
Daily Rhythms in the IGF-1 System in the Liver of Goldfish and Their Synchronization to Light/Dark Cycle and Feeding Time

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
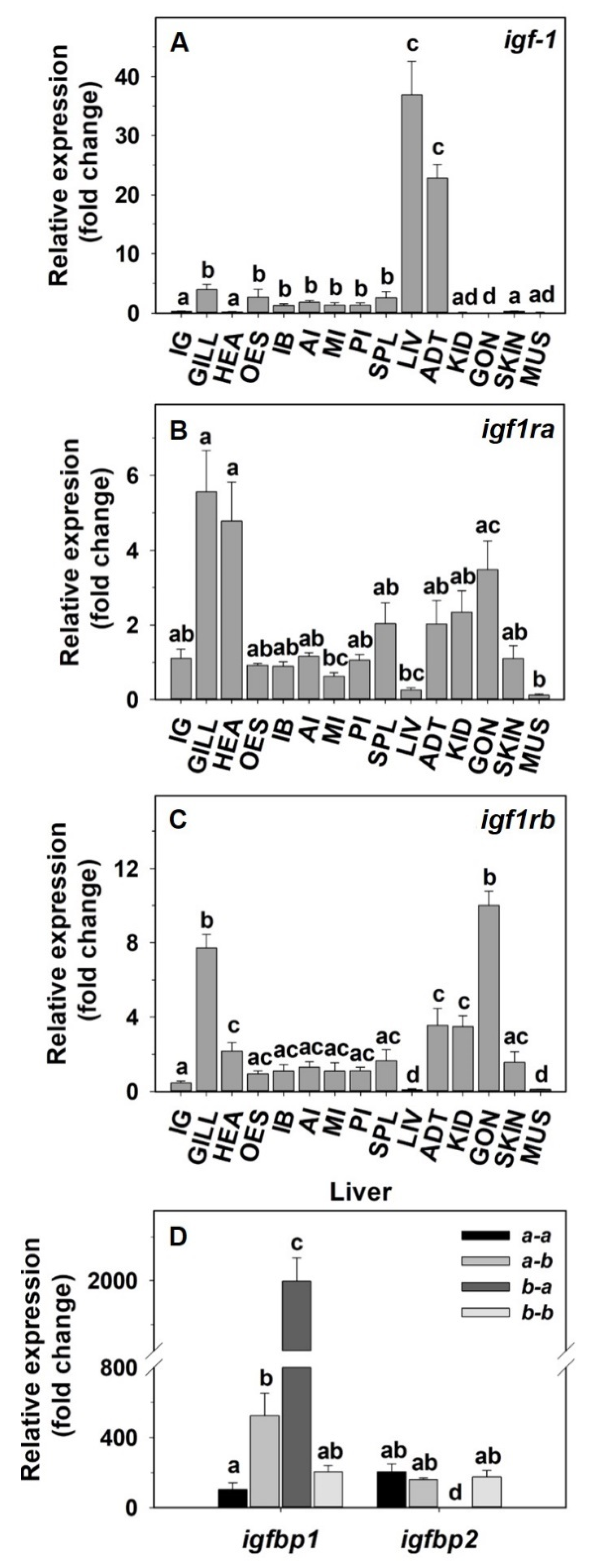
2.2. Tissue Distribution of the IGF-1 System in Goldfish
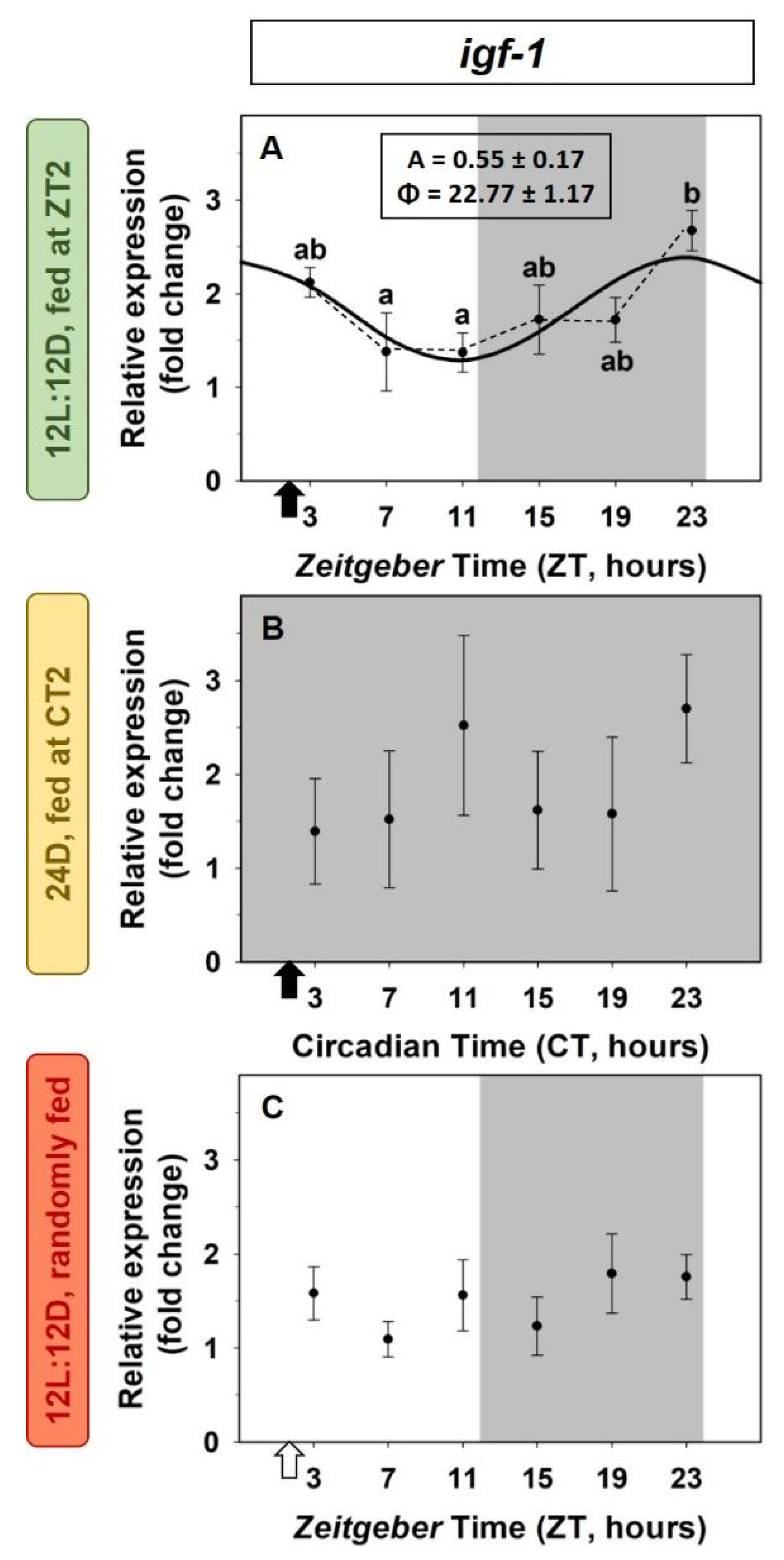
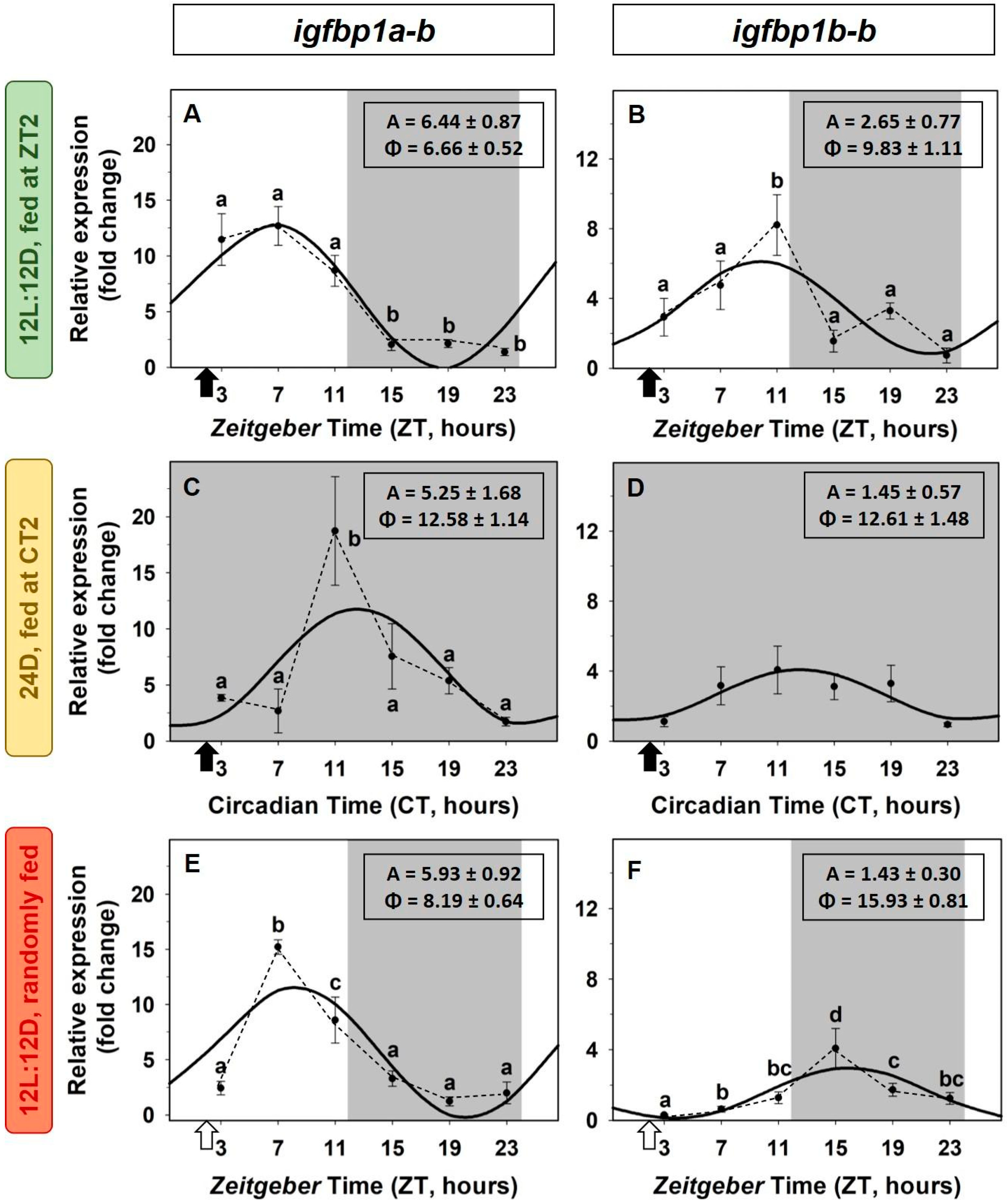
2.3. Daily Variations of the IGF-1 System in the Liver. Light/Dark Cycle and Scheduled Feeding as Zeitgebers
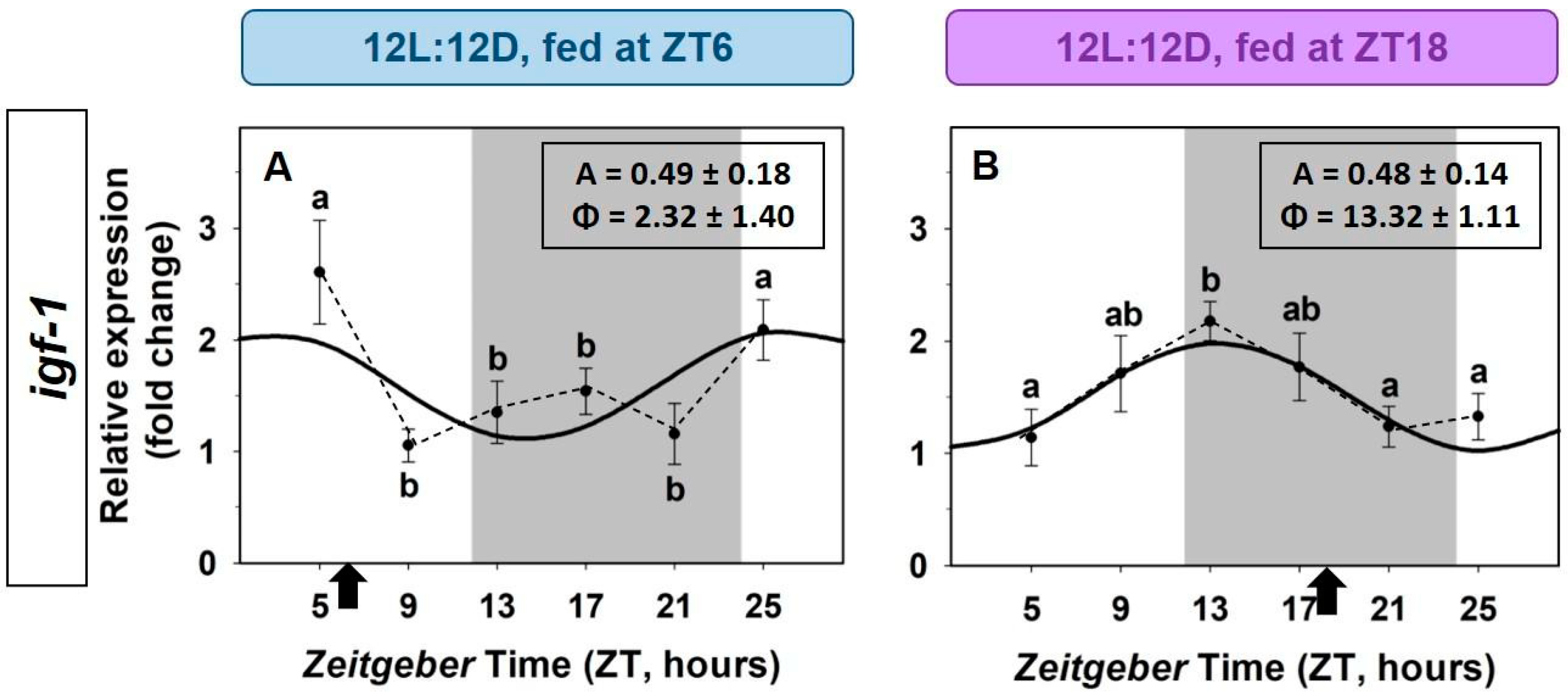
2.4. Entrainment by the Feeding Time of the IGF-1 System in the Liver
2.5. Gene Expression Analysis
2.6. Statistics
3. Results
3.1. Tissue Distribution of the IGF-1 System in Goldfish
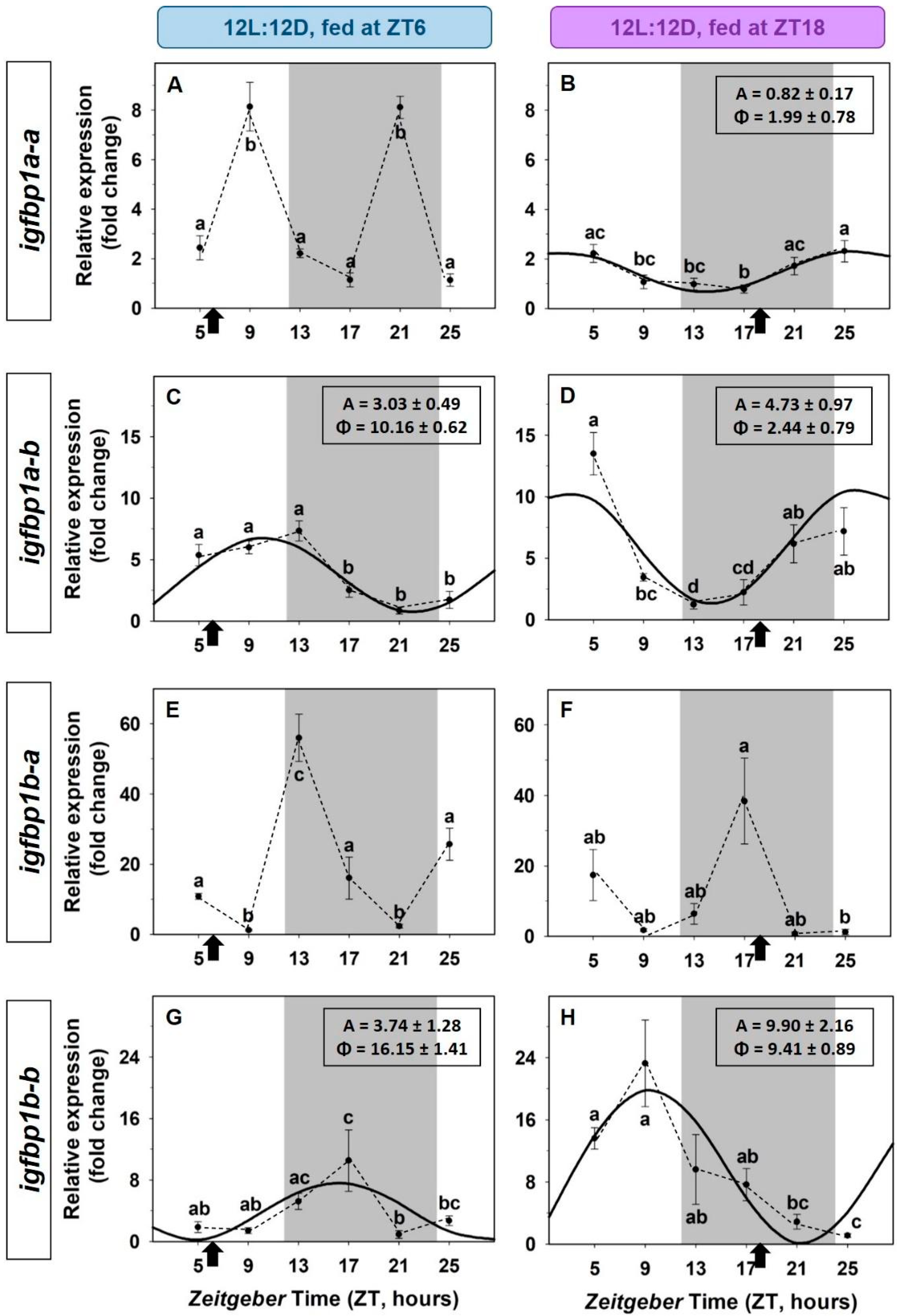
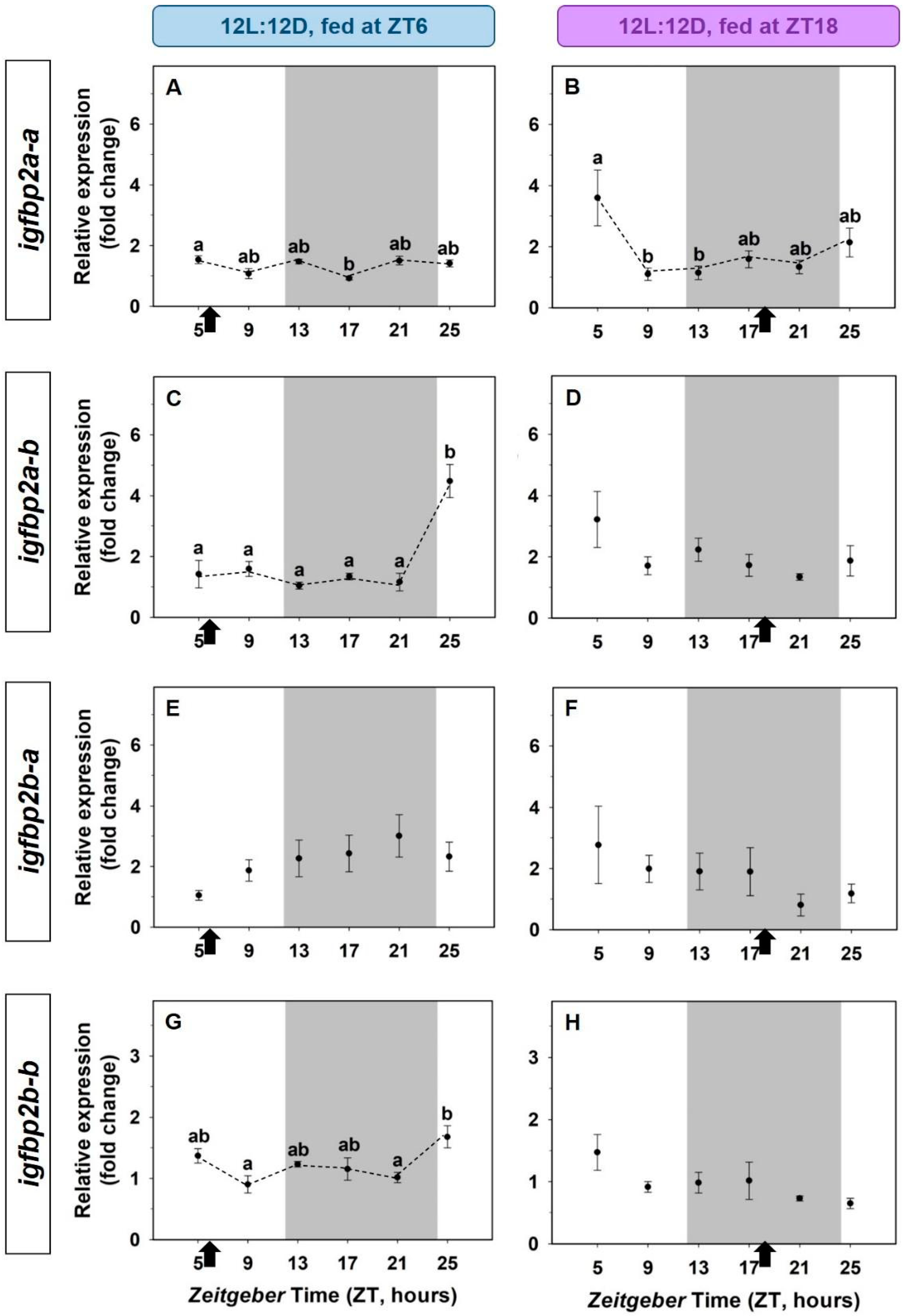
3.2. Daily Variations of the IGF-1 System in the Liver. Light/Dark Cycle and Scheduled Feeding as Zeitgebers
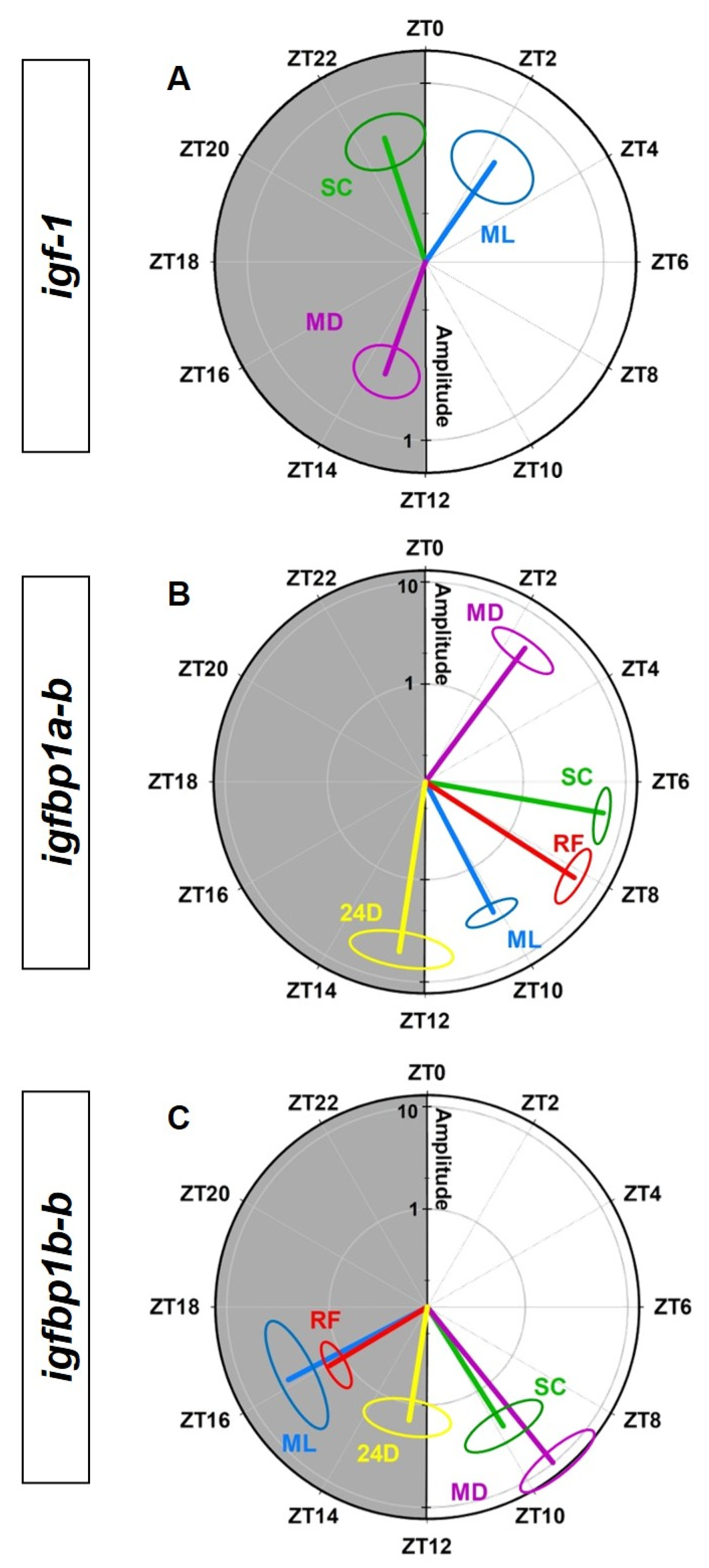
3.3. Entrainment by the Feeding Time of the IGF-1 System in the Liver
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ohlsson, C.; Mohan, S.; Sjögren, K.; Tivesten, Å.; Isgaard, J.; Isaksson, O.; Jansson, J.O.; Svensson, J. The role of liver-derived insulin-like growth factor-I. Endocr. Rev. 2009, 30, 494–535. [Google Scholar] [CrossRef] [PubMed]
- Triantaphyllopoulos, K.A.; Cartas, D.; Miliou, H. Factors influencing GH and IGF-I gene expression on growth in teleost fish: How can aquaculture industry benefit? Rev. Aquac. 2019, 12, 1637–1662. [Google Scholar] [CrossRef]
- Shimizu, M. Insulin-Like Growth Factor-1. In Handbook of Hormones. Comparative Endocrinology for Basic and Clinical Research; Ando, H., Ukena, K., Nagata, S., Eds.; Academic Press: London, UK, 2021; pp. 285–296. ISBN 9780128206492. [Google Scholar]
- Hakuno, F.; Takahashi, S.I. 40 years of IGF1: IGF1 receptor signaling pathways. J. Mol. Endocrinol. 2018, 61, T69–T86. [Google Scholar] [CrossRef] [PubMed]
- Duan, C. Nutritional and developmental roles of insulin-like growth factors between species: A brief history and introduction. J. Nutr. 1998, 128, 306–314. [Google Scholar] [CrossRef]
- Bertucci, J.I.; Blanco, A.M.; Sundarrajan, L.; Rajeswari, J.J.; Velasco, C.; Unniappan, S. Nutrient regulation of endocrine factors influencing feeding and growth in fish. Front. Endocrinol. 2019, 10, 83. [Google Scholar] [CrossRef]
- Beckman, B.R. Perspectives on concordant and discordant relations between insulin-like growth factor 1 (IGF1) and growth in fishes. Gen. Comp. Endocrinol. 2011, 170, 233–252. [Google Scholar] [CrossRef]
- Kamei, H.; Duan, C. Alteration of organ size and allometric scaling by organ-specific targeting of IGF signaling. Gen. Comp. Endocrinol. 2021, 314, 113922. [Google Scholar] [CrossRef] [PubMed]
- Picha, M.E.; Turano, M.J.; Beckman, B.R.; Borski, R.J. Endocrine Biomarkers of Growth and Applications to Aquaculture: A Minireview of Growth Hormone, Insulin-Like Growth Factor (IGF)-I, and IGF-Binding Proteins as Potential Growth Indicators in Fish. N. Am. J. Aquac. 2008, 70, 196–211. [Google Scholar] [CrossRef]
- Caruso, M.A.; Sheridan, M.A. New insights into the signaling system and function of insulin in fish. Gen. Comp. Endocrinol. 2011, 173, 227–247. [Google Scholar] [CrossRef] [PubMed]
- Datuin, J.P.; Ng, K.P.; Hayes, T.B.; Bern, H.A. Effects of glucocorticoids on cartilage growth and response to IGF-I in the tilapia (Oreochromis mossambicus). Gen. Comp. Endocrinol. 2001, 121, 289–294. [Google Scholar] [CrossRef]
- Otteson, D.C.; Cirenza, P.F.; Hitchcock, P.F. Persistent neurogenesis in the teleost retina: Evidence for regulation by the growth-hormone/insulin-like growth factor-I axis. Mech. Dev. 2002, 117, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Canosa, L.F.; Bertucci, J.I. Nutrient regulation of somatic growth in teleost fish. The interaction between somatic growth, feeding and metabolism. Mol. Cell. Endocrinol. 2020, 518, 111029. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Takahashi, E.; Morro, B.; Balseiro, P.; Albalat, A.; Pedrosa, C.; Mackenzie, S.; Nilsen, T.O.; Sveier, H.; Ebbesson, L.O.; et al. Changes in circulating insulin-like growth factor-1 and its binding proteins in yearling rainbow trout during spring under natural and manipulated photoperiods and their relationships with gill Na+, K+-ATPase and body size. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2022, 268, 111205. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.J.; Lee, Y.C.; Tsou, Y.L.; Tseng, Y.C.; Hwang, P.P. Insulin-like growth factor 1 triggers salt secretion machinery in fish under acute salinity stress. J. Endocrinol. 2020, 246, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, K.; Kazeto, Y.; Ozaki, Y.; Izumida, D.; Hotta, T.; Soyano, K.; Gen, K. Insulin-like growth factors 1 and 2 regulate gene expression and enzymatic activity of cyp17a1 in ovarian follicles of the yellowtail, Seriola quinqueradiata. Heliyon 2020, 6, e04181. [Google Scholar] [CrossRef] [PubMed]
- Reinecke, M. Insulin-like growth factors and fish reproduction. Biol. Reprod. 2010, 82, 656–661. [Google Scholar] [CrossRef]
- LeRoith, D.; Holly, J.M.P.; Forbes, B.E. Insulin-like growth factors: Ligands, binding proteins, and receptors. Mol. Metab. 2021, 52, 101245. [Google Scholar] [CrossRef]
- Wood, A.W.; Duan, C.; Bern, H.A. Insulin-like growth factor signaling in fish. Int. Rev. Cytol. 2005, 243, 215–285. [Google Scholar] [CrossRef]
- Hoegg, S.; Brinkmann, H.; Taylor, J.S.; Meyer, A. Phylogenetic timing of the fish-specific genome duplication correlates with the diversification of teleost fish. J. Mol. Evol. 2004, 59, 190–203. [Google Scholar] [CrossRef]
- Macqueen, D.J.; Johnston, I.A. A well-constrained estimate for the timing of the salmonid whole genome duplication reveals major decoupling from species diversification. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132881. [Google Scholar] [CrossRef]
- Yang, L.; Sado, T.; Vincent Hirt, M.; Pasco-Viel, E.; Arunachalam, M.; Li, J.; Wang, X.; Freyhof, J.; Saitoh, K.; Simons, A.M.; et al. Phylogeny and polyploidy: Resolving the classification of cyprinine fishes (Teleostei: Cypriniformes). Mol. Phylogenet. Evol. 2015, 85, 97–116. [Google Scholar] [CrossRef] [PubMed]
- Hitchcock, P.F.; Otteson, D.C.; Cirenza, P.F. Expression of the insulin receptor in the retina of the goldfish. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2125–2129. [Google Scholar]
- Kuang, Y.M.; Li, W.S.; Lin, H.R. Molecular cloning and mRNA profile of insulin-like growth factor type 1 receptor in orange-spotted grouper, Epinephelus coioides. Acta Biochim. Biophys. Sin. 2005, 37, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Nakao, N.; Tanaka, M.; Higashimoto, Y.; Nakashima, K. Molecular cloning, identification and characterization of four distinct receptor subtypes for insulin and IGF-I in Japanese flounder, Paralichthys olivaceus. J. Endocrinol. 2002, 173, 365–375. [Google Scholar] [CrossRef]
- García de la Serrana, D.; Macqueen, D.J. Insulin-like growth factor-binding proteins of teleost fishes. Front. Endocrinol. 2018, 9, 80. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Dickhoff, W.W. Circulating insulin-like growth factor binding proteins in fish: Their identities and physiological regulation. Gen. Comp. Endocrinol. 2017, 252, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.I.; Clemmons, D.R. Insulin-like growth factors and their binding proteins: Biological actions. Endocr. Rev. 1995, 16, 3–34. [Google Scholar] [CrossRef] [PubMed]
- Allard, J.B.; Duan, C. IGF-binding proteins: Why do they exist and why are there so many? Front. Endocrinol. 2018, 9, 117. [Google Scholar] [CrossRef]
- Bach, L.A. 40 years of IGF1: IGF-binding proteins. J. Mol. Endocrinol. 2018, 61, T11–T28. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, Z.; Dong, H.; Yan, F. Insulin-like growth factor-binding protein-1 (IGFBP-1) in goldfish, Carassius auratus: Molecular cloning, tissue expression, and mRNA expression responses to periprandial changes and cadmium exposure. Fish Physiol. Biochem. 2016, 42, 1043–1052. [Google Scholar] [CrossRef]
- Chen, W.; Li, W.; Zhang, Z.; Jiang, X.; Li, M. Cloning, molecular characterization and expression analysis of insulin-like growth factor binding protein-2 (IGFBP-2) cDNA in goldfish, Carassius auratus. Fish Physiol. Biochem. 2014, 40, 1669–1681. [Google Scholar] [CrossRef]
- Isorna, E.; de Pedro, N.; Valenciano, A.I.; Alonso-Gómez, Á.L.; Delgado, M.J. Interplay between the endocrine and circadian systems in fishes. J. Endocrinol. 2017, 232, R141–R159. [Google Scholar] [CrossRef] [PubMed]
- Ayson, F.G.; Takemura, A. Daily expression patterns for mRNAs of GH, PRL, SL, IGF-I and IGF-II in juvenile rabbitfish, Siganus guttatus, during 24-h light and dark cycles. Gen. Comp. Endocrinol. 2006, 149, 261–268. [Google Scholar] [CrossRef]
- Costa, L.S.; Rosa, P.V.; Fortes-Silva, R.; Sánchez-Vázquez, F.J.; López-Olmeda, J.F. Daily rhythms of the expression of genes from the somatotropic axis: The influence on tilapia (Oreochromis niloticus) of feeding and growth hormone administration at different times. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2016, 181, 27–34. [Google Scholar] [CrossRef]
- López-Olmeda, J.F.; Pujante, I.M.; Costa, L.S.; Galal-Khallaf, A.; Mancera, J.M.; Sánchez-Vázquez, F.J. Daily rhythms in the somatotropic axis of Senegalese sole (Solea senegalensis): The time of day influences the response to GH administration. Chronobiol. Int. 2016, 33, 257–267. [Google Scholar] [CrossRef]
- Metón, I.; Caseras, A.; Cantó, E.; Fernández, F.; Baanante, I.V. Liver insulin-like growth factor-I mRNA is not affected by diet composition or ration size but shows diurnal variations in regularly-fed gilthead sea bream (Sparus aurata). J. Nutr. 2000, 130, 757–760. [Google Scholar] [CrossRef]
- Paredes, J.F.; López-Olmeda, J.F.; Martínez, F.J.; Sánchez-Vázquez, F.J. Daily rhythms of lipid metabolic gene expression in zebra fish liver: Response to light/dark and feeding cycles. Chronobiol. Int. 2015, 32, 1438–1448. [Google Scholar] [CrossRef]
- Galal-Khallaf, A.; Mohammed-Geba, K.; Yúfera, M.; Martínez-Rodríguez, G.; Mancera, J.M.; López-Olmeda, J.F. Daily rhythms in endocrine factors of the somatotropic axis and their receptors in gilthead sea bream (Sparus aurata) larvae. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 250, 110793. [Google Scholar] [CrossRef] [PubMed]
- Feliciano, A.; Vivas, Y.; De Pedro, N.; Delgado, M.J.; Velarde, E.; Isorna, E. Feeding time synchronizes clock gene rhythmic expression in brain and liver of goldfish (Carassius auratus). J. Biol. Rhythms 2011, 26, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bretaño, A.; Alonso-Gómez, Á.L.; Delgado, M.J.; Isorna, E. The liver of goldfish as a component of the circadian system: Integrating a network of signals. Gen. Comp. Endocrinol. 2015, 221, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Velarde, E.; Haque, R.; Iuvone, P.M.; Azpeleta, C.; Alonso-Gómez, Á.L.; Delgado, M.J. Circadian clock genes of goldfish, Carassius auratus: cDNA cloning and rhythmic expression of Period and Cryptochrome transcripts in retina, liver, and gut. J. Biol. Rhythms 2009, 24, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Saiz, N.; Gómez-Boronat, M.; De Pedro, N.; Delgado, M.J.; Isorna, E. The lack of light-dark and feeding-fasting cycles alters temporal events in the goldfish (Carassius auratus) stress axis. Animals 2021, 11, 669. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Boronat, M.; Sáiz, N.; Delgado, M.J.; de Pedro, N.; Isorna, E. Time-Lag in Feeding Schedule Acts as a Stressor That Alters Circadian Oscillators in Goldfish. Front. Physiol. 2018, 9, 1749. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Delgado, M.J.; Alonso-Gómez, Á.L.; Gancedo, B.; De Pedro, N.; Valenciano, A.I.; Alonso-Bedate, M. Serotonin N-Acetyltransferase (NAT) activity and melatonin levels in the frog retina are not correlated during the seasonal cycle. Gen. Comp. Endocrinol. 1993, 92, 143–150. [Google Scholar] [CrossRef]
- Duggleby, R.G. A Nonlinear Regression Program for Small Computers. Anal. Biochem. 1981, 110, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.; Plisetskaya, E.M.M. Nutritional regulation of insulin-like growth factor-I mRNA expression in salmon tissues. J. Endocrinol. 1993, 139, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Pierce, A.L.; Dickey, J.T.; Larsen, D.A.; Fukada, H.; Swanson, P.; Dickhoff, W.W. A quantitative real-time RT-PCR assay for salmon IGF-I mRNA, and its application in the study of GH regulation of IGF-I gene expression in primary culture of salmon hepatocytes. Gen. Comp. Endocrinol. 2004, 135, 401–411. [Google Scholar] [CrossRef]
- Caelers, A.; Berishvili, G.; Meli, M.L.; Eppler, E.; Reinecke, M. Establishment of a real-time RT-PCR for the determination of absolute amounts of IGF-I and IGF-II gene expression in liver and extrahepatic sites of the tilapia. Gen. Comp. Endocrinol. 2004, 137, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Maures, T.; Chan, S.J.; Xu, B.; Sun, H.; Ding, J.; Duan, C. Structural, biochemical, and expression analysis of two distinct insulin-like growth factor I receptors and their ligands in zebrafish. Endocrinology 2002, 143, 1858–1871. [Google Scholar] [CrossRef] [PubMed]
- Vong, Q.P.; Chan, K.M.; Cheng, C.H.K. Quantification of common carp (Cyprinus carpio) IGF-I and IGF-II mRNA by real-time PCR: Differential regulation of expression by GH. J. Endocrinol. 2003, 178, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Aegerter, S.; Jalabert, B.; Bobe, J. Messenger RNA Stockpile of Cyclin B, Insulin-Like Growth Factor I, Insulin-Like Growth Factor II, Insulin-Like Growth Factor Receptor Ib, and p53 in the Rainbow Trout Oocyte in Relation with Developmental Competence. Mol. Reprod. Dev. 2004, 67, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Yang, Y.; Ning, C.; Wang, Y.; Hu, J.; Zhang, M.; Kuang, S.; Sun, Y.; Li, Y.; Zhang, Y.; et al. Identification and characteristics of insulin-like growth factor system in the brain, liver, and gonad during development of a seasonal breeding teleost, Pampus argenteus. Gen. Comp. Endocrinol. 2021, 300, 113645. [Google Scholar] [CrossRef] [PubMed]
- Alzaid, A.; Martin, S.A.M.; Macqueen, D.J. The complete salmonid IGF-IR gene repertoire and its transcriptional response to disease. Sci. Rep. 2016, 6, 34806. [Google Scholar] [CrossRef] [PubMed]
- LeRoith, D.; Werner, H.; Beitner-Johnson, D.; Roberts, C.T. Molecular and cellular aspects of the Insulin-like Growth Factor I Receptor. Endocr. Rev. 1995, 16, 143–163. [Google Scholar] [CrossRef]
- Mancera, J.M.; McCormick, S.D. Osmoregulatory actions of the GH/IGF axis in non-salmonid teleosts. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1998, 121, 43–48. [Google Scholar] [CrossRef]
- Macqueen, D.J.; Garcia De La Serrana, D.; Johnston, I.A. Evolution of ancient functions in the vertebrate insulin-like growth factor system uncovered by study of duplicated salmonid fish genomes. Mol. Biol. Evol. 2013, 30, 1060–1076. [Google Scholar] [CrossRef] [PubMed]
- Kamei, H.; Lu, L.; Jiao, S.; Li, Y.; Gyrup, C.; Laursen, L.S.; Oxvig, C.; Zhou, J.; Duan, C. Duplication and diversification of the hypoxia-inducible IGFBP-1 gene in Zebrafish. PLoS ONE 2008, 3, e3091. [Google Scholar] [CrossRef]
- Gómez-Boronat, M.; De Pedro, N.; Alonso-Gómez, Á.L.; Delgado, M.J.; Isorna, E. Nuclear Receptors (PPARs, REV-ERBs, RORs) and Clock Gene Rhythms in Goldfish (Carassius auratus) Are Differently Regulated in Hypothalamus and Liver. Front. Physiol. 2022, 13, 903799. [Google Scholar] [CrossRef] [PubMed]
- Pérez, L.; Ortiz-Delgado, J.B.; Manchado, M. Molecular characterization and transcriptional regulation by GH and GnRH of insulin-like growth factors I and II in white seabream (Diplodus sargus). Gene 2016, 578, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Vong, Q.P.; Chan, K.M.; Leung, K.; Cheng, C.H.K. Common carp insulin-like growth factor-I gene: Complete nucleotide sequence and functional characterization of the 5′-flanking region. Gene 2003, 322, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Bozek, K.; Relógio, A.; Kielbasa, S.M.; Heine, M.; Dame, C.; Kramer, A.; Herzel, H. Regulation of Clock-Controlled Genes in Mammals. PLoS ONE 2009, 4, e4882. [Google Scholar] [CrossRef] [PubMed]
- Nisembaum, L.G.; Velarde, E.; Tinoco, A.B.; Azpeleta, C.; De Pedro, N.; Alonso-Gómez, A.L.; Delgado, M.J.; Isorna, E. Light-dark cycle and feeding time differentially entrains the gut molecular clock of the goldfish (Carassius auratus). Chronobiol. Int. 2012, 29, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Zou, Z.; Zhu, J.; Xiao, W.; Li, D.; Yu, J.; Chen, B. Effects of different photoperiods on growth performance, daily rhythm of growth axis-related genes, and hormones in Nile tilapia (Oreochromis niloticus). Aquaculture 2022, 553, 738071. [Google Scholar] [CrossRef]
- Amaral, I.P.G.; Johnston, I.A. Circadian expression of clock and putative clock-controlled genes in skeletal muscle of the zebrafish. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R193–R206. [Google Scholar] [CrossRef] [PubMed]
- Vera, L.M.; Negrini, P.; Zagatti, C.; Frigato, E.; Sánchez-Vázquez, F.J.; Bertolucci, C. Light and feeding entrainment of the molecular circadian clock in a marine teleost (Sparus aurata). Chronobiol. Int. 2013, 30, 649–661. [Google Scholar] [CrossRef] [PubMed]
- López-Olmeda, J.F.; Tartaglione, E.V.; De La Iglesia, H.O.; Sánchez-Vázquez, F.J. Feeding entrainment of food-anticipatory activity and per1 expression in the brain and liver of zebrafish under different lighting and feeding conditions. Chronobiol. Int. 2010, 27, 1380–1400. [Google Scholar] [CrossRef] [PubMed]
- Breves, J.P.; Tipsmark, C.K.; Stough, B.A.; Seale, A.P.; Flack, B.R.; Moorman, B.P.; Lerner, D.T.; Grau, E.G. Nutritional status and growth hormone regulate insulin-like growth factor binding protein (igfbp) transcripts in Mozambique tilapia. Gen. Comp. Endocrinol. 2014, 207, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Breves, J.P.; Phipps-Costin, S.K.; Fujimoto, C.K.; Einarsdottir, I.E.; Regish, A.M.; Björnsson, B.T.; McCormick, S.D. Hepatic insulin-like growth-factor binding protein (igfbp) responses to food restriction in Atlantic salmon smolts. Gen. Comp. Endocrinol. 2016, 233, 79–87. [Google Scholar] [CrossRef]
- Strobel, J.S.; Hack, N.L.; Label, K.T.; Cordova, K.L.; Bersin, T.V.; Journey, M.L.; Beckman, B.R.; Lema, S.C. Effects of food deprivation on plasma insulin-like growth factor-1 (Igf1) and Igf binding protein (Igfbp) gene transcription in juvenile cabezon (Scorpaenichthys marmoratus). Gen. Comp. Endocrinol. 2020, 286, 113319. [Google Scholar] [CrossRef] [PubMed]
- Reinecke, M. Influences of the environment on the endocrine and paracrine fish growth hormone-insulin-like growth factor-I system. J. Fish Biol. 2010, 76, 1233–1254. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alonso-Gómez, A.; Madera, D.; Alonso-Gómez, Á.L.; Valenciano, A.I.; Delgado, M.J. Daily Rhythms in the IGF-1 System in the Liver of Goldfish and Their Synchronization to Light/Dark Cycle and Feeding Time. Animals 2022, 12, 3371. https://doi.org/10.3390/ani12233371
Alonso-Gómez A, Madera D, Alonso-Gómez ÁL, Valenciano AI, Delgado MJ. Daily Rhythms in the IGF-1 System in the Liver of Goldfish and Their Synchronization to Light/Dark Cycle and Feeding Time. Animals. 2022; 12(23):3371. https://doi.org/10.3390/ani12233371
Chicago/Turabian StyleAlonso-Gómez, Aitana, Diego Madera, Ángel Luis Alonso-Gómez, Ana Isabel Valenciano, and María Jesús Delgado. 2022. "Daily Rhythms in the IGF-1 System in the Liver of Goldfish and Their Synchronization to Light/Dark Cycle and Feeding Time" Animals 12, no. 23: 3371. https://doi.org/10.3390/ani12233371
APA StyleAlonso-Gómez, A., Madera, D., Alonso-Gómez, Á. L., Valenciano, A. I., & Delgado, M. J. (2022). Daily Rhythms in the IGF-1 System in the Liver of Goldfish and Their Synchronization to Light/Dark Cycle and Feeding Time. Animals, 12(23), 3371. https://doi.org/10.3390/ani12233371