
Heterologous Challenge with PRRSV-1 MLV in Pregnant Vaccinated Gilts: Potential Risk on Health and Immunity of Piglets

 ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Description of the Farms
2.3. Farms’ History
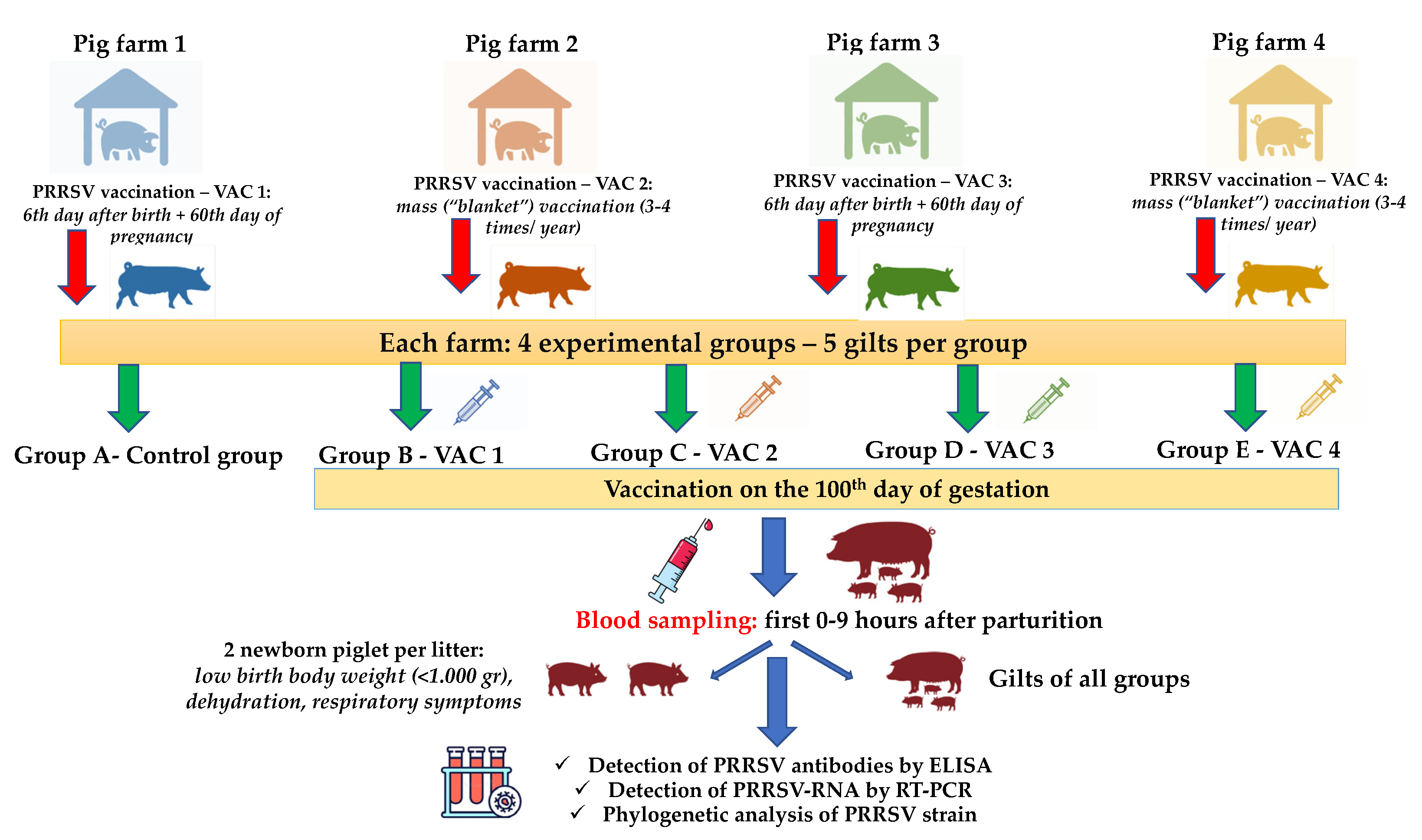
2.4. Experimental Design/Sample Collection/Records
2.5. Laboratory Examinations
2.5.1. Detection of PRRSV-RNA by RT-PCR and ORF5 Sequencing
2.5.2. Detection of PRRSV Antibodies by ELISA
2.6. Statistical Analysis
3. Results
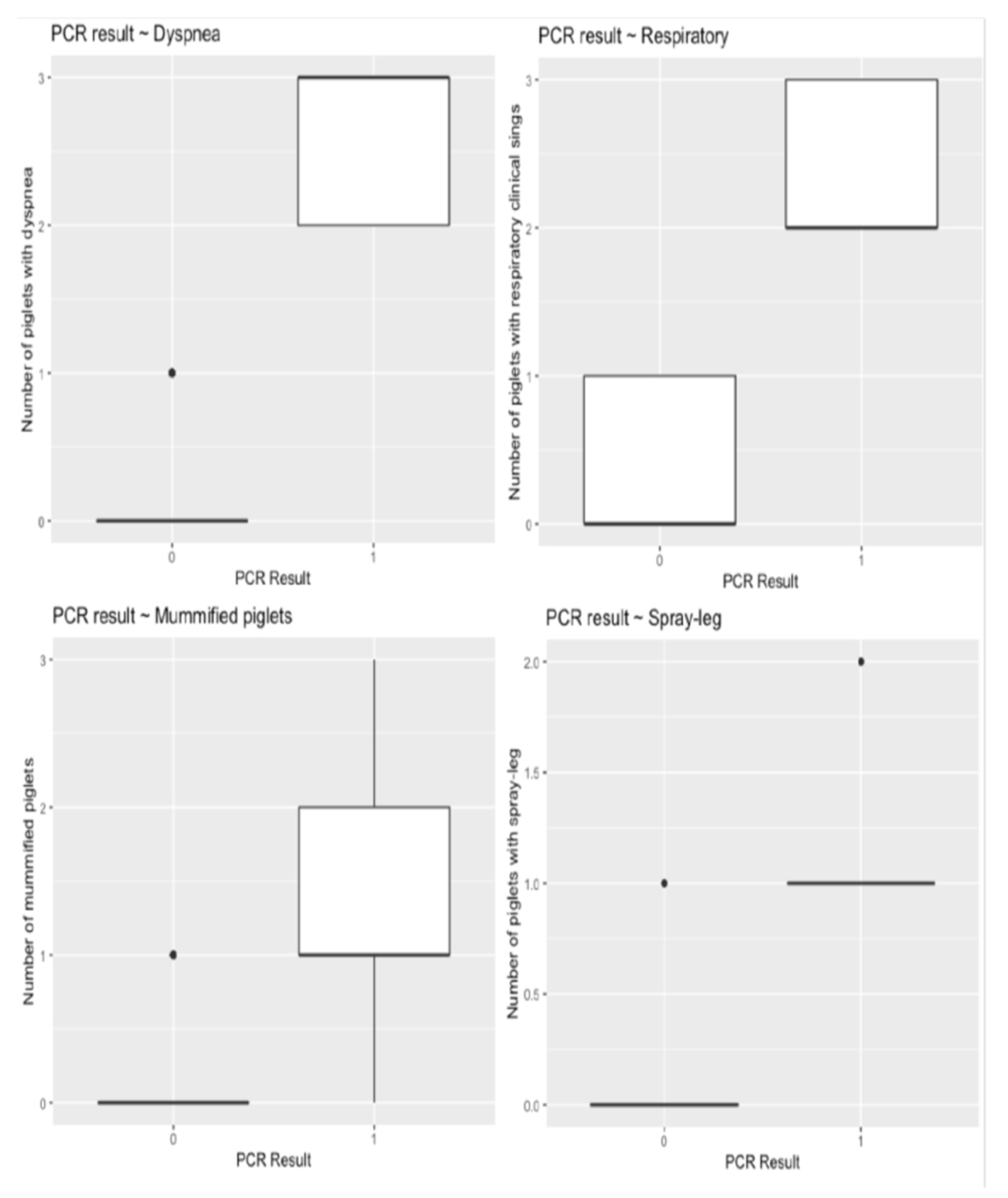
3.1. Clinical Evaluation and Reproductive Performance
3.2. Laboratory Results
3.2.1. Detection of PRRSV-RNA in Gilts and Piglets
3.2.2. Phylogenetic Analysis of PRRSV Strain
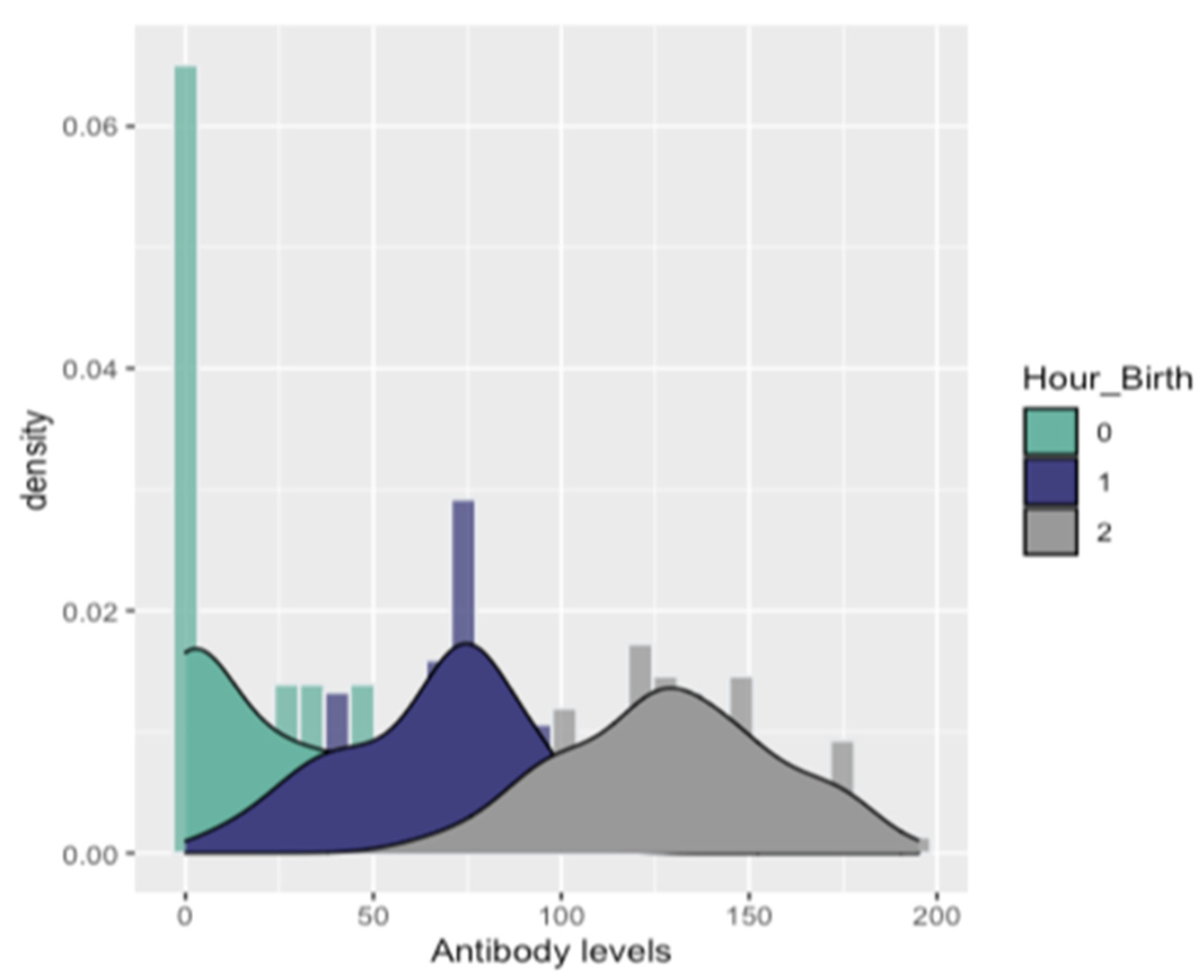
3.2.3. Detection of PRRSV Antibodies in Gilts and Piglets
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mang, S.; Tommy, T.Y.L.; Chung-Chau, H.; Raymond, K.H.H.; Faaberg, K.S.; Wennblom, T.; Murtaugh, M.P.; Stadejek, T.; Leung, F.C.C. Molecular epidemiology of PRRSV: A phylogenetic perspective. Virus Res. 2010, 154, 7–17. [Google Scholar]
- Cavanagh, D. Nidovirales: A new order comprising Coronaviridae and Arterividae. Arch. Virol. 1997, 142, 29–633. [Google Scholar]
- Papatsiros, V.G. Porcine Herd Health Management Practices for the Control of PRRSV Infection. In A Bird’s-Eye View of Veterinary Medicine; Perez-Marin, C.C., Ed.; In-Tech: Rijeka, Croatia, 2012. [Google Scholar]
- Stadler, J.; Zoels, S.; Eddicks, M.; Kraft, C.; Ritzmann, M.; Ladinig, A. Assessment of safety and reproductive performance after vaccination with a modified live-virus PRRS genotype 1 vaccine in pregnant sows at various stages of gestation. Vaccine 2016, 34, 3862–3866. [Google Scholar] [CrossRef] [PubMed]
- Murtaugh, M.P.; Elam, M.R.; Kakach, L.T. Comparison of the structural protein coding sequences of the VR-2332 and Lelystad virus strains of the PRRS virus. Arch. Virol. 1995, 140, 1451–1460. [Google Scholar] [CrossRef]
- Kappes, M.A.; Faaberg, K.S. PRRSV structure, replication and recombination: Origin of phenotype and genotype diversity. Virology 2015, 479–480, 475–486. [Google Scholar] [CrossRef] [Green Version]
- Benfield, D.A.; Collins, J.E.; Dee, S.A.; Halbur, P.G.; Joo, H.S.; Lager, K.M.; Mengeling, W.L.; Murtaugh, M.P.; Rossow, K.D.; Stevenson, G.W.; et al. Porcine reproductive and respiratory syndrome. In Diseases of Swine, 8th ed.; Straw, B.E., DAllaire, S., Mengeling, W.L., Taylor, D.J., Eds.; Iowa State University Press: Ames, IA, USA, 1999; pp. 201–232. [Google Scholar]
- Feng, D.; Nagy, J.A.; Pyne, K.; Dvorak, H.F.; Dvorak, A.M. In utero infection by porcine reproductive and respiratory syndrome virus is sufficient to increase susceptibility of piglets to challenge with Streptococcus suis type II. J. Virol. 2001, 75, 4889–4895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowland, R.R. The interaction between PRRSV and the late gestation pig fetus. Virus Res. 2010, 154, 114–222. [Google Scholar] [CrossRef] [PubMed]
- Pileri, E.; Mateu, E. Review on the transmission porcine reproductive and respiratory syndrome virus between pigs and farms and impact on vaccination. Vet. Res. 2016, 47, 108. [Google Scholar] [CrossRef] [Green Version]
- Holtkamp, D.; Kliebenstein, J.B.; Neumann, E.J.; Zimmerman, J.J.; Rotto, H.F.; Yoder, T.K.; Wang, C.; Yeske, P.E.; Mowrer, C.L.; Haley, C. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on U.S. pork producers. J. Swine Health Prod. 2013, 21, 72–84. [Google Scholar]
- Renke, C.; Nathues, C.; Swam, H.; Fiebig, K.; Weiss, C.; Eddicks, M.; Ritzmann, M.; Nathues, H. Application of an economic calculator to determine the cost of porcine reproductive and respiratory sundrome at farm- level in 21 pig herds in Germany. Porc. Health Manag. 2021, 7, 3. [Google Scholar] [CrossRef]
- Murtaugh, M.P.; Genzow, M. Immunological solutions for treatment and prevention of porcine reproductive and respiratory syndrome (PRRS). Vaccine 2011, 29, 8192–8204. [Google Scholar] [CrossRef]
- Papatsiros, V.G. Porcine Respiratory and Reproductive Syndrome Virus vaccinology: A review for commercial vaccines. Am. J. Anim. Vet. Sci. 2012, 7, 149–158. [Google Scholar]
- Lunney, J.K.; Fang, Y.; Ladinig, A.; Chen, N.; Li, Y.; Rowland, B. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV): Pathogenesis and Interaction with the Immune System. Annu. Rev. Anim. Biosci. 2016, 4, 129–154. [Google Scholar] [CrossRef] [PubMed]
- Scortti, M.; Prieto, C.; Alvarez, E.; Simarro, I.; Castro, J.M. Effects of two commercial European modifiedlive vaccines against porcine reproductive and respiratory syndrome viruses in pregnant gilts. Vet. J. 2006, 172, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Vanhee, M.; Delputte, P.L.; Delrue, I.; Geldhof, M.F.; Nauwynck, H.J. Development of an experimental inactivated PRRSV vaccine that induces virus-neutralizing antibodies. Vet. Res. 2009, 40, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karniychuk, U.; Nauwynck, H. Pathogenesis and prevention of placental and transplacental porcine reproductive and respiratory syndrome virus infection. Vet. Res. 2013, 44, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labarque, G.; Van Gucht, S.; Van Reeth, K.; Nauwynck, H.; Pensaert, M. Respiratory tract protection upon challenge of pigs vaccinated with attenuated porcine reproductive and respiratory syndrome virus vaccines. Vet. Microbiol. 2003, 95, 187–197. [Google Scholar] [CrossRef] [Green Version]
- Scortti, M.; Prieto, C.; Alvarez, E.; Simarro, I.; Castro, J.M. Failure of an inactivated vaccine against porcine reproductive and respiratory syndrome to protect gilts against a heterologous challenge with PRRSV. Vet. Rec. 2007, 161, 809–813. [Google Scholar]
- Nilubol, D.; Platt, K.B.; Halbur, P.G.; Torremorell, M.; Harris, D.L. The effect of a killed porcine reproductive and respiratory syndrome virus (PRRSV) vaccine treatment on virus shedding in previously PRRSV infected pigs. Vet. Microbiol. 2004, 102, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Nan, Y.; Wu, C.; Gu, G.; Sun, W.; Zhang, Y.-J.; Zhou, E.-M. Improved Vaccine against PRRSV: Current Progress and Future Perspective. Front. Microbiol. 2017, 8, 1635. [Google Scholar] [CrossRef] [PubMed]
- Benfield, D.A.; Nelson, E.; Collins, J.E.; Harris, L.; Goyal, S.M.; Bobinson, D.; Christianson, W.T.; Morrison, R.B.; Gorcyca, D.; Chladek, D. Characterization of swine infertility and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332). J. Vet. Diagn. Investig. 1997, 4, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Cheon, D.S.; Chae, C. Polymerase chain reaction-based restriction fragment length polymorphism pattern of porcine reproductive and respiratory syndrome virus directly from lung tissues without virus isolation in Korea. J. Vet. Med. Sci. 2001, 63, 567–571. [Google Scholar] [CrossRef] [Green Version]
- Bierk, M.D.; Dee, S.A.; Rossow, K.D.; Otake, S.; Collins, J.E.; Molitor, T.W. Diagnostic investigation of chronic porcine reproductive and respiratory syndrome virus in a breeding herd of pigs. Vet. Res. 2001, 148, 687–690. [Google Scholar] [CrossRef]
- Jiang, P.; Chen, P.Y.; Dong, Y.Y.; Cai, J.L.; Cai, B.X.; Jiang, Z.H. Isolation and genome characterization of porcine reproductive and respiratory syndrome virus in P.R. China. J. Vet. Diagn. Investig. 2000, 12, 156–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stadejek, T.; Stankevicius, A.; Storgaard, T.; Oleksiewicz, M.B.; Belak, S.; Drew, T.W.; Pejsak, Z. Identification of radically different variants of porcine reproductive and respiratory syndrome virus in Eastern Europe: Towards a common ancestor for European and American viruses. J. Gen. Virol. 2002, 83, 1861–1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann, E.J.; Kliebenstein, J.B.; Johnson, C.D.; Mabry, J.W.; Bush, E.; Seitzinger, A.H.; Green, A.; Zimmerman, J.J. Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States. J. Am. Vet. Med. Assoc. 2005, 227, 385–392. [Google Scholar] [CrossRef] [Green Version]
- Prieto, C.; Castro, J.M. Porcine reproductive and respiratory syndrome virus infection in the boar: A review. Theriogenology 2005, 63, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mateusen, B.; Van Soom, A.; Maes, D.G.D.; Favoreel, H.; Nauwynck, H.J. Receptor-determined susceptibility of preimplantation embryos to pseudorabies virus and porcine reproductive and respiratory syndrome virus. Biol. Reprod. 2007, 76, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Van Gorp, H.; Van Breedam, W.; Delputte, P.L.; Nauwynck, H.J. Sialoadhesin and CD163 join forces during entry of the porcine reproductive and respiratory syndrome virus. J. Gen. Virol. 2008, 89, 2943–2953. [Google Scholar] [CrossRef]
- Karniychuk, U.; De Spiegelaere, W.; Nauwynck, H.J. Porcine reproductive and respiratory syndrome virus infection is associated with an increased number of Sn- positive and CD8- positive cells in the maternal- fetal interface. Virus Res. 2013, 176, 285–291. [Google Scholar] [CrossRef]
- Kimman, T.G.; Cornelissen, L.A.; Moormann, R.J.; Rebel, J.M.J.; Stockhofe-Zurwieden, N. Challenges for porcine reproductive and respiratory syndrome virus (PRRSV) vaccinology. Vaccine 2009, 27, 3704–3718. [Google Scholar] [CrossRef] [PubMed]
- Chae, C. Commercial PRRS Modified-Live Virus Vaccines. Vaccines 2021, 9, 185. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Cai, X.; Tan, M.; Sun, F.; Wang, Y.; Shang, Y.; Liu, S.; Xiao, Y. Serodiagnosis, targeting nonstructural protein 4, of porcine reproductive and respiratory syndrome virus. Arch. Virol. 2018, 163, 411–418. [Google Scholar] [CrossRef]
- Martínez, E.; Riera, P.; Sitjà, M.; Fang, Y.; Oliveira, S.; Maldonado, J. Simultaneous detection and genotyping of porcine reproductive and respiratory syndrome virus (PRRSV) by real-time RT-PCR and amplicon melting curve analysis using SYBR Green. Res. Vet. Sci. 2008, 85, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Mateu, E.; Martin, M.; Vidal, D. Genetic diversity and phylogenetic analysis of glycoprotein 5 of European-type porcine reproductive and respiratory virus strains in Spain. J. Gen. Virol. 2003, 84, 529–534. [Google Scholar] [CrossRef]
- Indik, S.; Valíček, L.; Klein, D.; Klánová, J. Variations in the major envelope glycoprotein GP5 of Czech strains of porcine reproductive and respiratory syndrome virus. J. Gen. Virol. 2000, 81, 2497–2502. [Google Scholar] [CrossRef]
- Lee, J.A.; Lee, N.H.; Lee, J.B.; Park, S.Y.; Song, C.S.; Choi, I.S.; Lee, S.W. Genetic diversity of the Korean field strains of porcine reproductive and respiratory syndrome virus. Infect. Genet. Evol. 2016, 40, 288–294. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [Green Version]
- Biernacka, K.; Podgorska, K.; Tyszka, A.; Stadejek, T. Comparison of six commercial ELISAs for the detection of antibodies against porcine reproductive and respiratory syndrome virus (PRRSV) in field serum samples. Vet. Sci. Res. 2018, 121, 40–45. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Montgomery, D.C.; Peck, E.A.; Vining, G.G. Introduction to Linear Regression Analysis; John Wiley & Sons: Hoboken, NJ, USA, 2021. [Google Scholar]
- Peltoniemi, O.; Björkman, S.; Maes, D. Reproduction of group-housed sows. Porc. Health Manag. 2016, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Rowland, R.R.; Lunney, J.; Dekkers, J. Control of porcine reproductive and respiratory syndrome (PRRS) through genetic improvements in disease resistance and tolerance. Front. Genet. 2012, 3, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, C.; Chae, C. Colocalization of porcine reproductive and respiratory syndrome virus and porcine circovirus 2 in porcine dermatitis and nephrology syndrome by double-labelling. Vet. Pathol. 2001, 38, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Madapong, A.; Temeeyasen, G.; Saeng-Chuto, K.; Tripipat, T.; Navasakuljinda, W.; Boonsoongern, A.; Tantituvanont, A.; Nilubol, D. Humoral immune responses and viral shedding following vaccination with modified live porcine reproductive and respiratory syndrome virus vaccines. Arch. Virol. 2017, 162, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Schelkopf, A.; Nerem, J.; Cowles, B.; Amodie, D.; Swalla, R.; Dee, S. Reproductive, productivity, and mortality outcomes in late-gestation gilts and their litters following simulation of inadvertent exposure to a modified-live vaccine strain of porcine reproductive and respiratory syndrome (PRRS) virus. Vaccine 2014, 32, 4639–4643. [Google Scholar] [CrossRef]
- Reynaud, G.; Charreyre, C.; Tedeschi, L. Homologus PRRSV challenge after vaccination with an inactivated PRRS vaccine: PROGRESSIS® Merial. In Proceedings of the 18th International Pig Veterinary Society Congress, Hamburg, Germany, 27 June–1 July 2004; Volume 1, p. 138. [Google Scholar]
- Eclercy, J.; Renson, P.; Hirchaud, E.; Andraud, M.; Beven, V.; Paboeuf, F.; Rose, N.; Blanchard, Y.; Bourry, O. Phenotypic and Genetic Evolutions of a Porcine Reproductive and Respiratory Syndrome Modified Live Vaccine after Limited Passages in Pigs. Vaccine 2021, 9, 392. [Google Scholar] [CrossRef]
- Navarro, P.; Christenson, R.K.; Weber, P.; Rothschild, M.; Ekhard, G.; Butler, J.E. IgA allotypes are not equally transcribed or expressed in heterozygous swine. Mol. Immunol. 2001, 37, 653. [Google Scholar] [CrossRef] [Green Version]
- Guzman-Bautista, E.R.; Garcia-Ruiz, C.E.; Gama-Espinosa, A.L.; Ramirez- Estudillo, C.; Rojas-Gomez, O.I.; Vega-Lopez, M.A. Effect of age and maternal antibodies on the systemic and mucosal immune response after neonatal immunization in a porcine model. Immunology 2013, 141, 609–616. [Google Scholar] [CrossRef]
- Bandrick, M.; Ariza-Nieto, C.; Baidoo, S.K.; Molitor, T.W. Colostral antibody-mediated and cell-mediated immunity contributes to innate and antigen-specific immunity in piglets. Dev. Comp. Immunol. 2014, 43, 114–120. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, B.; Hill, M.; Reynaud, L.; Hey, A.; Barrow, P. FcRn Expression on Placenta and Fetal Jejunum during Early, Mid-, and Late Gestation in Minipigs. Toxicol. Pathol. 2016, 44, 486–491. [Google Scholar] [CrossRef] [Green Version]
- Hsueh, F.C.; Wang, S.Y.; Lin, W.H.; Lin, C.F.; Tsai, C.Y.; Huang, C.W.; Sun, N.; Chiou, M.T.; Lin, C.N. Correlation of Neutralizing Antibodies (NAbs) between Sows and Piglets and Evaluation of Protectability Associated with Maternally Derived NAbs in Pigs against Circulating Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) under Field Conditions. Vaccines 2021, 9, 414. [Google Scholar] [CrossRef]
- Greenwood, J.; Howes, R.; Lightman, S. The blood-retinal barrier in experimental autoimmune uveoretinitis. Leukocyte interactions and functional damage. Lab. Investig. 1994, 70, 39–52. [Google Scholar] [PubMed]
- Feng, D.; Nagy, J.A.; Pyne, K.; Dvorak, H.F.; Dvorak, A.M. Neutrophils emigrate from venules by a transendothelial cell pathway in response to FMLP. J. Exp. Med. 1998, 187, 903–915. [Google Scholar] [CrossRef] [Green Version]
- Butler, J.E.; Sun, J.; Weber, P.; Ford, S.P.; Rehakova, Z.; Sinkora, J.; Lager, K. Antibody Repertoire Development in Fetal and Neonatal Piglets. IV. Switch Recombination, Primarily in Fetal Thymus, Occurs Independent of Environmental Antigen and Is Only Weakly Associated with Repertoire Diversification. J. Immunol. 2001, 167, 3239–3249. [Google Scholar] [CrossRef]
- Montaner-Tarbes, S.; del Portillo, H.A.; Montoya, M.; Fraile, L. Key Gaps in the Knowledge of the Porcine Respiratory Reproductive Syndrome Virus (PRRSV). Front. Vet. Sci. 2019, 6, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, I.J.; Joo, H.S.; Goyal, S.M.; Molitor, T.W. A modified serum neutralization test for the detection of antibody to porcine reproductive and respiratory syndrome virus in swine sera. J. Vet. Diagn. Investig. 1994, 6, 289–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renukaradhya, G.J.; Aleksejev, K.; Jung, K.; Fang, Y.; Saif, L.J. Porcine reproductive and respiratory syndrome virus-induced immunosuppression exacerbates the inflammatory response to porcine respiratory coronavirus. Viral Immunol. 2010, 23, 457–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmerman, J.; Yoon, K.J.; Pirtle, E.C.; Wills, R.W.; Sanderson, T.J.; McGinley, M.J. Studies of porcine reproductive and respiratory syndrome (PRRS) virus infection in avian species. Vet. Microbiol. 1997, 55, 329–336. [Google Scholar] [CrossRef]
- Halbur, P.G.; Bush, E. Update on abortion storms and sow mortality. Swine Health Prod. 1997, 5, 73. [Google Scholar]
- Brockmeier, S.L.; Halbur, P.G.; Thacker, E.L. Porcine respiratory disease complex. In Polymicrobial Disease; Brogden, K.A., Guthmiller, J.M., Eds.; ASM Press: Washington, DC, USA, 2002; pp. 231–258. [Google Scholar]
- Chang, C.C.; Yoon, K.J.; Zimmermann, J.J.; Harmon, K.M.; Dixon, P.M.; Dvorak, C.M.; Murtaugh, M.P. Evolution of porcine reproductive and respiratory syndrome virus during sequential passages in pigs. J. Virol. 2002, 76, 4750–4763. [Google Scholar] [CrossRef] [Green Version]
- Butler, J.E.; Lager, K.M.; Golde, W.; Faaberg, K.S.; Sinkora, M.; Loving, C.; Zhang, Y.I. Porcine reproductive and respiratory syndrome (PRRS): An immune dysregulatory pandemic. Immunol. Res. 2014, 59, 81–108. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Farms | Group | Vaccination on the 100th Day of Gestation |
|---|---|---|
| Farm 1 (Gilts were vaccinated with VAC1 180th + 210th day of age + 60th day of gestation) | A | None–Control group |
| B | Vaccine One (VAC1)-strain DV | |
| C | Vaccine Two (VAC2)-strain VP- 046 BIS | |
| D | Vaccine Three (VAC3)-strain 94881 | |
| E | Vaccine Four (VAC4)-strain 96V198 | |
| Farm 2 (Gilts were vaccinated with strain VAC4 180th + 210th day of age + 60th day of gestation) | A | None–Control group |
| B | Vaccine One (VAC1)-strain DV | |
| C | Vaccine Two (VAC2)-strain VP-046 BIS | |
| D | Vaccine Three (VAC3)-strain 94881 | |
| E | Vaccine Four (VAC4)-strain 96V198 | |
| Farm 3 (Gilts were vaccinated with VAC3 180th + 210th day of age + 60th day of gestation) | A | None–Control group |
| B | Vaccine One (VAC1)-strain DV | |
| C | Vaccine Two (VAC2)-strain VP-046 BIS | |
| D | Vaccine Three (VAC3)-strain 94881 | |
| E | Vaccine Four (VAC4)-strain 96V198 | |
| Farm 4 (Gilts were vaccinated with strain VAC2 180th + 210th day of age + 60th day of gestation) | A | Non–Control group |
| B | Vaccine One (VAC1)-strain DV | |
| C | Vaccine Two (VAC2)-strain VP-046 BIS | |
| D | Vaccine Three (VAC3)-strain 94881 | |
| E | Vaccine Four (VAC4)-strain 96V198 |
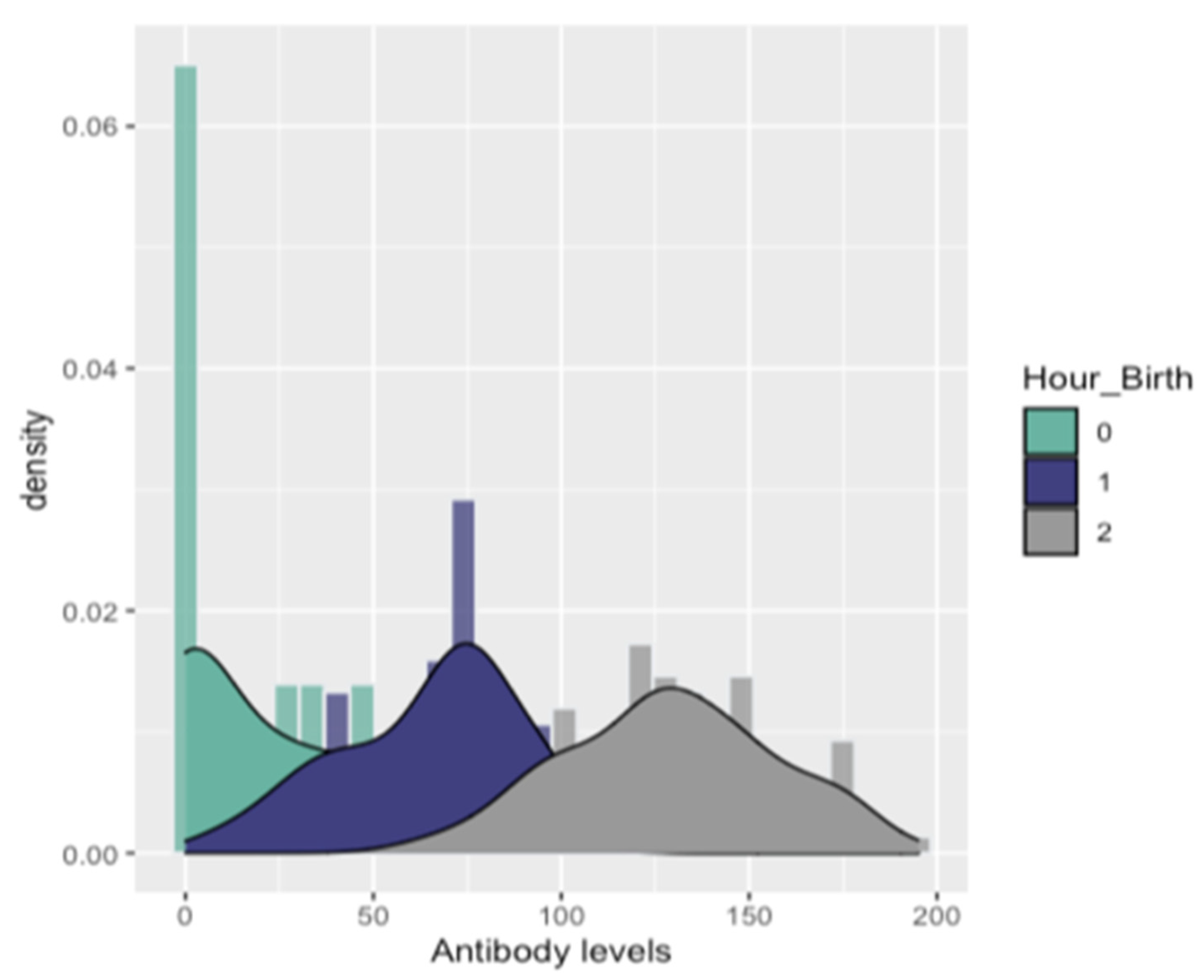
| Antibody Levels | 1st Quantile (25%) | Mean | Median | 3rd Quantile (75%) | RT-PCR Status | ||
|---|---|---|---|---|---|---|---|
| Positive | Negative | ||||||
| Hour_birth | 0–3 h | 0 | 14 | 23.7 | 45.5 | 12 | 20 |
| 3–6 h | 45.5 | 64.6 | 70.5 | 80.5 | 8 | 48 | |
| >6 h | 108 | 129 | 128 | 148 | 0 | 112 | |
| Experimental Farms | Group | DT (Days) | Number of Piglets Mean/Median (Min–Max) | |||||
|---|---|---|---|---|---|---|---|---|
| LBP | DBP | WBP | Mummies | SLP | RSP | |||
| Farm 1 | A | 118.4/119 (117–119) | 16/16 (14–18) | 1.4/1 (1–2) | 0.8/1 (0–1) | 0/0 (0–0) | 0/0 (0–0) | 0/0 (0–0) |
| B | 117.4/117 (116–119) | 13.4/13 (11–17) | 1.8/2 (1–3) | 1.6/1 (1–3) | 0.4/0 (0–1) | 0/0 (0–0) | 0.8/1 (0–1) | |
| C | 114.2/115 (112–116) | 9.8/10 (7–12) | 2.2/2 (2–3) | 3/3 (2–4) | 2/2 (1–3) | 1.2/1 (1–2) | 2.4/2 (2–3) | |
| D | 116.8/117 (116–118) | 8.8/10 (7–10) | 1.6/2 (1–2) | 1.6/2 (1–2) | 0.4/0 (0–1) | 0.4/0 (0–1) | 1/1 (1–1) | |
| E | 119.2/119 (118–120) | 15/15 (14–16) | 1.8/2 (1–3) | 2/2 (0–4) | 0/0 (0–0) | 0/0 (0–0) | 0.6/1 (0–1) | |
| Farm 2 | A | 117/117 (116–118) | 16.4/16 (16–17) | 1.8/2 (1–2) | 1.4/1 (0–3) | 0/0 (0–0) | 0/0 (0–0) | 1/1 (1–1) |
| B | 117/117 (116–118) | 8.2/8 (5–11) | 3.8/4 (3–5) | 1.4/1 (1–2) | 0/0 (0–0) | 0/0 (0–0) | 1/1 (1–1) | |
| C | 117/117 (116–118) | 12.4/12 (10–15) | 4.4/4 (2–7) | 2.4/2 (2–3) | 0.6/1 (0–1) | 0.4/0 (0–1) | 1/1 (1–1) | |
| D | 114.8/115 (113–117) | 5.6/6 (4–7) | 1.4/1 (1–2) | 2.4/2 (2–3) | 0.6/1 (0–1) | 1.2/1 (1–2) | 2.4/2 (2–3) | |
| E | 116.2/116 (115–118) | 14.6/14 (12–18) | 1.2/2 (0–2) | 2.4/2 (2–3) | 0/0 (0–0) | 0/0 (0–0) | 0/0 (0–0) | |
| Farm 3 | A | 116.6/117 (116–117) | 12.8/13 (12–14) | 1.6/2 (1–2) | 0/0 (0–0) | 0/0 (0–0) | 0/0 (0–0) | 0/0 (0–0) |
| B | 116.4/116 (116–117) | 13.4/13 (12–15) | 1/1 (0–2) | 0/0 (0–0) | 0/0 (0–0) | 0/0 (0–0) | 0/0 (0–0) | |
| C | 116.6/116 (115–119) | 13.4/13 (11–16) | 0.6/1 (0–1) | 0/0 (0–0) | 0/0 (0–0) | 0/0 (0–0) | 0/0 (0–0) | |
| D | 117/117 (116–118) | 12.2/12 (11–13) | 1.2/1 (1–2) | 0/0 (0–0) | 0.6/1 (0–1) | 0/0 (0–0) | 0/0 (0–0) | |
| E | 117/117 (116–118) | 14.4/14 (14–15) | 1/1 (0–2) | 0/0 (0–0) | 0/0 (0–0) | 0/0 (0–0) | 0/0 (0–0) | |
| Farm 4 | A | 117.4/117 (117–118) | 13/13 (11–15) | 1.6/2 (1–2) | 0.6/1 (0–1) | 0/0 (0–0) | 0/0 (0–0) | 0/0 (0–0) |
| B | 117.4/117 (117–118) | 11.6/12 (10–13) | 1.4/1 (1–2) | 0.6/1 (0–1) | 0/0 (0–0) | 0/0 (0–0) | 0.4/0 (0–1) | |
| C | 116.4/116 (116–117) | 11/11 (10–13) | 1.6/2 (1–2) | 0.6/1 (0–1) | 0/0 (0–0) | 0/0 (0–0) | 0.6/1 (0–1) | |
| D | 117.4/117 (117–118) | 11.8/12 (10–14) | 2.6/3 (2–3) | 1.4/1 (1–2) | 0/0 (0–0) | 0/0 (0–0) | 0.6/1 (0–1) | |
| E | 117/117 (116–118) | 14/14 (13–16) | 2.4/2 (2–3) | 1/1 (0–2) | 0/0 (0–0) | 0/0 (0–0) | 0.6/1 (0–1) | |
| Linear Regression | |||
|---|---|---|---|
| Parameter | Hour after birth | Estimate (95% CI) | p Value |
| Hour Birth | 0–3 h | 4.3 (2.75; 6.71) | <0.001 |
| 3–6 h | 8.26 (5.77; 11.8) | <0.001 | |
| 6–9 h | 26.33 (17.23; 40.24) | <0.001 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papakonstantinou, G.; Meletis, E.; Christodoulopoulos, G.; Tzika, E.D.; Kostoulas, P.; Papatsiros, V.G. Heterologous Challenge with PRRSV-1 MLV in Pregnant Vaccinated Gilts: Potential Risk on Health and Immunity of Piglets. Animals 2022, 12, 450. https://doi.org/10.3390/ani12040450
Papakonstantinou G, Meletis E, Christodoulopoulos G, Tzika ED, Kostoulas P, Papatsiros VG. Heterologous Challenge with PRRSV-1 MLV in Pregnant Vaccinated Gilts: Potential Risk on Health and Immunity of Piglets. Animals. 2022; 12(4):450. https://doi.org/10.3390/ani12040450
Chicago/Turabian StylePapakonstantinou, Georgios, Eleftherios Meletis, Georgios Christodoulopoulos, Eleni D. Tzika, Polychronis Kostoulas, and Vasileios G. Papatsiros. 2022. "Heterologous Challenge with PRRSV-1 MLV in Pregnant Vaccinated Gilts: Potential Risk on Health and Immunity of Piglets" Animals 12, no. 4: 450. https://doi.org/10.3390/ani12040450
APA StylePapakonstantinou, G., Meletis, E., Christodoulopoulos, G., Tzika, E. D., Kostoulas, P., & Papatsiros, V. G. (2022). Heterologous Challenge with PRRSV-1 MLV in Pregnant Vaccinated Gilts: Potential Risk on Health and Immunity of Piglets. Animals, 12(4), 450. https://doi.org/10.3390/ani12040450