed on 15 December 2021).
- Yang, D.; Yang, A.; Yang, J.; Xu, R.; Qiu, H. Unprecedented migratory bird die-off: A citizen-based analysis on the spatiotemporal patterns of mass mortality events in the western United States. GeoHealth 2021, 5, e2021GH000395. [Google Scholar] [CrossRef] [PubMed]
- Waller, E.K.; Crimmins, T.M.; Walker, J.J.; Posthumus, E.E.; Weltzin, J.F. Differential changes in the onset of spring across US National Wildlife Refuges and North American migratory bird flyways. PLoS ONE 2018, 13, e0202495. [Google Scholar]
- Gustafson, M.E.; Hildenbrand, J.; Metras, L. The North American Bird Banding Manual. Version 1.0. 1997. Available online: https://www.pwrc.usgs.gov/bbl/manual/ (accessed on 15 December 2021).
- Pyle, P. Identification Guide to North American Passerines; Slate Creek Press: Bolinas, CA, USA, 1997; Volume 1. [Google Scholar]
- Liknes, E.T.; Swanson, D.L. Phenotypic flexibility of body composition associated with seasonal acclimatization in passerine birds. J. Therm. Biol. 2011, 36, 363–370. [Google Scholar] [CrossRef]
- Benson, A.; Winker, K. Fat-deposition strategies among high-latitude passerine migrants. Auk 2005, 122, 544–557. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference—A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Belsley, D.A.; Kuh, E.; Welsch, R.E. Regression Diagnostics: Identifying Influential Data and Sources of Collinearity; John Wiley and Sons: New York, NY, USA, 1980. [Google Scholar]
- Ott, R.L.; Longnecker, M. An Introduction to Statistical Methods and Data Analysis; Duxbury Press: Belmont, CA, USA, 2001. [Google Scholar]
- Birds of North America; Rodewald, P. (Ed.) Cornell Laboratory of Ornithology: Ithaca, NY, USA; Available online: https://birdsna.org (accessed on 15 December 2021).
- The Climate Data Guide: Palmer Drought Severity Index (PDSI); Dai, A., National Center for Atmospheric Research Staff. Available online: https://climatedataguide.ucar.edu/climate-data/palmer-drought-severity-index-pdsi (accessed on 15 December 2021).
- National Centers for Environmental Information, Climate at a Glance: Global Mapping. Available online: https://www.ncdc.noaa.gov/cag/ (accessed on 10 February 2022).
- Rosenberg, K.V.; Dokter, A.M.; Blancher, P.J.; Sauer, J.R.; Smith, A.C.; Smith, P.A.; Stanton, J.C.; Panjabi, A.; Helft, L.; Parr, M.; et al. Decline of the North American avifauna. Science 2019, 366, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Pearse, I.S.; Koenig, W.K.; Kelly, D. Mechanisms of mast seeding: Resources, weather, cues, and selection. New Phytol. 2016, 212, 546–562. [Google Scholar] [CrossRef] [PubMed]
- Parmenter, R.R.; Zlotin, R.I.; Moore, D.I.; Myers, O.B. Environmental and endogenous drivers of tree mast production and synchrony in piñon–juniper–oak woodlands of New Mexico. Ecosphere 2018, 9, e02360. [Google Scholar] [CrossRef] [Green Version]
- McCullough, J. The Data behind Mysterious Bird Deaths in New Mexico. 2020. American Birding Association, North American Birds Field Ornithology. Available online: https://www.aba.org/the-data-behind-mysterious-bird-deaths-in-new-mexico/ (accessed on 15 December 2021).
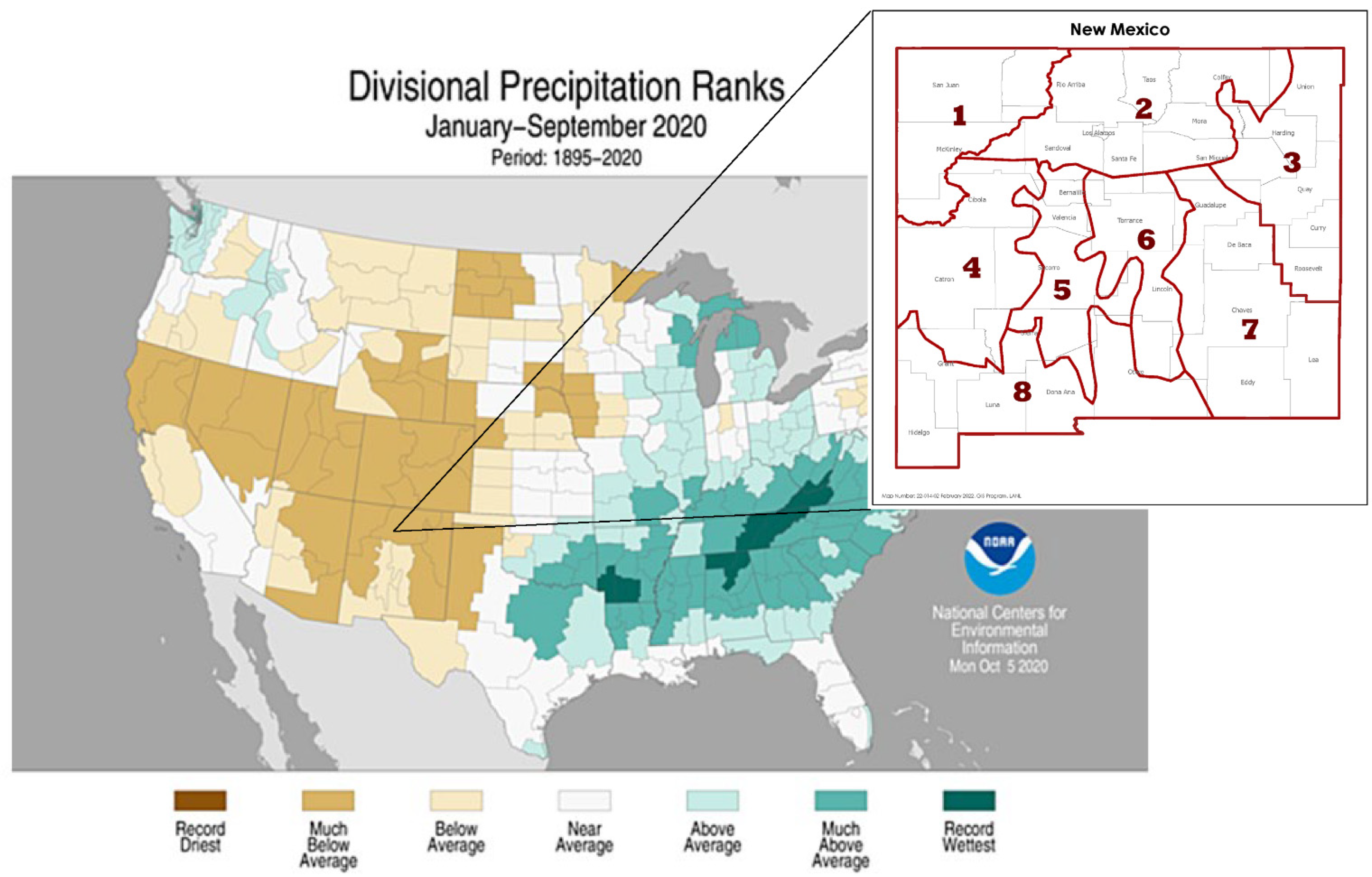


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name | Species | N | Mortality Event | Diet | Residency |
|---|---|---|---|---|---|
| Chipping Sparrow | Spizella passerina | 1892 | X | G | M |
| Dark-eyed Junco | Junco hyemalis | 1719 | X | G | M |
| Yellow-rumped Warbler | Setophaga coronata | 1390 | X | I | M |
| Wilson’s Warbler | Cardellina pusilla | 1181 | X | I | M |
| Pine Siskin | Spinus pinus | 1036 | X | G | YR |
| Ruby-crowned Kinglet | Corthylio calendula | 907 | X | I | M |
| Orange-crowned Warbler | Vermivora celata | 659 | X | I | M |
| Dusky Flycatcher | Empidonax oberholseri | 569 | X | I | M |
| Western Bluebird | Sialia mexicana | 504 | X | I | YR |
| House Wren | Troglodytes aedon | 499 | X | I | M |
| Lesser Goldfinch | Spinus psaltria | 468 | X | G | YR |
| Hermit Thrush | Catharus guttatus | 353 | X | I | M |
| Lincoln’s Sparrow | Melospiza lincolnii | 345 | X | O | M |
| Virginia’s Warbler | Leiothlypis virginiae | 289 | I | M | |
| Hammond’s Flycatcher | Empidonax hammondii | 282 | I | M | |
| MacGillivray’s Warbler | Geothlypis tolmiei | 245 | X | I | M |
| White-crowned Sparrow | Zonotrichia leucophrys | 242 | X | O | M |
| Western Wood-Pewee | Contopus sordidulus | 228 | X | I | M |
| Warbling Vireo | Vireo gilvus | 214 | I | M | |
| American Robin | Turdus migratorius | 177 | X | I | M |
| Western Tanager | Piranga ludoviciana | 165 | X | I | M |
| Green-tailed Towhee | Pipilo chlorurus | 146 | X | O | M |
| Bushtit | Psaltriparus minimus | 135 | X | I | YR |
| Mountain Chickadee | Poecile gambeli | 130 | X | I | YR |
| House Finch | Haemorhous mexicanus | 131 | X | G | YR |
| Spotted Towhee | Pipilo maculatus | 114 | X | O | YR |
| Townsend’s Warbler | Setophaga townsendi | 87 | X | I | M |
| Brewer’s Sparrow | Spizella breweri | 78 | X | I | M |
| Cordilleran Flycatcher | Empidonax occidentalis | 78 | X | I | M |
| Northern Flicker | Colaptes auratus | 74 | X | I | YR |
| Pygmy Nuthatch | Sitta pygmaea | 73 | X | I | YR |
| White-breasted Nuthatch | Sitta carolinensis | 62 | I | YR | |
| Williamson’s Sapsucker | Sphyrapicus thyroideus | 54 | X | I | YR |
| Yellow Warbler | Setophaga petechia | 50 | X | I | M |
| Model | K a | AIC b | ΔAIC c | wi |
|---|---|---|---|---|
| PDSI × Diet + PDSI × Resident + Age ‡ | 9 | 20208.64 | 0.00 | 1.000000 |
| PDSI + Resident + Age + Diet | 6 | 20280.51 | 71.87 | 0.000000 |
| PDSI + Resident + Diet | 5 | 20320.66 | 112.02 | 0.000000 |
| PDSI × Diet + Age | 7 | 20343.18 | 134.54 | 0.000000 |
| PDSI + Age + Diet | 5 | 20394.89 | 186.25 | 0.000000 |
| PDSI x Resident + Age | 5 | 20408.49 | 199.85 | 0.000000 |
| PDSI + Resident + Age | 4 | 20423.72 | 215.07 | 0.000000 |
| PDSI + Diet | 4 | 20424.61 | 215.96 | 0.000000 |
| PDSI + Resident | 3 | 20456.59 | 247.94 | 0.000000 |
| Age + Diet + Resident | 5 | 20460.35 | 251.71 | 0.000000 |
| Diet + Resident | 4 | 20508.08 | 299.44 | 0.000000 |
| PDSI + Age | 3 | 20601.37 | 392.73 | 0.000000 |
| PDSI | 2 | 20621.07 | 412.42 | 0.000000 |
| Diet + Resident | 3 | 20630.92 | 422.28 | 0.000000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stanek, J.E.; Thompson, B.E.; Milligan, S.E.; Tranquillo, K.A.; Fettig, S.M.; Hathcock, C.D. Does Age, Residency, or Feeding Guild Coupled with a Drought Index Predict Avian Health during Fall Migration? Animals 2022, 12, 454. https://doi.org/10.3390/ani12040454
Stanek JE, Thompson BE, Milligan SE, Tranquillo KA, Fettig SM, Hathcock CD. Does Age, Residency, or Feeding Guild Coupled with a Drought Index Predict Avian Health during Fall Migration? Animals. 2022; 12(4):454. https://doi.org/10.3390/ani12040454
Chicago/Turabian StyleStanek, Jenna E., Brent E. Thompson, Sarah E. Milligan, Keegan A. Tranquillo, Stephen M. Fettig, and Charles D. Hathcock. 2022. "Does Age, Residency, or Feeding Guild Coupled with a Drought Index Predict Avian Health during Fall Migration?" Animals 12, no. 4: 454. https://doi.org/10.3390/ani12040454
APA StyleStanek, J. E., Thompson, B. E., Milligan, S. E., Tranquillo, K. A., Fettig, S. M., & Hathcock, C. D. (2022). Does Age, Residency, or Feeding Guild Coupled with a Drought Index Predict Avian Health during Fall Migration? Animals, 12(4), 454. https://doi.org/10.3390/ani12040454