
Causes of Death and Pathological Findings in Stranded Harbour Porpoises (Phocoena phocoena) from Swedish Waters

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Sample and Data Collection
2.3. Microscopic Examination
2.4. Bacteriology
2.5. Virology
2.6. Chemistry
2.7. Diagnoses
3. Results
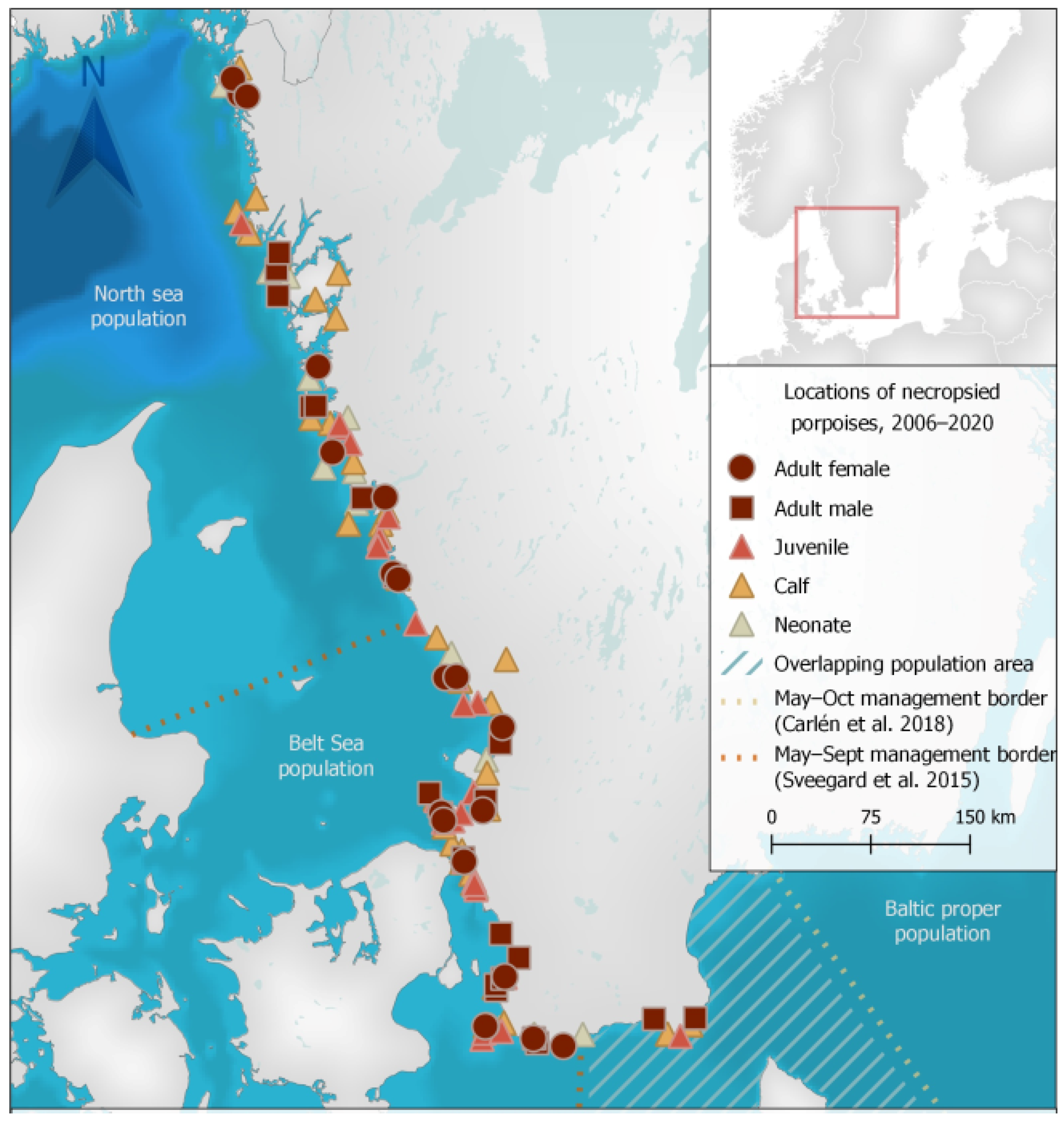
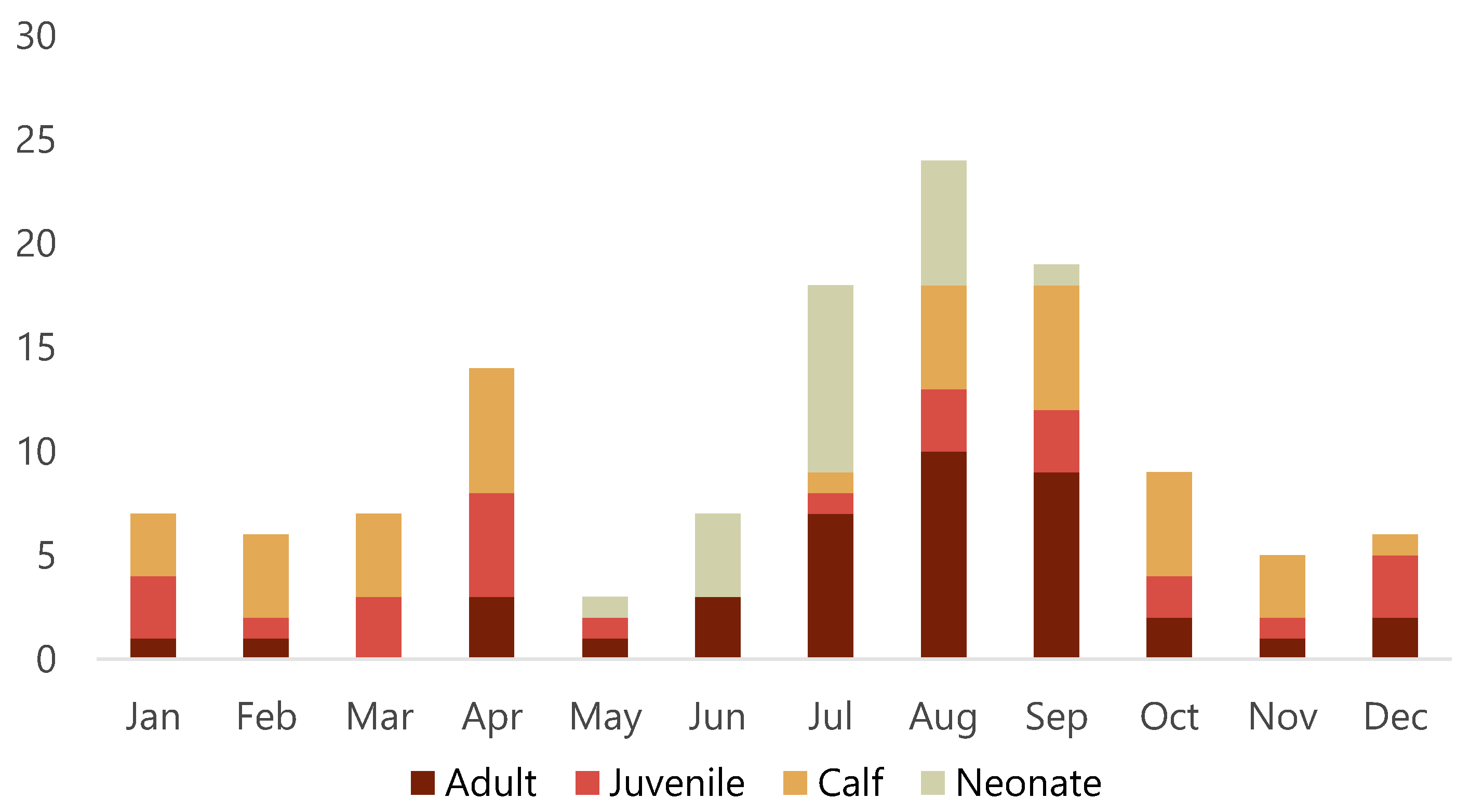
3.1. Animals
3.2. Ancillary Diagnostic Analyses
3.2.1. Bacteriology
3.2.2. Virology
3.3. Diagnoses and Causes of Death
3.3.1. Primary Diagnoses
3.3.2. Secondary Diagnoses
3.3.3. Parasitism
4. Discussion
4.1. Animals
4.2. Diagnoses, Disease, Other Threats
4.2.1. Bycatch and Probable Bycatch
4.2.2. Infectious Disease
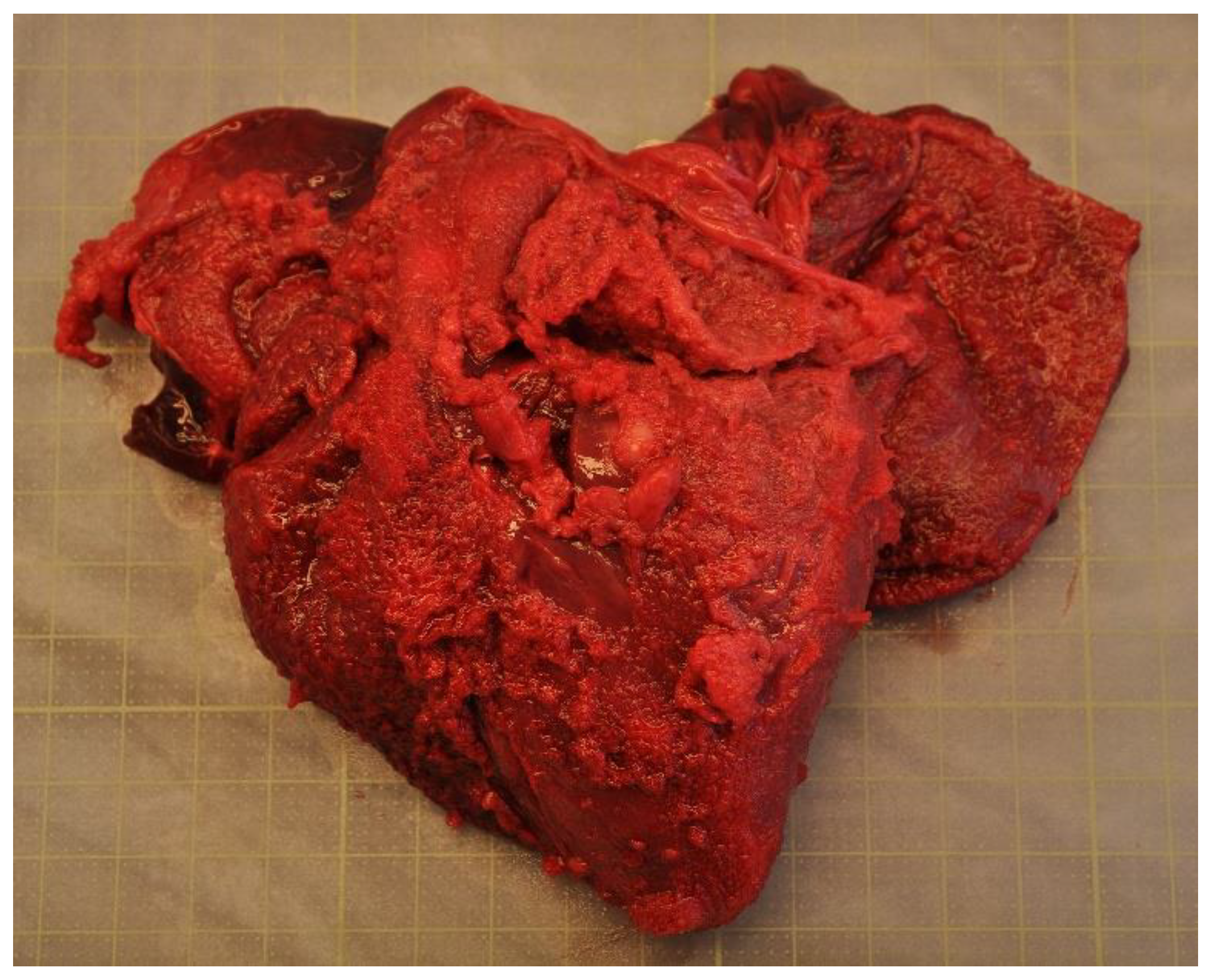
4.2.3. Non-Infectious Disease
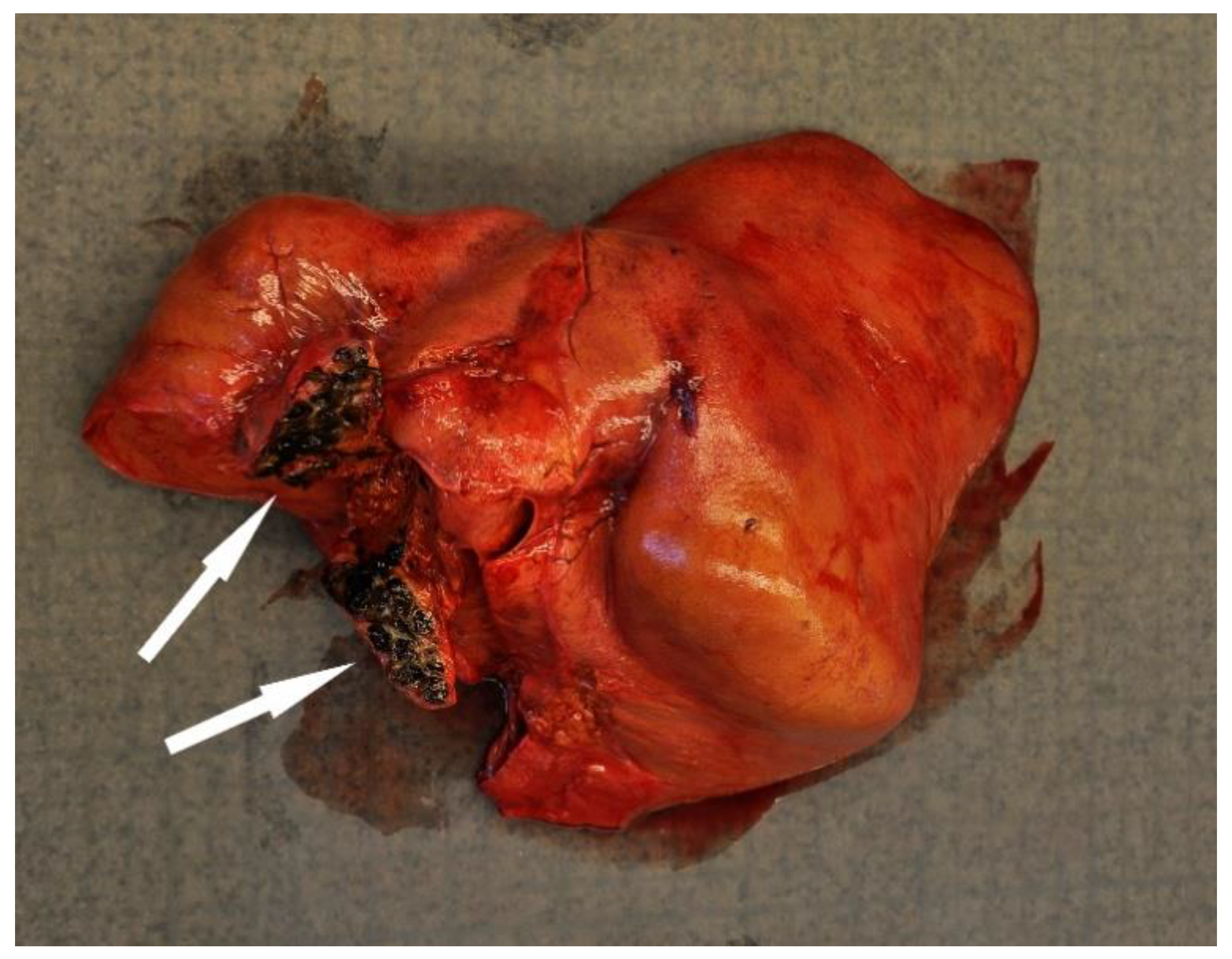
4.2.4. Trauma
4.2.5. Undetermined or Unsuitable Material
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Braulik, G.; Minton, G.; Amano, M.; Bjørge, A. Phocoena Phocoena, The IUCN Red List of Threatened Species 2020. 2020. Available online: https://www.iucnredlist.org/species/17027/50369903 (accessed on 28 December 2021).
- SAMBAH. Final Report for LIFE+ Project Static Acoustic Monitoring of the Baltic SeA Harbour Porpoise LIFE08 NAT/S/000261. 2016. Available online: http://www.sambah.org/SAMBAH-Final-Report-FINAL-for-website-April-2017.pdf (accessed on 28 December 2021).
- Leatherwood, S.; Reeves, R.R.; Foster, L. Sierra Club Handbook of Whales and Dolphins; Sierra Club Books: San Francisco, CA, USA, 1983. [Google Scholar]
- Read, A.J.; Hohn, A.A. Life in the fast lane: The life history of harbor porpoises from the Gulf of Maine. Mar. Mamm. Sci. 1995, 11, 423–440. [Google Scholar] [CrossRef]
- Brownell, R.L., Jr.; Reeves, R.R.; Read, A.J.; Smith, B.D.; Thomas, P.O.; Ralls, K.; Amano, M.; Berggren, P.; Chit, A.M.; Collins, T.; et al. Bycatch in gillnet fisheries threatens Critically Endangered small cetaceans and other aquatic megafauna. Endanger. Spec. Res. 2019, 40, 285–296. [Google Scholar] [CrossRef] [Green Version]
- Van Elk, C.E.; van de Bildt, M.W.G.; van Run, P.; Bunskoek, P.; Meerbeek, J.; Foster, G.; Osterhaus, A.D.M.E.; Kuiken, T. Clinical, pathological, and laboratory diagnoses of diseases of harbour porpoises (Phocoena phocoena), live stranded on the Dutch and adjacent coasts from 2003 to 2016. Vet. Res. 2019, 50, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siebert, U.; Wünschmann, A.; Weiss, R.; Frank, H.; Benke, H.; Frese, K. Post-mortem findings in harbour porpoises (Phocoena phocoena) from the German North and Baltic Seas. J. Comp. Pathol. 2001, 124, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Jauniaux, T.; Petitjean, D.; Brenez, C.; Borrens, M.; Brosens, L.; Haelters, J.; Tavernier, T.; Coignoul, F. Post-mortem findings and causes of death of harbour porpoises (Phocoena phocoena) stranded from 1990 to 2000 along the coastlines of Belgium and Northern France. J. Comp. Pathol. 2002, 126, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Kirkwood, J.; Bennett, P.; Jepson, P.; Kuiken, T.; Simpson, V.; Baker, J. Entanglement in fishing gear and other causes of death in cetaceans stranded on the coasts of England and Wales. Vet. Rec. 1997, 141, 94–98. [Google Scholar] [CrossRef]
- Fenton, H.; Daoust, P.Y.; Forzan, M.J.; Vanderstichel, R.V.; Ford, J.K.; Spaven, L.; Lair, S.; Raverty, S. Causes of mortality of harbor porpoises Phocoena phocoena along the Atlantic and Pacific coasts of Canada. Dis. Aquat. Organ. 2017, 122, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Read, A.J.; Murray, K.T. Gross Evidence of Human-Induced Mortality in Small Cetaceans. Available online: https://repository.library.noaa.gov/view/noaa/3679 (accessed on 28 December 2021).
- Kuiken, T.; Hartman, M.G. (Eds.) In Proceedings of the First European Cetacean Society Workshop on Cetacean Pathology: Dissection Techniques and Tissue Sampling, ECS Newsletter, Leiden, The Netherlands, 13–14 September 1991. No. 17, special issue 1991.
- Lockyer, C. Aspects of the biology of the harbour porpoise, Phocoena phocoena, from British waters. Dev. Mar. Biol. 1995, 4, 443–457. [Google Scholar]
- Koopman, H.N.; Pabst, D.A.; Mclellan, W.A.; Dillaman, R.; Read, A. Changes in blubber distribution and morphology associated with starvation in the harbor porpoise (Phocoena phocoena): Evidence for regional differences in blubber structure and function. Physiol. Biochem. Zool. 2002, 75, 498–512. [Google Scholar] [CrossRef] [Green Version]
- Bancroft, J.D.; Cook, H.C. Manual of Histological Techniques; Churchill Livingstone: Edinburgh, UK, 1984. [Google Scholar]
- Sandholt, A.K.; Neimanis, A.; Roos, A.; Eriksson, J.; Söderlund, R. Genomic signatures of host adaptation in group B Salmonella enterica ST416/ST417 from harbour porpoises. Vet. Res. 2021, 52, 1–11. [Google Scholar] [CrossRef]
- Boskani, T.; Edvinsson, B.; Wahab, T. Development of nineteen Taqman real-time PCR assays for screening and detection of select highly pathogenic bacteria. Infect. Ecol. Epidemiol. 2018, 8, 1553462. [Google Scholar] [CrossRef]
- Dagleish, M.P.; Perri, A.; Maley, M.; Ballingall, K.T.; Baily, J.L.; Davison, N.J.; Brownlow, A.C.; Rocchi, M.S. Novel Dermatitis and Relative Viral Nucleic Acid Tissue Loads in a Fin Whale (Balaenoptera physalus) with Systemic Cetacean Morbillivirus Infection. J. Comp. Pathol. 2021, 183, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Eurosurveill. 2020, 25, 23–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlén, I.; Thomas, L.; Carlström, J.; Amundin, M.; Teilmann, J.; Tregenza, N.; Tougaard, J.; Koblitz, J.C.; Sveegaard, S.; Wennerberg, D.; et al. Basin-scale distribution of harbour porpoises in the Baltic Sea provides basis for effective conservation actions. Biol. Conserv. 2018, 226, 42–53. [Google Scholar]
- HELCOM. Administrative Boundaries. Available online: https://metadata.helcom.fi/geonetwork/srv/eng/catalog.search#/metadata/b58b0b56-1a98-41d9-9bf4-5dc76f5069d3 (accessed on 28 December 2021).
- Seifert, T.; Tauber, F.; Kayser, B. A high resolution spherical grid topography of the Baltic Sea–revised edition. In Proceedings of the Baltic Sea Science Congress, Stockholm, Sweden, 25–29 November 2001; pp. 25–29. [Google Scholar]
- Sveegaard, S.; Galatius, A.; Dietz, R.; Kyhn, L.; Koblitz, J.C.; Amundin, M.; Nabe-Nielsen, J.; Sinding, M.H.; Andersen, L.W.; Teilmann, J. Defining management units for cetaceans by combining genetics, morphology, acoustics and satellite tracking. Glob Ecol Conserv. 2015, 3, 839–850. [Google Scholar]
- Lantmäteriet (Sverigekartan). Available online: https://www.lantmateriet.se/sv/Kartor-och-geografisk-information/geodataprodukter/produktlista/sverigekartor/ (accessed on 28 December 2021).
- Ryeng, K.A.; Lakemeyer, J.; Roller, M.; Wohlsein, P.; Siebert, U. Pathological findings in bycaught harbour porpoises (Phocoena phocoena) from the coast of Northern Norway. Polar Biol. 2022, 45, 45–57. [Google Scholar] [CrossRef]
- Peltier, H.; Baagøe, H.J.; Camphuysen, K.C.J.; Czeck, R.; Dabin, W.; Daniel, P.; Deaville, R.; Haelters, J.; Jauniaux, T.; Jensen, L.F.; et al. The stranding anomaly as population indicator: The case of harbour porpoise Phocoena phocoena in North-Western Europe. PLoS ONE 2013, 8, e62180. [Google Scholar]
- IJsseldijk, L.L.; Scheidat, M.; Siemensma, M.L.; Couperus, B.; Leopold, M.F.; Morell, M.; Gröne, A.; Kik, M.J.L. Challenges in the Assessment of Bycatch: Postmortem Findings in Harbor Porpoises (Phocoena phocoena) Retrieved from Gillnets. Vet. Path. 2021, 58, 405–415. [Google Scholar] [CrossRef]
- Jauniaux, T.; Delrez, N.; Haelters, J.; Kerckhof, F.; Coignoul, F. Causes of death of harbour porpoises (Phocoena phocoena) found in Belgium between 1990 and 2017. In Proceedings of the World Marine Mammal Conference, Barcelona, Spain, 9–12 December 2019. [Google Scholar]
- Siebert, U.; Tolley, K.; Vikingsson, G.; Olafsdottir, D.; Lehnert, K.; Weiss, R.; Baumgärtnerb, W. Pathological findings in harbour porpoises (Phocoena phocoena) from Norwegian and Icelandic waters. J. Comp. Pathol. 2006, 134, 134–142. [Google Scholar] [CrossRef]
- Cox, T.M.; Read, A.J.; Barco, S.; Evans, J.; Gannon, D.P.; Koopman, H.N.; McLellan, W.A.; Murray, K.; Nicolas, J.; Pabst, D.A.; et al. Documenting the bycatch of harbor porpoises, Phocoena phocoena, in coastal gillnet fisheries from stranded carcasses. Fish. Bull. 1998, 96, 727–734. [Google Scholar]
- Siebert, U.; Prenger-Berninghoff, E.; Weiss, R. Regional differences in bacterial flora in harbour porpoises from the North Atlantic: Environmental effects? J. Appl Microbiol. 2009, 106, 329–337. [Google Scholar] [CrossRef]
- Kuiken, T.; Bennett, P.M.; Allchin, C.R.; Kirkwood, J.K.; Baker, J.R.; Lockyer, C.H.; Walton, M.J.; Sheldrick, M.C. PCBs, cause of death and body condition in harbour porpoises (Phocoena phocoena) from British waters. Aquat. Toxicol. 1994, 28, 13–28. [Google Scholar] [CrossRef]
- Swenshon, M.; Lämmler, C.; Siebert, U. Identification and Molecular Characterization of Beta-Hemolytic Streptococci Isolated from Harbor Porpoises (Phocoena phocoena) of the North and Baltic Seas. J. Clin. Microbiol. 1998, 36, 1902–1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davison, N.; Simpson, V.; Chappell, S.; Monies, R.; Stubberfield, E.; Koylass, M.; Quinney, S.; Deaville, R.; Whatmore, A.M.; Jepson, P.D. Prevalence of a host-adapted group B Salmonella enterica in harbour porpoises (Phocoena phocoena) from the south-west coast of England. Vet. Rec. 2010, 167, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Siebert, U.; Müller, G.; Baumgärtner, W.; Weiss, R.; Desportes, G.; Hansen, K. Pyogranulomatous myocarditis due to Staphylococcus aureus septicaemia in two harbour porpoises (Phocoena phocoena). Vet. Rec. 2002, 150, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Delgado, J.; Arbelo, M.; Sierra, E.; Vela, A.; Domínguez, M.; Paz, Y.; Andrada, M.; Domínguez, L.; Fernández, A. Fatal Erysipelothrix rhusiopathiae septicemia in two Atlantic dolphins (Stenella frontalis and Tursiops truncatus). Dis. Aquat. Org. 2015, 116, 75–81. [Google Scholar] [CrossRef]
- Dagleish, M.; Barley, J.; Finlayson, J.; Reid, R.; Foster, G. Brucella ceti associated pathology in the testicle of a harbour porpoise (Phocoena phocoena). J. Comp. Pathol. 2008, 139, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.G.; Adams, L.G.; Ficht, T.A.; Cheville, N.F.; Payeur, J.P.; Harley, D.R.; House, C.; Ridgway, S.H. Brucella-induced abortions and infection in bottlenose dolphins (Tursiops truncatus). J. Zoo. Wildl. Med. 1999, 100–110. [Google Scholar]
- Perrett, L.L.; Dawson, C.E.; Davison, N.; Quinney, S. Brucella infection of lungworms from a harbour porpoise. Vet. Rec. 2004, 154, 800. [Google Scholar] [PubMed]
- Reckendorf, A.; Everaarts, E.; Bunskoek, P.; Haulena, M.; Springer, A.; Lehnert, K.; Lakemeyer, J.; Siebert, U.; Strube, C. Lungworm infections in harbour porpoises (Phocoena phocoena) in the German Wadden Sea between 2006 and 2018, and serodiagnostic tests. Int. J. Parasitol. Parasites Wildl. 2021, 14, 53–61. [Google Scholar] [CrossRef]
- Dzido, J.; Rolbiecki, L.; Izdebska, J.N.; Rokicki, J.; Kuczkowski, T.; Pawliczka, I. A global checklist of the parasites of the harbor porpoise Phocoena phocoena, a critically-endangered species, including new findings from the Baltic Sea. Int. J. Parasitol. Parasites Wildl. 2021, 15, 290–302. [Google Scholar] [CrossRef]
- Gibson, D.; Harris, E.; Bray, R.; Jepson, P.; Kuiken, T.; Baker, J.; Simpson, V.R. A survey of the helminth parasites of cetaceans stranded on the coast of England and Wales during the period 1990–1994. J. Zool. 1998, 244, 563–574. [Google Scholar] [CrossRef]
- Van Bressem, M.-F.; Duignan, P.J.; Banyard, A.; Barbieri, M.; Colegrove, K.M.; De Guise, S.; di Guardo, G.; Dobson, A.; Domingo, M.; Fauquier, D.; et al. Cetacean morbillivirus: Current knowledge and future directions. Viruses 2014, 6, 5145–5181. [Google Scholar] [CrossRef] [PubMed]
- Van Beurden, S.J.; Ijsseldijk, L.L.; Ordonez, S.R.; Förster, C.; de Vrieze, G.; Gröne, A.; Verheije, M.H.; Kik, M. Identification of a novel gammaherpesvirus associated with (muco) cutaneous lesions in harbour porpoises (Phocoena phocoena). Arch. Virol. 2015, 160, 3115–3120. [Google Scholar] [CrossRef] [PubMed]
- Mathavarajah, S.; Stoddart, A.K.; Gagnon, G.A.; Dellaire, G. Pandemic danger to the deep: The risk of marine mammals contracting SARS-CoV-2 from wastewater. Sci. Total. Environ. 2021, 760, 143346. [Google Scholar] [CrossRef]
- Williams, R.; ten Doeschate, M.; Curnick, D.J.; Brownlow, A.; Barber, J.L.; Davison, N.J.; Deaville, R.; Perkins, M.; Jepson, P.D.; Jobling, S. Levels of polychlorinated biphenyls are still associated with toxic effects in harbor porpoises (Phocoena phocoena) despite having fallen below proposed toxicity thresholds. Environ. Sci. Technol. 2020, 54, 2277–2286. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.; Barber, J.L.; Learmonth, J.A.; Read, F.L.; Deaville, R.; Perkins, M.W.; Brownlow, A.; Davison, N.; Penrose, R.; Pierce, G.J.; et al. Reproductive failure in UK harbour porpoises phocoena phocoena: Legacy of pollutant exposure? PLoS ONE 2015, 10, e0131085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leopold, M.F.; Begeman, L.; van Bleijswijk, J.D.; IJsseldijk, L.L.; Witte, H.J.; Gröne, A. Exposing the grey seal as a major predator of harbour porpoises. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stringell, T.; Hill, D.; Rees, D.; Rees, F.; Rees, P.; Morgan, G.; Morgan, L.; Morris, C. Predation of harbour porpoises (Phocoena phocoena) by grey seals (Halichoerus grypus) in Wales. Aquat. Mamm. 2015, 41, 188. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Identification Number | Sex | Age Class | Bacteria Cultured | Tissue(s) Cultured | Lesion |
|---|---|---|---|---|---|
| 12-VLT000154 | M | Calf | Staphylococcus aureus | Pericardium, lymph node | Myocarditis, pericarditis, lymphadenitis |
| 16-VLT001592 | M | Adult | Brucella sp. * | Testis | Orchitis |
| 17-VLT002645 | F | Adult | Edwardsiella tarda | Heart valve | Endocarditis and pulmonary thrombosis |
| 17-VLT002646 | F | Adult | Streptococcus canis | Sepsis | |
| 17-VLT002652 | M | Adult | group B Salmonella enterica ST416/ST417 | Lung | Pneumonia |
| 18-VLT001113 | M | Adult | Schwanella putrefaciens | Lung | Pneumonia |
| 19-VLT002835 | F | Juvenile | Streptococcus sp. | Lung | Pneumonia |
| 19-VLT002839 | F | Adult | Streptococcus sp. | Lung | Pneumonia |
| 19-VLT002851 | M | Calf | Erysipelothrix rhusiopathiae | Lung | Pneumonia |
| 20-VLT001389 | F | Juvenile | group B Salmonella enterica ST416/ST417 | Lung | Pneumonia ** |
| 20-VLT001392 | M | Calf | Streptococcus phocae | Dermal abscess, lung, spleen | Sepsis |
| 20-VLT002367 | M | Adult | Erysipelothrix rhusiopathiae | Lung | Pneumonia |
| Cause of Death | Neonate | Calf | Juvenile | Adult | Total |
|---|---|---|---|---|---|
| Bycatch | 0 | 7 | 4 | 2 | 13 |
| Probable bycatch | 1 | 7 | 5 | 8 | 21 |
| Infectious disease | 0 | 6 | 3 | 10 | 19 |
| Non-infectious disease | 2 | 0 | 0 | 2 | 4 |
| Trauma | 0 | 5 | 3 | 3 | 11 |
| Emaciation | 3 | 4 | 1 | 3 | 11 |
| Abandoned | 9 | 0 | 0 | 0 | 9 |
| Undetermined | 2 | 6 | 5 | 7 | 20 |
| Unsuitable | 4 | 5 | 5 | 6 | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neimanis, A.; Stavenow, J.; Ågren, E.O.; Wikström-Lassa, E.; Roos, A.M. Causes of Death and Pathological Findings in Stranded Harbour Porpoises (Phocoena phocoena) from Swedish Waters. Animals 2022, 12, 369. https://doi.org/10.3390/ani12030369
Neimanis A, Stavenow J, Ågren EO, Wikström-Lassa E, Roos AM. Causes of Death and Pathological Findings in Stranded Harbour Porpoises (Phocoena phocoena) from Swedish Waters. Animals. 2022; 12(3):369. https://doi.org/10.3390/ani12030369
Chicago/Turabian StyleNeimanis, Aleksija, Jasmine Stavenow, Erik Olof Ågren, Emil Wikström-Lassa, and Anna Maria Roos. 2022. "Causes of Death and Pathological Findings in Stranded Harbour Porpoises (Phocoena phocoena) from Swedish Waters" Animals 12, no. 3: 369. https://doi.org/10.3390/ani12030369
APA StyleNeimanis, A., Stavenow, J., Ågren, E. O., Wikström-Lassa, E., & Roos, A. M. (2022). Causes of Death and Pathological Findings in Stranded Harbour Porpoises (Phocoena phocoena) from Swedish Waters. Animals, 12(3), 369. https://doi.org/10.3390/ani12030369