
Seasonal Trophic Ecology and Diet Shift in the Common Sole Solea solea in the Central Adriatic Sea

 , , and
, , and 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
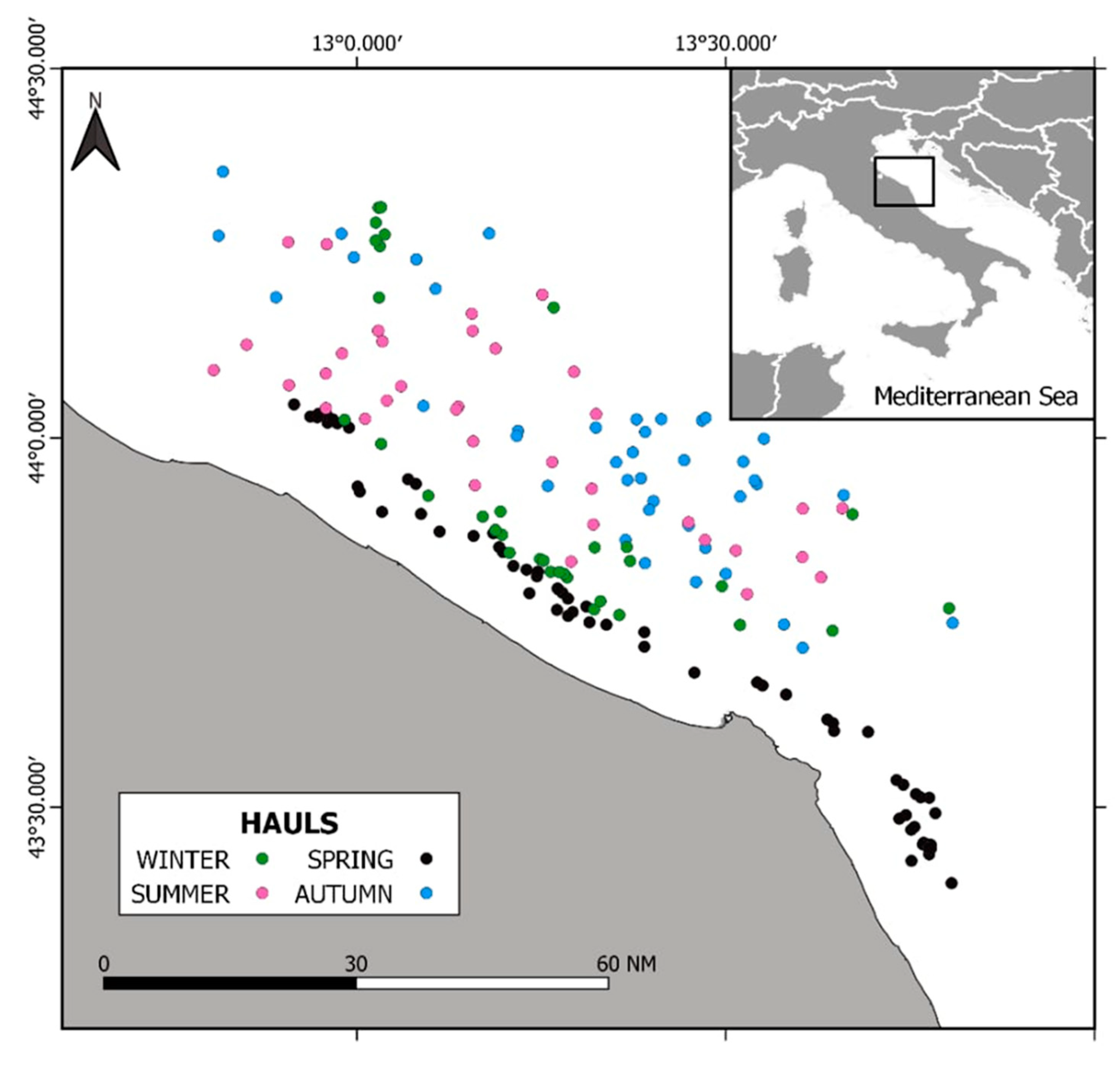
2.1. Study Area
2.2. Samples Collection and Processing
2.3. Stable Isotopes Analysis
2.4. Data Treatment
2.4.1. Biological Indices, Stomach Contents, and Trophic Diversity
2.4.2. Stable Isotopes Data
3. Results
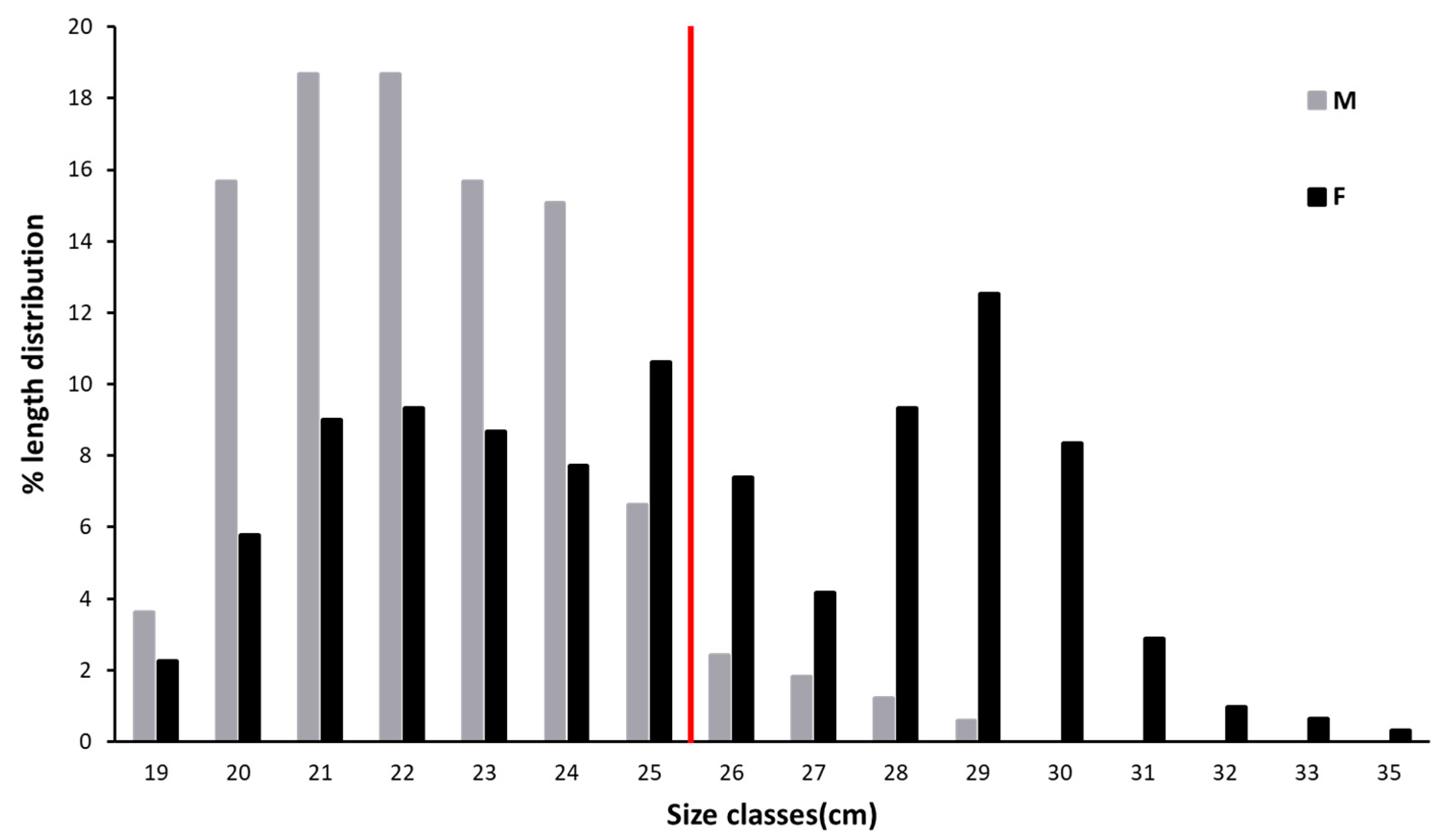
3.1. Population Structure
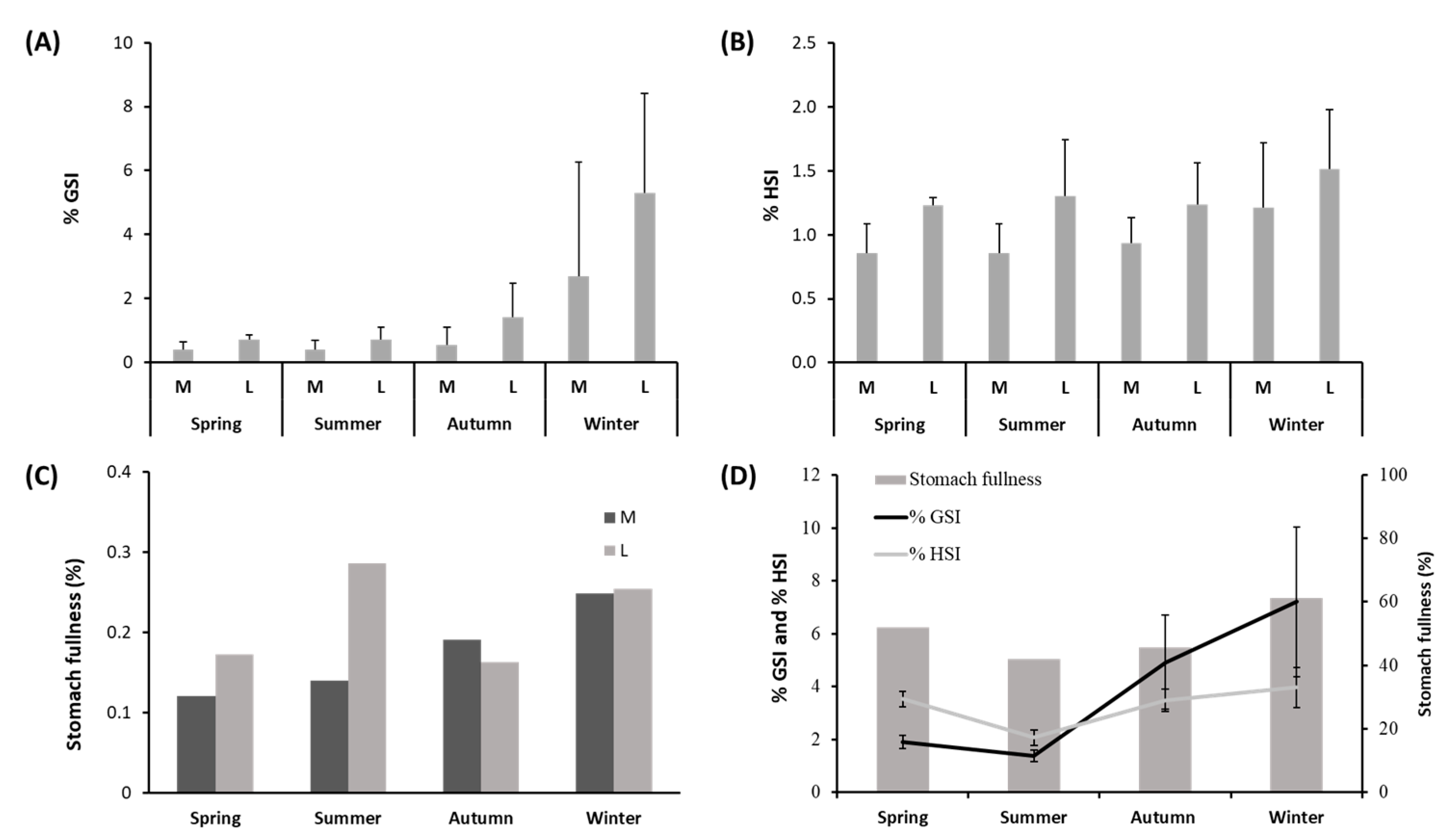
3.2. Variations in Condition Indices and Fullness
3.3. Seasonal Variations in Diet Composition
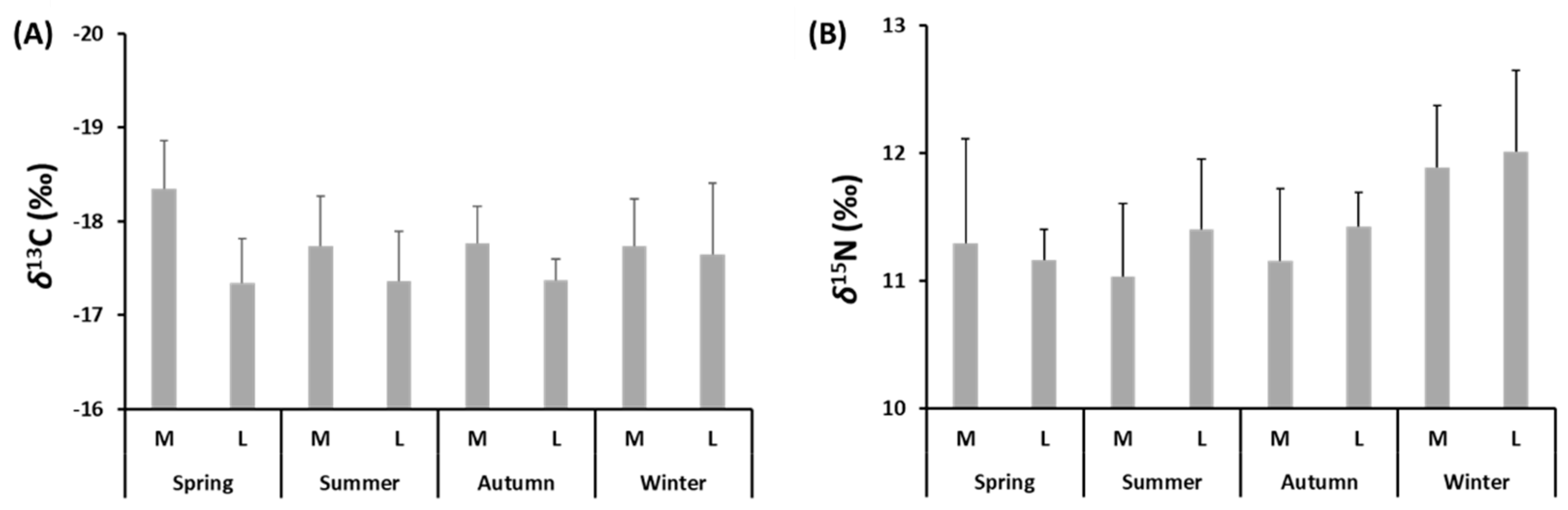
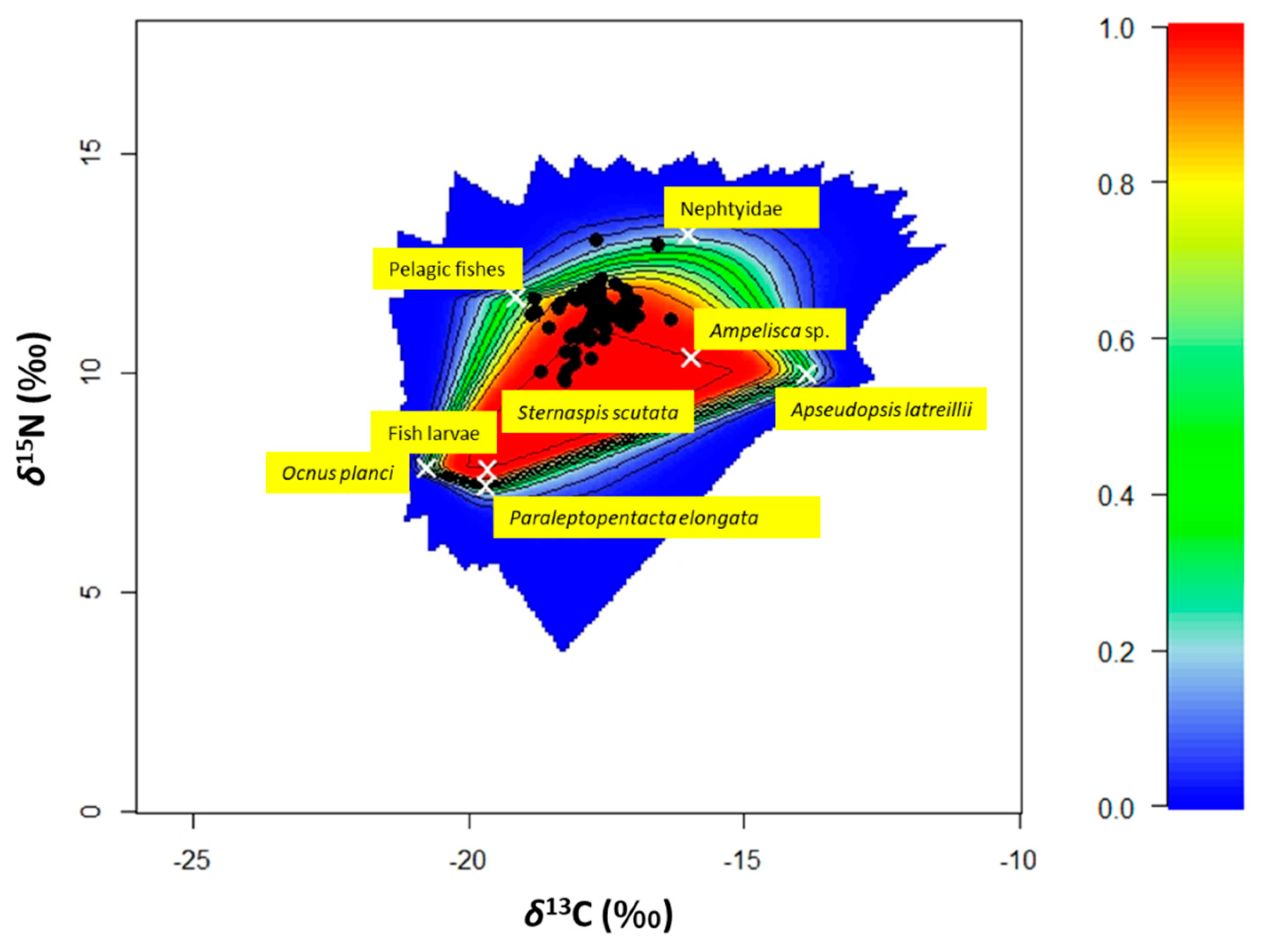
3.4. Seasonal Variations in Stable Isotope Composition
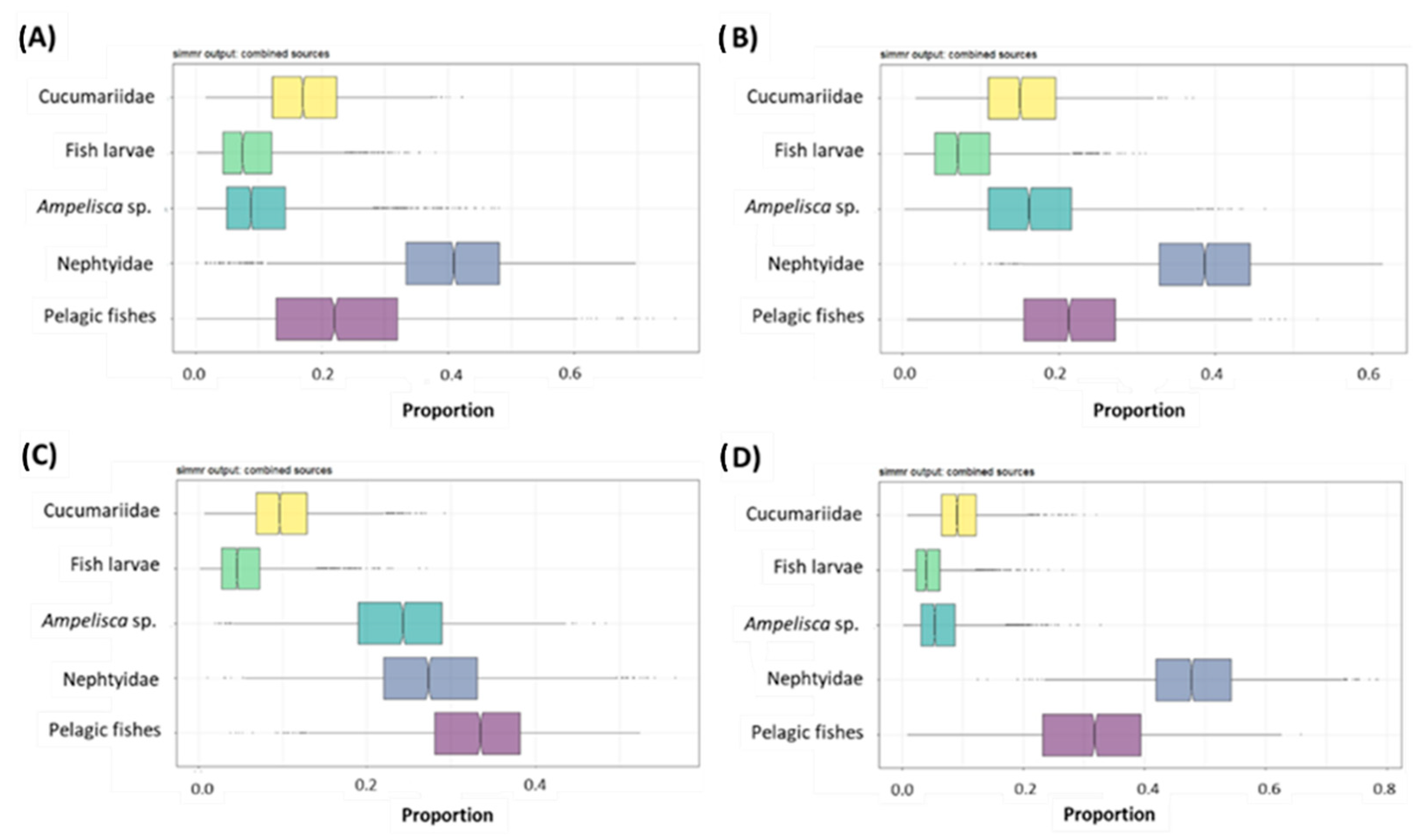
3.5. Bayesian Mixing Models and SEAc
4. Discussion
4.1. Seasonal Changes in the Feeding Ecology of Solea solea
4.2. Biological Traits of Common Sole in the Adriatic Sea and Correlation with Feeding Habits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bjørndal, T.; Guillen, J.; Imsland, A. The potential of aquaculture sole production in Europe: Production costs and markets. Aquac. Econ. Manag. 2016, 20, 109–129. [Google Scholar] [CrossRef]
- Jayasinghe, R.P.P.K.; Amarasinghe, U.S.; Newton, A. Evaluation of status of commercial fish stocks in European marine subareas using mean trophic levels of fish landings and spawning stock biomass. Ocean. Coast. Manag. 2017, 143, 154–163. [Google Scholar] [CrossRef][Green Version]
- Quéro, J.-C.; Desoutter, M.; Lagardère, F.S. Fishes of the North-Eastern Atlantic and the Mediterranean; Whitehead, P.J.P., Bauchot, M.L., Hureau, J.-C., Nielsen, J., Tortonese, E., Eds.; UNESCO: Paris, France, 1986; Volume 3. [Google Scholar]
- Fischer, W.; Schneider, M.; Bauchot, M.-L. Fiches FAO d’identification des especes pour les besoins de la pêche. In Méditerranée et Mer Noire (Zone De Pêche 37); FAO: Rome, Italy, 1987; Volume I–II. [Google Scholar]
- Jardas, I.; Pallaoro, A.; Vrgoc, N.; Jukić-Peladić, S.; Dadic, V. Red Book of Sea Fishes of Croatia; State Institute for Nature Protection: Zagreb, Croatia, 2008. [Google Scholar]
- Cerim, H.; Ateş, C. Reproductive biology of female common sole, Solea solea (Linnaeus, 1758) in the southern Aegean Sea. Acta Biol. Turc. 2019, 32, 143–148. [Google Scholar]
- Teixeira, C.M.; Cabral, H.N. Comparative analysis of the diet, growth and reproduction of the soles, Solea solea and Solea senegalensis, occurring in sympatry along the Portuguese coast. J. Mar. Biol. Assoc. UK 2010, 90, 995–1003. [Google Scholar] [CrossRef]
- Daban, I.B.; Arslan İHsanoĞLu, M.; İŞMen, A.; YiĞIn, C.Ç. Age, Growth, and Reproduction of Common Sole, Solea solea (Linnaeus, 1758) in the Sea of Marmara. Acta Aquat. Turc. 2021, 17, 395–408. [Google Scholar] [CrossRef]
- Bromley, P.J. The use of market sampling to generate maturity ogives and to investigate growth, sexual dimorphism and reproductive strategy in central and south-western North Sea sole (Solea solea L.). ICES J. Mar. Sci. 2003, 60, 52–65. [Google Scholar] [CrossRef]
- Türkmen, M. Investigation of Some Population Parameters of Common Sole, Solea solea (L., 1758) from Iskenderun Bay. Turk. J. Vet. Anim. Sci. 2003, 27, 317–323. [Google Scholar]
- Froglia, C.; Giannetti, G. Growth of common sole Solea vulgaris Quensel in the Adriatic Sea (Osteichthyes, Soleidae). Rapp. Et Procés Verbaux Des Réunions De La Comm. Int. Pour I’Exploration De La Mer Méditerranée 1985, 29, 91–93. [Google Scholar]
- Froglia, C.; Giannetti, G. Remarks on rings formation in otoliths of Solea vulgaris and other flatfishes from the Adriatic sea. FAO Fish. Rep. 1986, 345, 121–122. [Google Scholar]
- Lagardère, J.P. Feeding ecology and daily food consumption of common sole, Solea vulgaris Quensel, juveniles on the French Atlantic coast. J. Fish Biol. 1987, 30, 91–104. [Google Scholar] [CrossRef]
- van de Wolfshaar, K.E.; Barbut, L.; Lacroix, G. From spawning to first-year recruitment: The fate of juvenile sole growth and survival under future climate conditions in the North Sea. ICES J. Mar. Sci. 2021, 79, 495–505. [Google Scholar] [CrossRef]
- Tortonese, E. Osteichthyes (Bony Fish) Part Two. Fauna Italy 1975, 11, 1–636. [Google Scholar]
- Piccinetti, C.; Giovanardi, O. Donnees biologiques sur Solea vulgaris Quensel en Adriatique. FAO Fish. Rep. 2012, 290, 117–121. [Google Scholar]
- Jardas, I. Jadranska Ihtiofauna; Školska knjiga: Zagreb, Croatia, 1996. [Google Scholar]
- Vallisneri, M.; Piccinetti, C.; Stagni, A.M.; Colombari, A.; Tinti, F. Dinamica di popolazione, accrescimento, riproduzione di Solea vulgaris (Quensel 1806) nell’alto Adriatico. Biol. Mar. Mediterr. 2000, 7, 65–70. [Google Scholar]
- Pagotto, G.; Piccinetti, C. Censimento della popolazione di Solea vulgaris Quensel 1814. in Adriatico mediante marcatura. In Proceedings of the Atti del I Seminario Italiano sui Censimenti Faunistici, Metodi e applicabilità alla gestione territoriale, Urbino, Italy, 21–22 September 1982; pp. 345–359. [Google Scholar]
- Pagotto, G.; Piccinetti, C.; Specchi, M. Premières résultats des campagnes de marquage des soles en Adriatique: Déplacements. Rapp. Comm. Int. Mer Médit. 1979, 25/26, 111–112. [Google Scholar]
- Braber, L.; de Groot, S.J. On the morphology of the alimentary tract of flatfishes (Pleuronectiformes). J. Fish Biol. 1973, 5, 147–153. [Google Scholar] [CrossRef]
- Cabral, H.; Costa, M.J. Differential use of nursery areas within the Tagus estuary by sympatric soles, Solea solea and Solea senegalensis. Environ. Biol. Fishes 1999, 56, 389–397. [Google Scholar] [CrossRef]
- De Groot, S.J. On the interrelationships between morphology of the alimentary tract, food and feeding behaviour in flatfishes (Pisces Pleuronectiformes). Neth. J. Sea Res. 1971, 5, 121–196. [Google Scholar] [CrossRef]
- Molinero, A.; Flos, R. Influence of sex and age on the feeding habits of the common sole Solea solea. Mar. Biol. 1991, 111, 493–501. [Google Scholar] [CrossRef]
- Molinero, A.; Flos, R. Influence of season on the feeding habits of the common sole Solea solea. Mar. Biol. 1992, 113, 499–507. [Google Scholar] [CrossRef]
- FAO-GFCM. The State of Mediterranean and Black Sea Fisheries; FAO: Rome, Italy, 2018; p. 172. [Google Scholar]
- Pellini, G.; Gomiero, A.; Fortibuoni, T.; Ferrà, C.; Grati, F.; Tassetti, A.N.; Polidori, P.; Fabi, G.; Scarcella, G. Characterization of microplastic litter in the gastrointestinal tract of Solea solea from the Adriatic Sea. Environ. Pollut. 2018, 234, 943–952. [Google Scholar] [CrossRef] [PubMed]
- FAO. Fishery and Aquaculture Statistics. Gfcm Capture Production 1970–2019 (FishstatJ). Updated 2021, 2021.
- Scarcella, G.; Grati, F.; Raicevich, S.; Russo, T.; Gramolini, R.; Scott, R.D.; Polidori, P.; Domenichetti, F.; Bolognini, L.; Giovanardi, O.; et al. Common sole in the northern and central Adriatic Sea: Spatial management scenarios to rebuild the stock. J. Sea Res. 2014, 89, 12–22. [Google Scholar] [CrossRef]
- Fanelli, E.; Badalamenti, F.; D’Anna, G.; Pipitone, C. Diet and trophic level of scaldfish Arnoglossus laterna in the southern Tyrrhenian Sea (western Mediterranean): Contrasting trawled versus untrawled areas. J. Mar. Biol. Assoc. UK 2009, 89, 817–828. [Google Scholar] [CrossRef]
- Fanelli, E.; Badalamenti, F.; D’Anna, G.; Pipitone, C.; Romano, C. Trophodynamic effects of trawling on the feeding ecology of pandora, Pagellus erythrinus, off the northern Sicily coast (Mediterranean Sea). Mar. Freshw. Res. 2010, 61, 408–417. [Google Scholar] [CrossRef]
- Lowerre-Barbieri, S.K.; Ganias, K.; Saborido-Rey, F.; Murua, H.; Hunter, J.R. Reproductive Timing in Marine Fishes: Variability, Temporal Scales, and Methods. Mar. Coast. Fish. 2011, 3, 71–91. [Google Scholar] [CrossRef]
- Papiol, V.; Cartes, J.E.; Fanelli, E. The role of food availability in regulating the feeding dynamics and reproductive cycles of bathyal benthopelagic fish in the northwest Mediterranean slope. Limnol. Oceanogr. 2014, 59, 1779–1794. [Google Scholar] [CrossRef]
- Rijnsdorp, A.D. The mechanism of energy allocation over reproduction and somatic growth in female North Sea plaice, Pleuronectes platessa L. Neth. J. Sea Res. 1990, 25, 279–289. [Google Scholar] [CrossRef]
- Tomiyama, T.; Kurita, Y. Seasonal and spatial variations in prey utilization and condition of a piscivorous flatfish Paralichthys olivaceus. Aquat. Biol. 2011, 11, 279–288. [Google Scholar] [CrossRef]
- Hyslop, E. Stomach contents analysis—A review of methods and their application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef]
- Peterson, B.J.; Fry, B. Stable isotopes in ecosystem studies. Annu. Rev. Ecol. Syst. 1987, 18, 293–320. [Google Scholar] [CrossRef]
- Macdonald, J.S.; Waiwood, K.G. Rates of Digestion of Different Prey in Atlantic Cod (Gadus morhua), Ocean Pout (Macrozoarces americanus), Winter Flounder (Pseudopleuronectes amerkanus), and American Plaice (Hippoglossoides platessoides). J. Fish. Aquat. Sci. 1982, 39, 651–659. [Google Scholar] [CrossRef]
- Fanelli, E.; Badalamenti, F.; D’Anna, G.; Pipitone, C.; Riginella, E.; Azzurro, E. Food partitioning and diet temporal variation in two coexisting sparids, Pagellus erythrinus and Pagellus acarne. J. Fish Biol. 2011, 78, 869–900. [Google Scholar] [CrossRef] [PubMed]
- Pinnegar, J.K.; Polunin, N.V.C. Differential fractionation of δ13C and δ15N among fish tissues: Implications for the study of trophic interactions. Funct. Ecol. 1999, 13, 225–231. [Google Scholar] [CrossRef]
- Bautista-Vega, A.A.; Letourneur, Y.; Harmelin-Vivien, M.; Salen-Picard, C. Difference in diet and size-related trophic level in two sympatric fish species, the red mullets Mullus barbatus and Mullus surmuletus, in the Gulf of Lions (north-west Mediterranean Sea). J. Fish Biol. 2008, 73, 2402–2420. [Google Scholar] [CrossRef]
- Fanelli, E.; Cartes, J.E.; Rumolo, P.; Sprovieri, M. Food-web structure and trophodynamics of mesopelagic–suprabenthic bathyal macrofauna of the Algerian Basin based on stable isotopes of carbon and nitrogen. Deep Sea Res. Part I Oceanogr. Res. Pap. 2009, 56, 1504–1520. [Google Scholar] [CrossRef]
- Fanelli, E.; Cartes, J.E.; Papiol, V.; Rumolo, P.; Sprovieri, M. Effects of preservation on the δ13C and δ15N values of deep sea macrofauna. J. Exp. Mar. Biol. Ecol. 2010, 395, 93–97. [Google Scholar] [CrossRef]
- Curzi, P.V.; Giordani, P. Geomorfologia e Sedimentologia il “L’eutrofizzazione nel Mar Adriatico”. In Proceedings of the Atti del Convegno Nazionale di Ancona, Ancona, Italy; 1989; pp. 32–43. [Google Scholar]
- Lucchetti, A.; Sala, A. Impact and performance of Mediterranean fishing gear by side-scan sonar technology. Can. J. Fish. Aquat. Sci. 2012, 69, 1806–1816. [Google Scholar] [CrossRef]
- Grati, F.; Scarcella, G.; Polidori, P.; Domenichetti, F.; Bolognini, L.; Gramolini, R.; Vasapollo, C.; Giovanardi, O.; Raicevich, S.; Celić, I.; et al. Multi-annual investigation of the spatial distributions of juvenile and adult sole (Solea solea L.) in the Adriatic Sea (northern Mediterranean). J. Sea Res. 2013, 84, 122–132. [Google Scholar] [CrossRef]
- Carbonara, P.; Porcu, C.; Donnaloia, M.; Pesci, P.; Sion, L.; Spedicato, M.T.; Zupa, W.; Vitale, F.; Follesa, M.C. The spawning strategy of European hake (Merluccius merluccius, L. 1758) across the Western and Central Mediterranean Sea. Fish. Res. 2019, 219, 105333. [Google Scholar] [CrossRef]
- Candelma, M.; Marisaldi, L.; Bertotto, D.; Radaelli, G.; Gioacchini, G.; Santojanni, A.; Colella, S.; Carnevali, O. Aspects of Reproductive Biology of the European Hake (Merluccius merluccius) in the Northern and Central Adriatic Sea (GSA 17-Central Mediterranean Sea). J. Mar. Sci. Eng. 2021, 9, 389. [Google Scholar] [CrossRef]
- Olaso, I.; Sànchez, F.; Rodrìguez-Cabello, C.; Velasco, F. The feeding behaviour of some demersal fish species in response to artificial discarding. Sci. Mar. 2002, 66, 301–311. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecological Methodology; Harper & Row Publisher: New York, NY, USA, 1989. [Google Scholar]
- Fanelli, E.; Papiol, V.; Cartes, J.E.; Rumolo, P.; Brunet, C.; Sprovieri, M. Food web structure of the epibenthic and infaunal invertebrates on the Catalan slope (NW Mediterranean): Evidence from δ13C and δ15N analysis. Deep Sea Res. Part I Oceanogr. Res. Pap. 2011, 58, 98–109. [Google Scholar] [CrossRef]
- Logan, M.J.; Jardine, T.D.; Miller, T.J.; Bunn, S.E.; Cunjak, R.A.; Lutcavage, M.E. Lipid Corrections in Carbon and Nitrogen Stable Isotope Analyses: Comparison of Chemical Extraction and Modelling Methods. J. Anim. Ecol. 2008, 77, 838–846. [Google Scholar] [CrossRef] [PubMed]
- Post, D.M.; Layman, C.A.; Arrington, D.A.; Takimoto, G.; Quattrochi, J.; Montana, C.G. Getting to the fat of the matter: Models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia 2007, 152, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.; Gorley, R.N.; Clarke, K. PERMANOVA+ for Primer: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER V6: User Manual/Tutorial; Plymouth Marine Laboratory: Plymouth, UK, 2006. [Google Scholar]
- Clarke, K.; Warwick, R. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; Primer-E Ltd: Plymouth, UK, 2001. [Google Scholar]
- Gower, J.C. Some Distance Properties of Latent Root and Vector Methods Used in Multivariate Analysis. Biometrika 1966, 53, 325–338. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: http://www.R-project.org (accessed on 2 October 2022).
- Parnell, A.C. simmr: A Stable Isotope Mixing Model. R Package Version 0.4.5. Available online: https://CRAN.R-project.org/package=simmr (accessed on 2 October 2022).
- Vanderklift, M.A.; Ponsard, S. Sources of variation in consumer-diet delta 15N enrichment: A meta-analysis. Oecologia 2003, 136, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Caut, S.; Angulo, E.; Courchamp, F. Variation in discrimination factors (Δ15N and Δ13C): The effect of diet isotopic values and applications for diet reconstruction. J. Appl. Ecol. 2009, 46, 443–453. [Google Scholar] [CrossRef]
- Smith, J.A.; Mazumder, D.; Suthers, I.M.; Taylor, M.D.; Bowen, G. To fit or not to fit: Evaluating stable isotope mixing models using simulated mixing polygons. Methods Ecol. Evol. 2013, 4, 612–618. [Google Scholar] [CrossRef]
- Phillips, D.L.; Inger, R.; Bearhop, S.; Jackson, A.L.; Moore, J.W.; Parnell, A.C.; Semmens, B.X.; Ward, E.J. Best practices for use of stable isotope mixing models in food-web studies. Can. J. Zool. 2014, 92, 823–835. [Google Scholar] [CrossRef]
- Fanelli, E.; Menicucci, S.; Malavolti, S.; Da Ros, Z.; De Felice, A.; Leonori, I. Ontogenetic and spatial changes in the feeding habits and trophic levels of small pelagic fishes in the Mediterranean sea. Front. Mar. Sci. 2022; submitted. [Google Scholar]
- Fanelli, E.; Menicucci, S.; Malavolti, S.; De Felice, A.; Leonori, I. Mesoscale variations in the assemblage structure and trophodynamics of mesozooplankton communities of the Adriatic basin (Mediterranean Sea). Biogeosciences 2021, 2012, 1–33. [Google Scholar] [CrossRef]
- Bongiorni, L.; Nasi, F.; Fiorentino, F.; Auriemma, R.; Rampazzo, F.; Nordstrom, M.C.; Berto, D. Contribution of deltaic wetland food sources to coastal macrobenthic consumers (Po River Delta, north Adriatic Sea). Sci. Total Environ. 2018, 643, 1373–1386. [Google Scholar] [CrossRef]
- Fanelli, E.; Dell’Anno, A.; Nepote, E.; Lo Martire, M.; Musco, L.; Bianchelli, S.; Gambi, C.; Falco, F.; Memmola, F.; Coluccelli, A.; et al. Positive effects of two decades of passive restoration in a highly polluted marine site. Water Res. 2022; submitted. [Google Scholar]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER—Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef]
- Layman, C.A.; Quattrochi, J.P.; Peyer, C.M.; Allgeier, J.E. Niche width collapse in a resilient top predator following ecosystem fragmentation. Ecol. Lett. 2007, 10, 937–944. [Google Scholar] [CrossRef]
- Aarnio, K.; Bonsdorff, E.; Rosenback, N. Food and feeding habits of juvenile flounder Platichthys flesus (L.), abd turbot Scophthalmus maximus L. in the Aland archipelago, northern Baltic Sea. J. Sea Res. 1996, 36, 311–320. [Google Scholar] [CrossRef]
- Methven, D. Annotated Bibliography of Demersal Fish Feeding with Emphasis on Selected Studies from the Scotian Shelf and Grand Banks of the Northwestern Atlantic; Department of Fisheries and Oceans.: St. John’s, NL, Canada, 1999. [Google Scholar]
- El-mor, M.; Ashraf, I.A. Feeding habits of the common sole, Solea vulgaris (Quensel, 1806), from Mediterranean Sea, Port Said, Egypt. Egypt. J. Aquat. Biol. Fish. 2008, 12, 51–61. [Google Scholar] [CrossRef][Green Version]
- Rajaguru, A. Biology of two co-occurring tonguefishes, Cynoglossus arel and C. lida (Pleuronectiformes: Cynoglossidae), from Indian waters. Fish. Bull. 1992, 90, 328–367. [Google Scholar]
- Quiniou, L. Les Poissons Demersaux de la baie de Douarnenez; Alimentation et ecologie: Brest, France, 1978. [Google Scholar]
- Henderson, P.A.; James, D.; Holmes, R.H.A. Trophic structure within the Bristol Channel: Seasonality and stability in Bridgwater Bay. J. Mar. Biol. Assoc. UK 1992, 72, 675–690. [Google Scholar] [CrossRef]
- Collie, J.S. Food consumption by yellowtail flounder in relation to production of its benthic prey. Mar. Ecol. Prog. Ser. 1987, 36, 205–213. [Google Scholar] [CrossRef]
- Stickney, R.R. Food Habits of Georgia Estuarine Fishes II. Symphurus plagiusa (Pleuronectiformes: Cynoglossidae). Trans. Am. Fish. Soc. 1976, 105, 202–207. [Google Scholar] [CrossRef]
- de Morais, T.L.; Bodiou, J.Y. Predation on meiofauna by juvenile fish in a Western Mediterranean flatfish nursery ground. Mar. Biol. 1984, 82, 209–215. [Google Scholar] [CrossRef]
- Cabral, H. Comparative feeding ecology of sympatric Solea solea and S. senegalensis, within the nursery areas of the Tagus estuary, Portugal. J. Fish Biol. 2000, 57, 1550–1562. [Google Scholar] [CrossRef]
- Amara, R.; Mahé, K.; LePape, O.; Desroy, N. Growth, feeding and distribution of the solenette Buglossidium luteum with particular reference to its habitat preference. J. Sea Res. 2004, 51, 211–217. [Google Scholar] [CrossRef][Green Version]
- Ramos, J. Regimeny comportamiento alimentario del lenguado (Solea solea L.) (Pisces, Soleidae). Lnformes Tec. Del Inst. De Investig. Pesq. 1981, 83, 3–15. [Google Scholar]
- Seshappa, G.; Bhimacar, B.S. Studies on the fishery and biology of malabar sole, Cynoglossus semifasciatus day. Indian J. Fish. 1955, 2, 180–230. [Google Scholar]
- Zacchetti, L.; Martinelli, M.; Colella, S.; Santojanni, A.; Fanelli, E. Seasonal variations in the feeding ecology of Nephrops norvegicus in the Adriatic Sea: Insights from stomach contents and stable isotope analyses. Mar. Ecol. Prog. Ser. 2022, 695, 109–123. [Google Scholar] [CrossRef]
- STEFC. 23rd Report of the Scientific, Technical and Economic Committee for Fisheries (Second Plenary Meeting); Publications Office of the European Union: Barza d’Ispra, Italy, 2006. [Google Scholar]
- Eigaard, O.R.; Bastardie, F.; Hintzen, N.T.; Buhl-Mortensen, L.; Buhl-Mortensen, P.; Catarino, R.; Dinesen, G.E.; Egekvist, J.; Fock, H.O.; Geitner, K.; et al. The footprint of bottom trawling in European waters: Distribution, intensity, and seabed integrity. ICES J. Mar. Sci. 2017, 74, 847–865. [Google Scholar] [CrossRef]
- Milisenda, G.; Garofalo, G.; Fiorentino, F.; Colloca, F.; Maynou, F.; Ligas, A.; Musumeci, C.; Bentes, L.; Gonçalves, J.M.S.; Erzini, K.; et al. Identifying Persistent Hot Spot Areas of Undersized Fish and Crustaceans in Southern European Waters: Implication for Fishery Management Under the Discard Ban Regulation. Front. Mar. Sci. 2021, 8, 610241. [Google Scholar] [CrossRef]
- Amundsen, P.A.; Sanchez-Hernandez, J. Feeding studies take guts—critical review and recommendations of methods for stomach contents analysis in fish. J. Fish Biol. 2019, 95, 1364–1373. [Google Scholar] [CrossRef]
- Nielsen, J.M.; Clare, E.L.; Hayden, B.; Brett, M.T.; Kratina, P.; Gilbert, M.T.P. Diet tracing in ecology: Method comparison and selection. Methods Ecol. Evol. 2017, 9, 278–291. [Google Scholar] [CrossRef]
- Maiorano, P.; Sabatella, R.F.; Marzocchi, B.M. Annuario Sullo Stato Delle Risorse e Sulle Strutture Produttive dei mari Italiani. 2019. 432p. Available online: http://www.nisea.eu/dir/wp-content/uploads/2019/09/Annuario-20142016_2019_08_05.pdf (accessed on 2 October 2022).
- Ferrer-Maza, D.; Muñoz, M.; Lloret, J.; Faliex, E.; Vila, S.; Sasal, P. Health and reproduction of red mullet, Mullus barbatus, in the western Mediterranean Sea. Hydrobiologia 2015, 753, 189–204. [Google Scholar] [CrossRef]
- Pacetti, D.; Balzano, M.; Colella, S.; Santojanni, A.; Frega, N.G. Effect of spawning on furan fatty acid profile of edible muscle and organ tissues from sardine (Sardina pilchardus) and anchovy (Engraulis encrasicolus). J. Agric. Food Chem. 2013, 61, 3969–3977. [Google Scholar] [CrossRef] [PubMed]
- Dall, W. Lipid absorption and utilization in the Norwegian lobster, Nephrops norvegicus (L.). J. Exp. Mar. Biol. Ecol. 1981, 50, 33–45. [Google Scholar] [CrossRef]
- Ahmed, A.; Sharaf, M.; Laban, H. Reproduction of the Egyptian sole, Solea aegyptiaca (Actinopterygii: Pleuronectiformes: Soleidae), from Port Said, Egypt, Mediterranean Sea. Acta Ichthyol. Et Piscat. 2010, 40, 161–166. [Google Scholar] [CrossRef]
- Relini, G.; Tixi, F.; Relini, M.; Torchia, G. The macrofouling on offshore platforms at Ravenna. Int. Biodeterior. Biodegrad. 1998, 41, 41–55. [Google Scholar] [CrossRef]
- Fanelli, E.; Da Ros, Z.; Martino, I.; Azzurro, E.; Bargione, G.; Donato, F.; Lucchetti, A. Crowding in the middle of marine food webs: A focus on Raja asterias and other mediterranean batoids. Mar. Environ. Res. 2022, 183, 105830. [Google Scholar] [CrossRef]
- Cortés, E. Standardized diet compositions and trophic levels of sharks. ICES J. Mar. Sci. 1999, 56, 707–717. [Google Scholar] [CrossRef]
- Keser, R.; Bray, R.; Oguz, M.; Çelen, S.; Erdoğan, S.; Doğuturk, S.; Aklanoğlu, G.; Marti, B. Helminth parasites of digestive tract of some teleost fish caught in the Dardanelles at Çanakkale, Turkey. Helminthologia 2007, 44, 217–221. [Google Scholar] [CrossRef]
- Abdel-Ghaffar, F.; Abdel-Gaber, R.; Bashtar, A.R.; Morsy, K.; Mehlhorn, H.; Al Quraishy, S.; Saleh, R. Hysterothylacium aduncum (Nematoda, Anisakidae) with a new host record from the common sole Solea solea (Soleidae) and its role as a biological indicator of pollution. Parasitol. Res. 2015, 114, 513–522. [Google Scholar] [CrossRef]
- Tsagarakis, K.; Palialexis, A.; Vassilopoulou, V. Mediterranean fishery discards: Review of the existing knowledge. ICES J. Mar. Sci. 2014, 71, 1219–1234. [Google Scholar] [CrossRef]
- Santojanni, A.; Cingolani, N.; Arneri, E.; Kirkwood, G.; Belardinelli, A.; Giannetti, G.; Colella, S.; Donato, F.; Barry, C. Stock assessment of sardine (Sardina pilchardus, Walb.) in the Adriatic Sea with an estimate of discards. Sci. Mar. 2005, 69, 603–617. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Df | Deviance | Resid. Df | Resid. Dev | F | Pr(>F) | ||
|---|---|---|---|---|---|---|---|
| NULL | 308 | 10.03 | |||||
| HSI | 1 | 0.55 | 307 | 9.48 | 17.92 | 0.00003 | + |
| GSI | 1 | 0.15 | 306 | 9.33 | 5.07 | 0.03 | - |
| Season | TA | SEAC | CD |
|---|---|---|---|
| Spring | 2.1 | 1.5 | 0.83 |
| Summer | 2.8 | 0.9 | 0.66 |
| Autumn | 1.6 | 0.6 | 0.56 |
| Winter | 1.8 | 0.8 | 0.58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fanelli, E.; Principato, E.; Monfardini, E.; Da Ros, Z.; Scarcella, G.; Santojanni, A.; Colella, S. Seasonal Trophic Ecology and Diet Shift in the Common Sole Solea solea in the Central Adriatic Sea. Animals 2022, 12, 3369. https://doi.org/10.3390/ani12233369
Fanelli E, Principato E, Monfardini E, Da Ros Z, Scarcella G, Santojanni A, Colella S. Seasonal Trophic Ecology and Diet Shift in the Common Sole Solea solea in the Central Adriatic Sea. Animals. 2022; 12(23):3369. https://doi.org/10.3390/ani12233369
Chicago/Turabian StyleFanelli, Emanuela, Elena Principato, Eleonora Monfardini, Zaira Da Ros, Giuseppe Scarcella, Alberto Santojanni, and Sabrina Colella. 2022. "Seasonal Trophic Ecology and Diet Shift in the Common Sole Solea solea in the Central Adriatic Sea" Animals 12, no. 23: 3369. https://doi.org/10.3390/ani12233369
APA StyleFanelli, E., Principato, E., Monfardini, E., Da Ros, Z., Scarcella, G., Santojanni, A., & Colella, S. (2022). Seasonal Trophic Ecology and Diet Shift in the Common Sole Solea solea in the Central Adriatic Sea. Animals, 12(23), 3369. https://doi.org/10.3390/ani12233369