
Effects of the Antimicrobial Peptide Mastoparan X on the Performance, Permeability and Microbiota Populations of Broiler Chickens
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Peptide Synthesis
2.3. Experimental Animals
2.4. Production Performance
2.5. Dissection and Collection of Viscera and Intestinal Samples
2.6. Gene Expression in the Intestine
2.7. Determination of the Immune Organ Index
2.8. Observation of the Intestinal Morphology
2.9. Microbiome Total DNA Extraction and PCR Amplification of Target Fragments
2.10. Magnetic Bead Purification and Recovery
2.11. Fluorescence-Based Quantification of the Amplified Products
2.12. High-Throughput Sequencing
2.13. Statistical Analysis
3. Results
3.1. Effects of MPX on the Performance of Chickens
3.2. Effects of MPX on the Intestinal Morphology and Structure
3.3. Effects of MPX on the Immune Organ Index
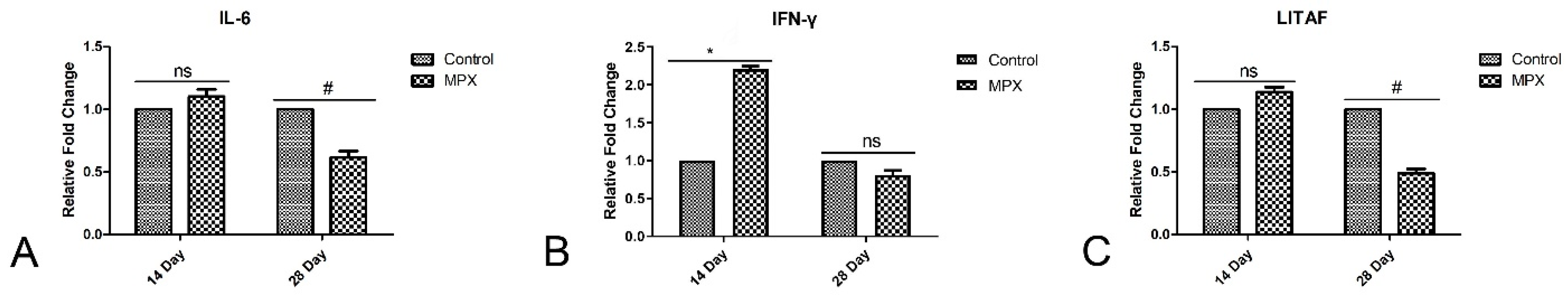
3.4. Effects of MPX on Immunological Markers
3.5. Effects of MPX on Jejunum Barrier-Associated Proteins
3.6. Effects of MPX on Nutrient Transporters and Digestive Enzymes
3.7. Effects of MPX on the Intestinal Antioxidant Capacity
3.8. Effects of MPX on the Cecal Microbial Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Xie, Z.; Zhao, Q.; Wang, H.; Wen, L.; Li, W.; Zhang, X.; Lin, W.; Li, H.; Xie, Q.; Wang, Y. Effects of antibacterial peptide combinations on growth performance, intestinal health, and immune function of broiler chickens. Poult. Sci. 2020, 99, 6481–6492. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.B.; Hwang, S.; Yang, J.H.; Lee, S.; Bang, W.Y.; Moon, K.H. Dynamics of the Gut Microbiome and Transcriptome in Korea Native Ricefish (Oryzias latipes) during Chronic Antibiotic Exposure. Genes 2022, 13, 1243. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhan, Y.; Ma, W.; Zhu, Y.; Wang, Z. Effects of Antimicrobial peptides on egg production, egg quality and caecal microbiota of hens during the late laying period. Anim. Sci. J. 2020, 91, e13387. [Google Scholar] [CrossRef] [PubMed]
- Stepien-Pysniak, D.; Marek, A.; Banach, T.; Adaszek, L.; Pyzik, E.; Wilczynski, J.; Winiarczyk, S. Prevalence and antibiotic resistance of Enterococcus strains isolated from poultry. Acta Vet. Hung. 2016, 64, 148–163. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, Q.; Wen, L.; Wu, C.; Yao, Z.; Yan, Z.; Li, R.; Chen, L.; Chen, F.; Xie, Z.; et al. The Effect of the Antimicrobial Peptide Plectasin on the Growth Performance, Intestinal Health, and Immune Function of Yellow-Feathered Chickens. Front. Vet. Sci. 2021, 8, 688611. [Google Scholar] [CrossRef]
- Paul, S.S.; Vantharam, V.H.; Raju, M.V.; Rama, R.S.; Nori, S.S.; Suryanarayan, S.; Kumar, V.; Perveen, Z.; Prasad, C.S. Dietary supplementation of extracts of red sea weed (Kappaphycus alvarezii) improves growth, intestinal morphology, expression of intestinal genes and immune responses in broiler chickens. J. Sci. Food Agric. 2021, 101, 997–1008. [Google Scholar] [CrossRef]
- Koorakula, R.; Schiavinato, M.; Ghanbari, M.; Wegl, G.; Grabner, N.; Koestelbauer, A.; Klose, V.; Dohm, J.C.; Domig, K.J. Metatranscriptomic Analysis of the Chicken Gut Resistome Response to In-Feed Antibiotics and Natural Feed Additives. Front. Microbiol. 2022, 13, 833790. [Google Scholar] [CrossRef]
- Rathnayake, D.; Mun, H.S.; Dilawar, M.A.; Baek, K.S.; Yang, C.J. Time for a Paradigm Shift in Animal Nutrition Metabolic Pathway: Dietary Inclusion of Organic Acids on the Production Parameters, Nutrient Digestibility, and Meat Quality Traits of Swine and Broilers. Life 2021, 11, 476. [Google Scholar] [CrossRef]
- Li, Y.; Yang, Q.; Cai, D.; Guo, H.; Fang, J.; Cui, H.; Gou, L.; Deng, J.; Wang, Z.; Zuo, Z. Resistin, a Novel Host Defense Peptide of Innate Immunity. Front Immunol. 2021, 12, 699807. [Google Scholar] [CrossRef]
- Xia, X.; Cheng, L.; Zhang, S.; Wang, L.; Hu, J. The role of natural antimicrobial peptides during infection and chronic inflammation. Antonie Van Leeuwenhoek 2018, 111, 5–26. [Google Scholar] [CrossRef]
- Phoenix, D.A.; Harris, F.; Dennison, S.R. Antimicrobial Peptides with pH-Dependent Activity and Alkaline Optima: Their Origins, Mechanisms of Action and Potential Applications. Curr. Protein Pept. Sci. 2021, 22, 775–799. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Wang, L.; Xie, Y.; Chen, Y.; Yi, H.; He, D. Effects of antimicrobial peptide cathelicidin-BF on diarrhea controlling, immune responses, intestinal inflammation and intestinal barrier function in piglets with postweaning diarrhea. Int Immunopharmacol. 2020, 85, 106658. [Google Scholar] [CrossRef] [PubMed]
- Zeiders, S.M.; Chmielewski, J. Antibiotic-cell-penetrating peptide conjugates targeting challenging drug-resistant and intracellular pathogenic bacteria. Chem. Biol. Drug. Des. 2021, 98, 762–778. [Google Scholar] [CrossRef] [PubMed]
- Jangpromma, N.; Konkchaiyaphum, M.; Punpad, A.; Sosiangdi, S.; Daduang, S.; Klaynongsruang, S.; Tankrathok, A. Rational Design of RN15m4 Cathelin Domain-Based Peptides from Siamese Crocodile Cathelicidin Improves Antimicrobial Activity. Appl. Biochem. Biotechnol. 2022; Online ahead of print. [Google Scholar]
- Bao, H.; She, R.; Liu, T.; Zhang, Y.; Peng, K.S.; Luo, D.; Yue, Z.; Ding, Y.; Hu, Y.; Liu, W. Effects of pig antibacterial peptides on growth performance and intestine mucosal immune of broiler chickens. Poult. Sci. 2009, 88, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zeng, X.; Yang, Q.; Qiao, S. Antimicrobial Peptides as Potential Alternatives to Antibiotics in Food Animal Industry. Int. J. Mol. Sci. 2016, 17, 603. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, M.P.; Alvares, D.S.; Leite, N.B.; de Souza, B.M.; Palma, M.S.; Riske, K.A.; Neto, J.R. New insight into the mechanism of action of wasp mastoparan peptides: Lytic activity and clustering observed with giant vesicles. Langmuir 2011, 27, 10805–10813. [Google Scholar] [CrossRef]
- da Silva, A.M.B.; Silva-Gonçalves, L.C.; Oliveira, F.A.; Arcisio-Miranda, M. Pro-necrotic Activity of Cationic Mastoparan Peptides in Human Glioblastoma Multiforme Cells via Membranolytic Action. Mol. Neurobiol. 2018, 55, 5490–5504. [Google Scholar] [CrossRef]
- Henriksen, J.R.; Etzerodt, T.; Gjetting, T.; Andresen, T.L. Side chain hydrophobicity modulates therapeutic activity and membrane selectivity of antimicrobial peptide mastoparan-X. PLoS ONE 2014, 9, e91007. [Google Scholar] [CrossRef]
- Zhu, C.L.; Wang, L.; Zhao, X.Q.; Yang, R.; Zhang, B.Y.; Zhao, Y.Y.; Xia, X.J.; Zhang, H.H.; Chen, S.J.; Jiang, J.Q.; et al. Antimicrobial peptide MPX attenuates LPS-induced inflammatory response and blood-testis barrier dysfunction in Sertoli cells. Theriogenology 2022, 189, 301–312. [Google Scholar] [CrossRef]
- Zhu, C.; Zhao, Y.; Zhao, X.; Liu, S.; Xia, X.; Zhang, S.; Wang, Y.; Zhang, H.; Xu, Y.; Chen, S.; et al. The Antimicrobial Peptide MPX Can Kill Staphylococcus aureus, Reduce Biofilm Formation, and Effectively Treat Bacterial Skin Infections in Mice. Front. Vet. Sci. 2022, 9, 819921. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, L.; Zhu, C.; Xia, X.; Zhang, S.; Wang, Y.; Zhang, H.; Xu, Y.; Chen, S.; Jiang, J.; et al. The Antimicrobial Peptide Mastoparan X Protects Against Enterohemorrhagic Escherichia coli O157:H7 Infection, Inhibits Inflammation, and Enhances the Intestinal Epithelial Barrier. Front Microbiol. 2021, 12, 644887. [Google Scholar] [CrossRef] [PubMed]
- Chasiotis, H.; Kolosov, D.; Bui, P.; Kelly, S.P. Tight junctions, tight junction proteins and paracellular permeability across the gill epithelium of fishes: A review. Respir. Physiol. Neurobiol. 2012, 184, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Wang, Z.; Cui, H.; Nie, H.; Zhang, T.; Gao, X.; Qiao, Y. Effects of enzymatic hydrolysate of locust bean gum on digestibility, intestinal morphology and microflora of broilers. J. Anim. Physiol. Anim. Nutr. 2020, 104, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Omonijo, F.A.; Liu, S.; Hui, Q.; Zhang, H.; Lahaye, L.; Bodin, J.C.; Gong, J.; Nyachoti, M.; Yang, C. Thymol Improves Barrier Function and Attenuates Inflammatory Responses in Porcine Intestinal Epithelial Cells during Lipopolysaccharide (LPS)-Induced Inflammation. J. Agric. Food Chem. 2019, 67, 615–624. [Google Scholar] [CrossRef]
- Wickramasuriya, S.S.; Park, I.; Lee, Y.; Kim, W.H.; Przybyszewski, C.; Gay, C.G.; van Oosterwijk, J.G.; Lillehoj, H.S. Bacillus subtilis Oral Delivery of Expressing Chicken NK-2 Peptide Protects Against Infection in Broiler Chickens. Front. Vet. Sci. 2021, 8, 684818. [Google Scholar] [CrossRef]
- Venardou, B.; O’Doherty, J.V.; Vigors, S.; O’Shea, C.J.; Burton, E.J.; Ryan, M.T.; Sweeney, T. Effects of dietary supplementation with a laminarin-rich extract on the growth performance and gastrointestinal health in broilers. Poult. Sci. 2021, 100, 101179. [Google Scholar] [CrossRef]
- Maropola, M.K.; Ramond, J.B.; Trindade, M. Impact of metagenomic DNA extraction procedures on the identifiable endophytic bacterial diversity in Sorghum bicolor (L. Moench). J. Microbiol. Methods 2015, 112, 104–117. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, G.; Siegel, P.; He, C.; Wang, H.; Zhao, W.; Zhai, Z.; Tian, F.; Zhao, J.; Zhang, H.; et al. Quantitative genetic background of the host influences gut microbiomes in chickens. Sci. Rep. 2013, 3, 1163. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods. 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef]
- Li, X.; Zuo, S.; Wang, B.; Zhang, K.; Wang, Y. Antimicrobial Mechanisms and Clinical Application Prospects of Antimicrobial Peptides. Molecules 2022, 27, 2675. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, A.R.; Wellington, D.A.; Kumar, P.; Kassa, H.; Booth, C.J.; Cresswell, P.; MacMicking, J.D. GBP5 promotes NLRP3 inflammasome assembly and immunity in mammals. Science 2012, 336, 481–485. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Lee, J.; Vu, T.H.; Lee, S.; Lillehoj, H.S.; Hong, Y.H. Chicken avian beta-defensin 8 modulates immune response via the mitogen-activated protein kinase signaling pathways in a chicken macrophage cell line. Poult. Sci. 2020, 99, 4174–4182. [Google Scholar] [CrossRef]
- Zhu, F.; Zhang, B.; Li, J.; Zhu, L. Effects of fermented feed on growth performance, immune response, and antioxidant capacity in laying hen chicks and the underlying molecular mechanism involving nuclear factor-kappaB. Poult. Sci. 2020, 99, 2573–2580. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Ai, W.; Ye, J.; Wang, C.; Yuan, S.; Xie, Y.; Mo, X.; Li, W.; He, Z.; Chen, Y.; et al. Inflammatory markers and risk factors of RA patients with depression and application of different scales in judging depression. Clin. Rheumatol. 2022, 41, 2309–2317. [Google Scholar] [CrossRef]
- Karshikoff, B.; Astrom, J.; Holmstrom, L.; Lekander, M.; Kemani, M.K.; Wicksell, R.K. Baseline Pro-Inflammatory Cytokine Levels Moderate Psychological Inflexibility in Behavioral Treatment for Chronic Pain. J. Clin. Med. 2022, 11, 2285. [Google Scholar] [CrossRef]
- Oshima, T.; Miwa, H. Gastrointestinal mucosal barrier function and diseases. J. Gastroenterol. 2016, 51, 768–778. [Google Scholar] [CrossRef]
- Schneeberger, E.E.; Lynch, R.D. The tight junction: A multifunctional complex. Am. J. Physiol. Cell Physiol. 2004, 286, C1213–C1228. [Google Scholar] [CrossRef]
- Daneshmand, A.; Kermanshahi, H.; Sekhavati, M.H.; Javadmanesh, A.; Ahmadian, M.; Alizadeh, M.; Aldawoodi, A. Effects of cLFchimera peptide on intestinal morphology, integrity, microbiota, and immune cells in broiler chickens challenged with necrotic enteritis. Sci. Rep. 2020, 10, 17704. [Google Scholar] [CrossRef]
- Dai, F.; Lin, T.; Cheng, L.; Wang, J.; Zuo, J.; Feng, D. Effects of micronized bamboo powder on growth performance, serum biochemical indexes, cecal chyme microflora and metabolism of broilers aged 1-22 days. Trop. Anim. Health Prod. 2022, 54, 166. [Google Scholar] [CrossRef]
- Vlasova, A.N.; Takanashi, S.; Miyazaki, A.; Rajashekara, G.; Saif, L.J. How the gut microbiome regulates host immune responses to viral vaccines. Curr. Opin. Virol. 2019, 37, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.S.; Zheng, Y.; Huang, Z.; Min, W.X.; Yang, H. Lactococcus nasutitermitis sp. nov. isolated from a termite gut. Int. J. Syst. Evol. Microbiol. 2016, 66, 518–522. [Google Scholar] [CrossRef]
- Wang, K.; Wang, K.; Wang, J.; Yu, F.; Ye, C. Protective Effect of Clostridium butyricum on Escherichia coli-Induced Endometritis in Mice via Ameliorating Endometrial Barrier and Inhibiting Inflammatory Response. Microbiol. Spectr. 2022, e0328622. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, I.I.; Atarashi, K.; Manel, N.; Brodie, E.L.; Shima, T.; Karaoz, U.; Wei, D.; Goldfarb, K.C.; Santee, C.A.; Lynch, S.V.; et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell 2009, 139, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.S.; Gong, Y.N.; Chi, C.Y.; Hsieh, S.Y.; Chen, W.T.; Ku, C.L.; Cheng, H.T.; Lin, L.; Sung, C.M. Gut microbiome profiles and associated metabolic pathways in patients of adult-onset immunodeficiency with anti-interferon-gamma autoantibodies. Sci. Rep. 2022, 12, 9126. [Google Scholar] [CrossRef]
- Liu, G.; Zhu, H.; Ma, T.; Yan, Z.; Zhang, Y.; Geng, Y.; Zhu, Y.; Shi, Y. Effect of chronic cyclic heat stress on the intestinal morphology, oxidative status and cecal bacterial communities in broilers. J. Therm. Biol. 2020, 91, 102619. [Google Scholar] [CrossRef]
- Yang, L.; Chen, L.; Zheng, K.; Ma, Y.J.; He, R.X.; Arowolo, M.A.; Zhou, Y.J.; Xiao, D.F.; He, J.H. Effects of fenugreek seed extracts on growth performance and intestinal health of broilers. Poult. Sci. 2022, 101, 101939. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Pre-Starter (Days 1–14) | Starter (Days 15–28) | Finisher (29 Days Onward) |
|---|---|---|---|
| Ingredients | |||
| Corn | 54.2 | 57.2 | 59.2 |
| Soya bean meal | 34 | 31.5 | 22.5 |
| Rapeseed meal | 5.0 | 5.0 | 9.5 |
| Calcium bicarbonate | 1.5 | 1.3 | 1.3 |
| Mountain flour | 1.0 | 1.2 | 1.2 |
| Salt | 0.3 | 0.3 | 0.3 |
| Oil | 3.0 | 2.5 | 3.0 |
| Wheat bran | 2.0 | ||
| Premix 1 | 1.0 | 1.0 | 1.0 |
| Nutrient composition | |||
| Metabolizable energy (MJ/kg) | 12.54 | 12.54 | 12.96 |
| Crude protein (%) | 21.50 | 21.50 | 20.00 |
| Total phosphorus (%) | 0.68 | 0.68 | 0.65 |
| Lys | 1.15 | 1.15 | 1.03 |
| Met | 0.51 | 0.51 | 0.41 |
| Met + Cys | 0.87 | 0.87 | 0.76 |
| Gene | Group | Primer Sequence (5′-3′) | Reference |
|---|---|---|---|
| HMOX1 | Anti-oxidant gene | F-CTGGAGAAGGGTTGGCTTTCT | [26] |
| R-GAAGCTCTGCCTTTGGCTGTA | |||
| CAT | F-ACTGCAAGGCGAAAGTGTTT | [26] | |
| R-GGCTATGGATGAAGGATGGA | |||
| SOD1 | F-ATTACCGGCTTGTCTGATGG | [26] | |
| R-CCTCCCTTTGCAGTCACATT | |||
| JAM-2 | Markers of intestinal integrity | F-AGCCTCAAATGGGATTGGATT | [26] |
| R-CATCAACTTGCATTCGCTTCA | |||
| ZO-1 | F-GCCTGAATCAAACCCAGCAA | [1] | |
| R-TATGCGGCGGTAAGGATGAT | |||
| Occludin | F-CCCAGAAGACGCGCAGTAAG | [27] | |
| R-GCGCGGTCCCAGTAGATG | |||
| MUC2 | F-TTCATGATGCCTGCTCTTGTC | [1] | |
| R-CCGTAGCCTTGGTACATTCTTGT | |||
| Claudin | F-CATACTCCTGGGTCTGGTTGGT | [6] | |
| R-GACAGCCATCCGCATCTTCT | |||
| FABP2 | Nutrient transporters and digestive enzymes | F-CTTGGAAAATAGAGAAAAATGAGAACTATG | [27] |
| R-GGCTCCTAACTTTCTTTTCATCACA | |||
| SLC15A2/PepT2 | F-CGAAACTCTGTGGCTCCAACT | [27] | |
| R-CGCTCGCAGAACTCGTTCA | |||
| SLC2A5/GLUT5 | F-GGATCAATGCAGTCTTCTACTATGCA | [27] | |
| R-CACCTATGGACACGGTGACATACT | |||
| AMY2A | F-CACGGGCACCCACTCAAC | [27] | |
| R-GGCACAGCGGGAAAATCTC | |||
| LITAF | Markers of immune response | F-CCCCTACCCTGTCCCACAA | [27] |
| R-TGAGTACTGCGGAGGGTTCAT | |||
| IFN-γ | F-ATCATACTGAGCCAGATTGTTTCG | [1] | |
| R-TCTTTCACCTTCTTCACGCCAT | |||
| IL-6 | F- GTGTGCGAGAACAGCATGGAGA | [5] | |
| R-CTGGAGAGCTTCGTCAGGCATT | |||
| ACTB | Housekeeping reference genes | F-TGCTGCGCTCGTTGTTGA | [27] |
| R-CGTCCCCGGCGAAA | |||
| GAPDH | F-GCTGTGGAGAGATGGCAGAGGT | [5] | |
| R-ACGGCAGGTCAGGTCAACAACA |
| Stage | Index | Control | MPX |
|---|---|---|---|
| 1 day | BW (g) | 44.26 ± 1.03 a | 45.13 ± 0.97 a |
| 14 days | BW (g) | 235.75 ± 0.86 a | 235.45 ± 0.58 a |
| 28 days | BW (g) | 684.59 ± 0.49 a | 719.57 ± 1.21 b |
| 42 days | BW (g) | 1260.97 ± 1.74 a | 1322.55 ± 0.53 b |
| 1–14 days | ADG/g | 14.73 ± 0.55 b | 14.64 ± 0.50 b |
| ADFI/g | 29.03 ± 0.12 a | 28.85 ± 0.58 a | |
| FCR | 1.97 ± 0.08 a | 1.98 ± 0.05 a | |
| 15–28 days | ADG/g | 32.06 ± 1.72 b | 34.58 ± 1.35 a |
| ADFI/g | 53.45 ± 0.58 a | 52.28 ± 1.31 a | |
| FCR | 1.67 ± 0.09 a | 1.51 ± 0.05 a | |
| 29–42 days | ADG/g | 41.17 ± 1.26 b | 43.07 ± 1.34 a |
| ADFI/g | 92.67 ± 0.93 b | 93.90 ± 0.71 a | |
| FCR | 2.24 ± 0.01 a | 2.18 ± 0.03 b | |
| 1–42 days | ADG/g | 24.87 ± 1.5 b | 28.88 ± 2.04 a |
| ADFI/g | 74.48 ± 1.3 a | 76.64 ± 0.77 b | |
| FCR | 2.95 ± 0.09 a | 2.7 ± 0.05 b | |
| 1–14 days | Mortality (%) | 6 (3/50) | 4 (2/50) |
| 15–28 days | Mortality (%) | 0 | 0 |
| 29–42 days | Mortality (%) | 0 | 0 |
| 1–42 days | Mortality (%) | 6 (3/50) | 4 (2/50) |
| Site | Index | Control | MPX |
|---|---|---|---|
| lleum | Villus length | 683.33 ± 9.61 a | 681.67 ± 9.61 a |
| Crypt depth | 156.67 ± 7.09 a | 154.33 ± 11.02 a | |
| Villus length/Crypt depth | 4.37 ± 0.16 a | 4.43 ± 0.34 a | |
| Jejunum | Villus length | 974.33 ± 6.51 a | 985.67 ± 9.87 b |
| Crypt depth | 163.67 ± 8.33 a | 151.00 ± 8.54 b | |
| Villus length/Crypt depth | 5.96 ± 0.31 a | 6.53 ± 0.32 b | |
| Duodenum | Villus length | 1356.00 ± 13.11 a | 1423.67 ± 11.06 b |
| Crypt depth | 255.33 ± 7.09 a | 256.33 ± 9.07 a | |
| Villus length/Crypt depth | 5.31 ± 0.20 a | 5.56 ± 0.18 a |
| Stage | Indicator | Control | MPX |
|---|---|---|---|
| Day 14 | Thymus | 2.01 ± 0.61 a | 1.10 ± 0.68 b |
| Bursa | 2.45 ± 0.18 b | 2.95 ± 0.85 a | |
| Spleen | 0.68 ± 0.12 a | 0.97 ± 0.30 a | |
| Day 28 | Thymus | 2.03 ± 0.66 b | 2.41 ± 1.05 a |
| Bursa | 2.27 ± 0.48 b | 2.67 ± 0.92 a | |
| Spleen | 1.08 ± 0.25 a | 0.91 ± 0.21 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, C.; Bai, Y.; Xia, X.; Zhang, M.; Wu, X.; Wu, Y.; Bai, Y.; Liu, S.; Zhang, G.; Hu, J.; et al. Effects of the Antimicrobial Peptide Mastoparan X on the Performance, Permeability and Microbiota Populations of Broiler Chickens. Animals 2022, 12, 3462. https://doi.org/10.3390/ani12243462
Zhu C, Bai Y, Xia X, Zhang M, Wu X, Wu Y, Bai Y, Liu S, Zhang G, Hu J, et al. Effects of the Antimicrobial Peptide Mastoparan X on the Performance, Permeability and Microbiota Populations of Broiler Chickens. Animals. 2022; 12(24):3462. https://doi.org/10.3390/ani12243462
Chicago/Turabian StyleZhu, Chunling, Yilin Bai, Xiaojing Xia, Man Zhang, Xilong Wu, Yundi Wu, Yueyu Bai, Shanqin Liu, Gaiping Zhang, Jianhe Hu, and et al. 2022. "Effects of the Antimicrobial Peptide Mastoparan X on the Performance, Permeability and Microbiota Populations of Broiler Chickens" Animals 12, no. 24: 3462. https://doi.org/10.3390/ani12243462
APA StyleZhu, C., Bai, Y., Xia, X., Zhang, M., Wu, X., Wu, Y., Bai, Y., Liu, S., Zhang, G., Hu, J., Fotina, H., Wang, L., & Zhao, X. (2022). Effects of the Antimicrobial Peptide Mastoparan X on the Performance, Permeability and Microbiota Populations of Broiler Chickens. Animals, 12(24), 3462. https://doi.org/10.3390/ani12243462