
Progesterone Induces Apoptosis and Steroidogenesis in Porcine Placental Trophoblasts


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Reagents
2.3. Cell Culture
2.4. Enzyme-Linked Immunosorbent Assay (ELISA) and Cell Counting
2.5. RNA Extraction and Quantitative Reverse-Transcriptase PCR (RT-qPCR)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence of Primers (5′ to 3′) | Fragment (bp) | GenBank ID | Reference |
|---|---|---|---|---|
| CCND1 | F: GACCGCTTCCTGTCCCTGR: GTGGCACAGAGGGCGACGA | 317 | XM_021082686 | [8] |
| CCND2 | F: CGTCCAAGCTCAAAGAGACCR: CGAAGAATGTGCTCGATGAA | 169 | NM_214088 | [36] |
| CDK4 | F: GCATCCCAATGTTGTCCGR: GGGGTGCCTTGTCCAGATA | 125 | NM_001123097 | [8] |
| CASP8 | F: TCCTGAGCCTGGACTACATR: CTCCTCCTCATTGGTTTCC | 185 | NM_001031779.2 | [8] |
| CASP3 | F: GCCATGGTGAAGAAGGAAAAR: GTCCGTCTCAATCCCACAGT | 167 | NM_214131 | [8] |
| StAR | F: GGAAAAGACACAGTCATCACCCATR: CAGCCAGCACACACACGGAAC | 121 | NM_213755.2 | [8] |
| HSD17B4 | F: TGCCATGAGAGTTGTGAGGAAAR: CCTCAGGAGTCATTGGCTGATT | 127 | XM_021081514.1 | [8] |
| HSD3B | F: TCCACACCAGCAGCATAGAGR: ATACATGGGCCTCAGAGCAC | 206 | NM_001004049.2 | [8] |
| CYP19A1 | F: GTATATCGCCATGGTCATGR: AGCAGGCCGCTGGTCTCAT | 144 | NM_214429.1 | [8] |
| CYP11A1 | F: GCCGCATGGGACACTATTTTR: ATTTCCCAGGAGGCGGTAGA | 120 | NM_214427.1 | [8] |
| Bax | F: AAGCGCATTGGAGATGAACTR: CGATCTCGAAGGAAGTCCAG | 251 | XM_003127290.5 | [37] |
| Bcl–2 | F: TGTGTGGAGAGCGTCAACCGR: CCCATACAGCTCCACAAAGGCAT | 138 | XM_021099593.1 | [38] |
| PGR | F: GATTCAGAAGCCAGCCAGAGR: GATGCTTCATCCCCACAGAT | 83 | GQ903679 | [39] |
| GAPDH | F: AAGGAGTAAGAGCCCCTGGAR: TCTGGGATGGAAACTGGAA | 140 | NM_001206359.1 | [8] |
2.6. Western Blot Analysis
2.7. Experimental Design
2.8. Statistical Analysis
3. Results
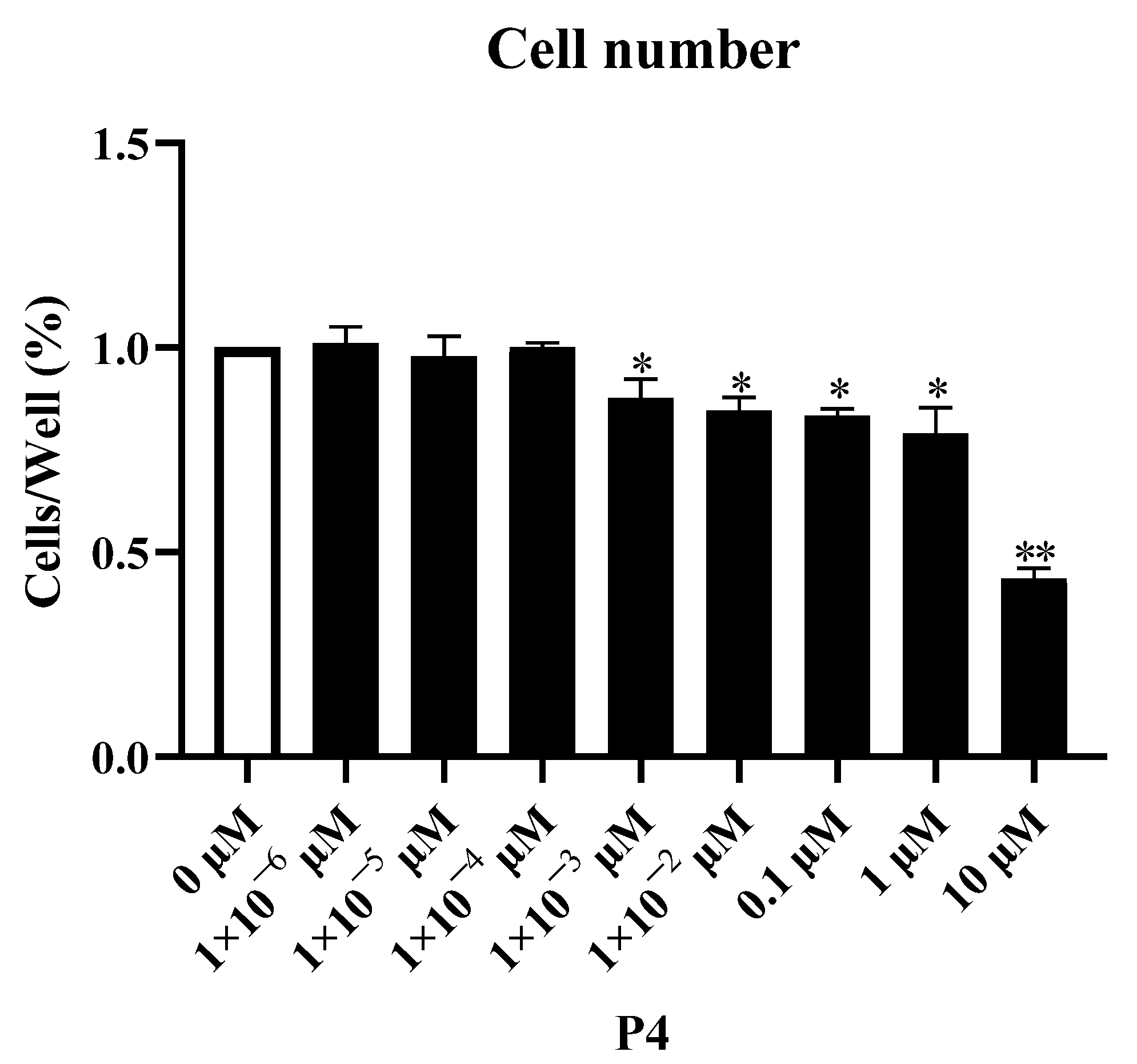
3.1. Effect of P4 on Proliferation of pTr Cells
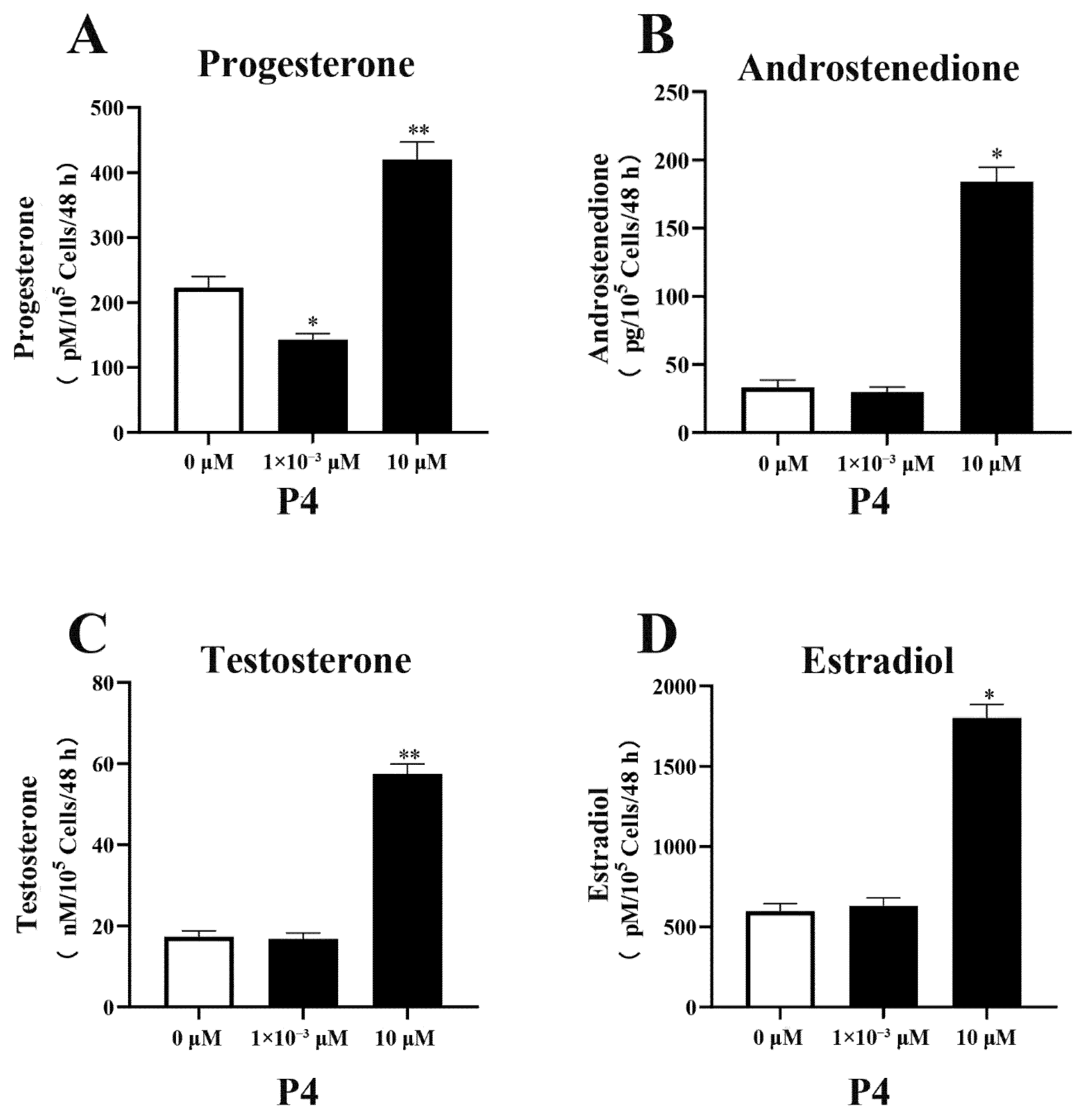
3.2. Effect of P4 on Steroid Hormone Synthesis in pTr Cells
3.3. Effects of P4 on Gene Expression in pTr Cells
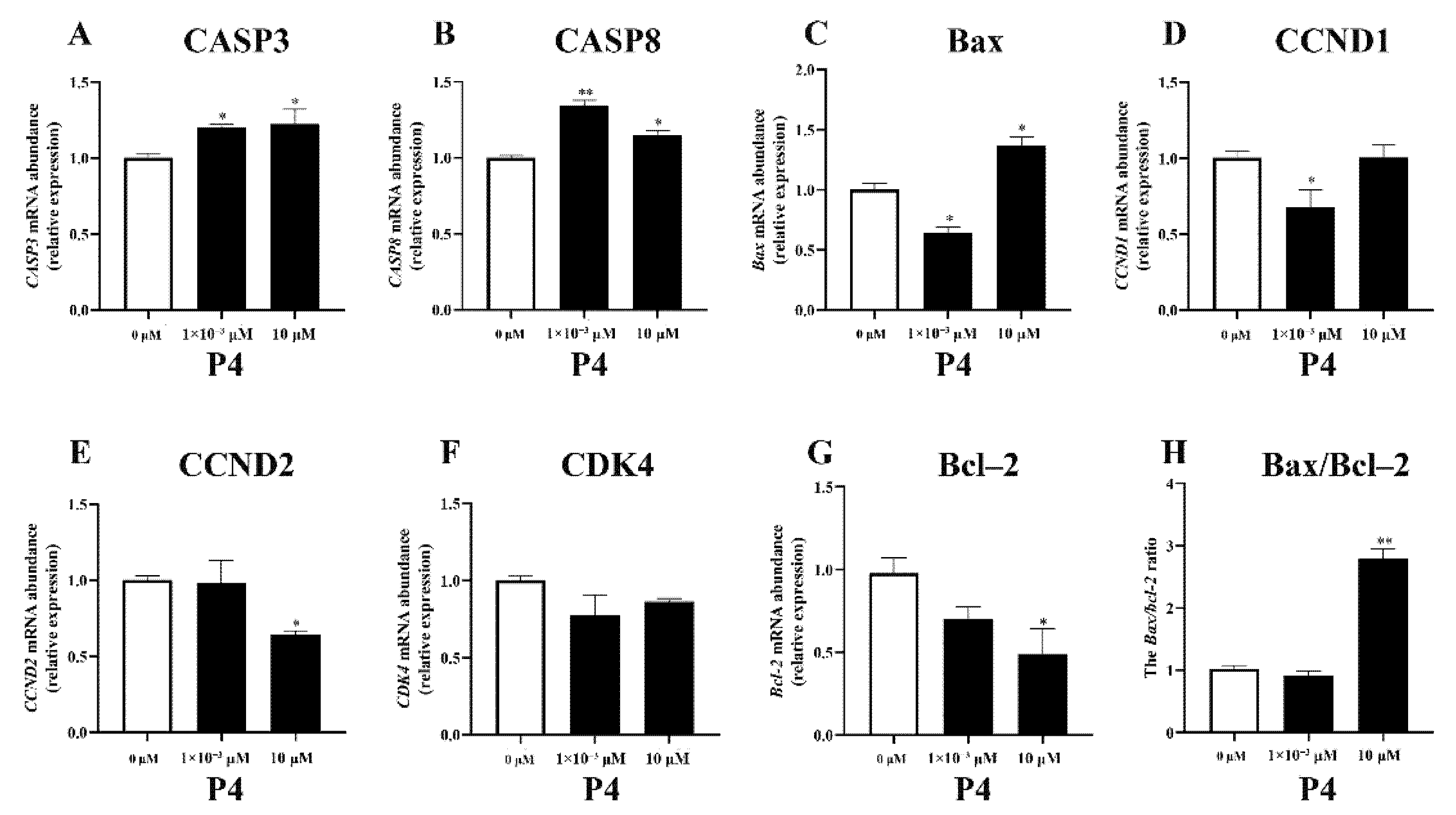
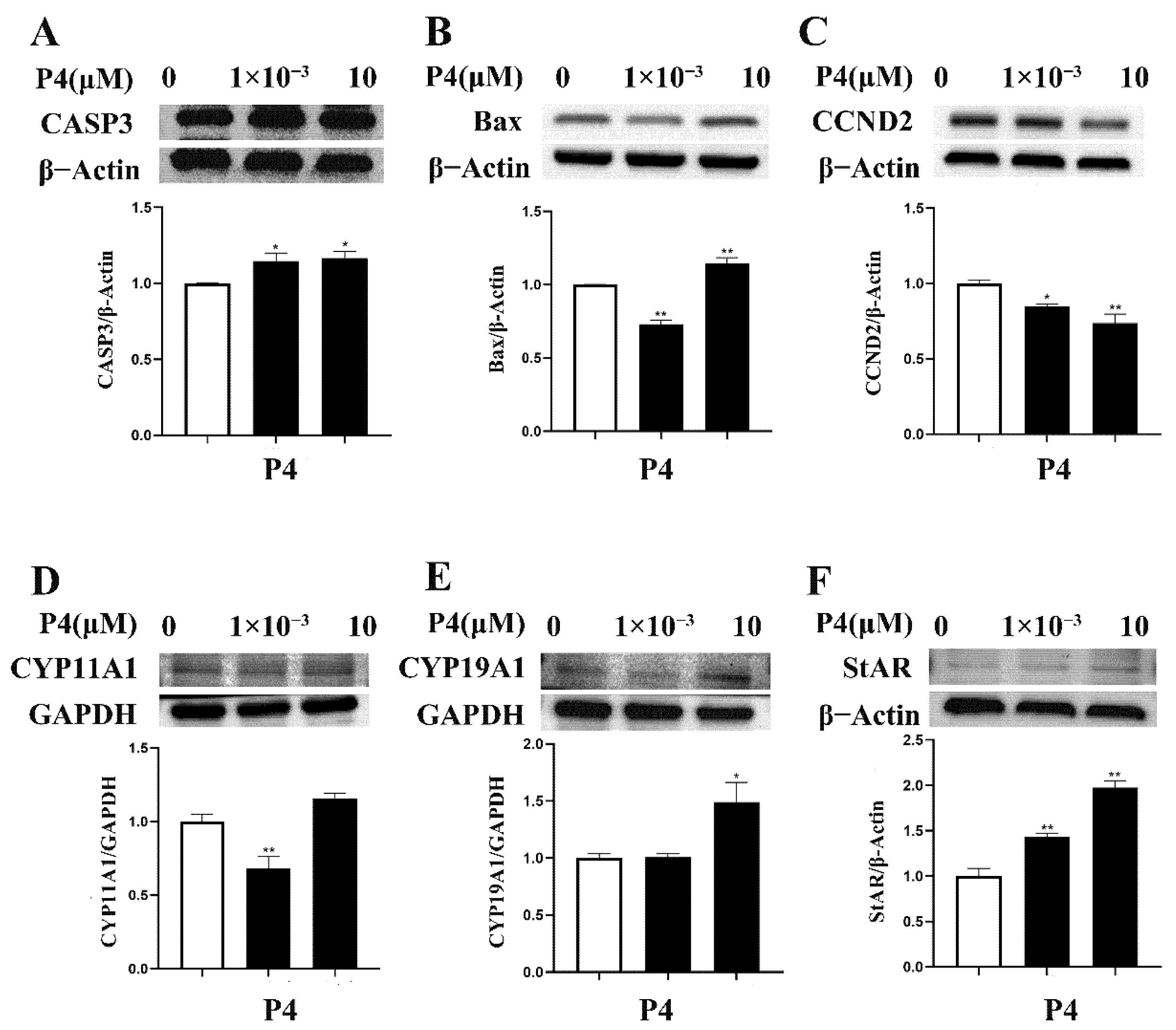
3.3.1. Cell-Proliferation-/Apoptosis-Related Gene Expression
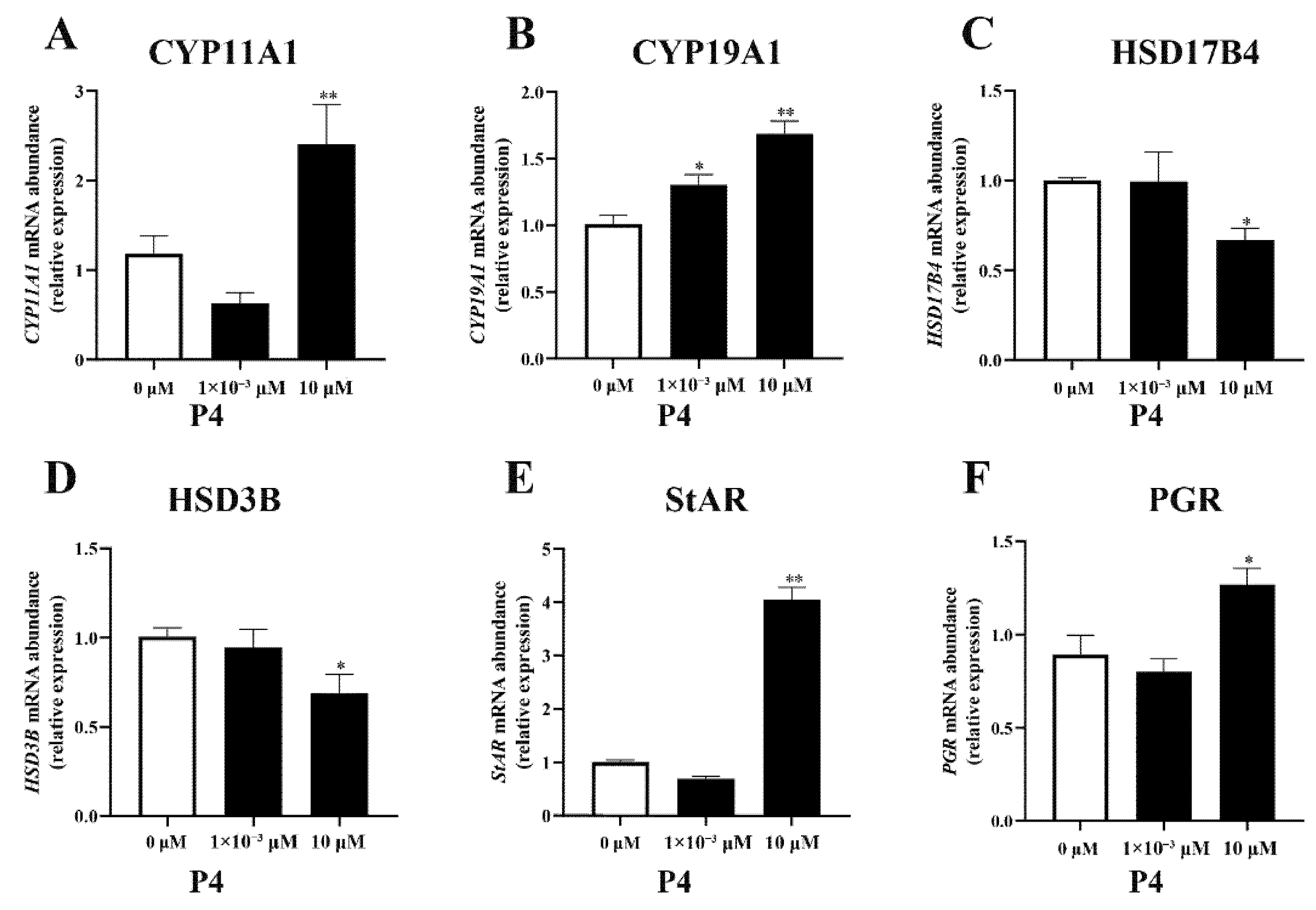
3.3.2. Expression of Steroidogenesis-Related Genes
3.4. Effects of P4 on Protein Expression in pTr Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cameo, P.; Bischof, P.; Calvo, J.C. Effect of leptin on progesterone, human chorionic gonadotropin, and interleukin-6 secretion by human term trophoblast cells in culture. Biol. Reprod. 2003, 68, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Chavatte-Palmer, P.; Tarrade, A. Placentation in different mammalian species. Ann. Endocrinol. 2016, 77, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Rielland, M.; Hue, I.; Renard, J.P.; Alice, J. Trophoblast stem cell derivation, cross-species comparison and use of nuclear transfer: New tools to study trophoblast growth and differentiation. Dev. Biol. 2008, 322, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Wang, X.; Yin, Y.; Li, X.; Gao, H.; Bazer, F.W.; Wu, G. Putrescine stimulates the mTOR signaling pathway and protein synthesis in porcine trophectoderm cells. Biol. Reprod. 2014, 91, 106. [Google Scholar] [CrossRef]
- Kim, J.; Song, G.; Wu, G.; Bazer, F.W. Functional roles of fructose. Proc. Natl. Acad. Sci. USA 2012, 109, E1619–E1628. [Google Scholar] [CrossRef]
- Guo, P. Effects of Arginine on Reproductive Performance in Gilts and the Proliferation of Porcine Placenta Trophectoderm Cells; Northeast Agricultural University: Harbin, China, 2017. (In Chinese) [Google Scholar]
- Suleman, M.; Malgarin, C.M.; Detmer, S.E.; Harding, J.; MacPhee, D.J. The porcine trophoblast cell line PTr2 is susceptible to porcine reproductive and respiratory syndrome virus-2 infection. Placenta 2019, 88, 44–51. [Google Scholar] [CrossRef]
- Wei, S.L.; Yang, Y.Z.; Xiao, Y.X.; Liu, Y.; Tian, J.H.; Spicer, L.J.; Feng, T. Effects of N-carbamylglutamate on steroidogenesis and relative abundances of mRNA transcripts in pig placental trophoblasts. Anim. Reprod. Sci. 2020, 221, 106569. [Google Scholar] [CrossRef]
- Wei, S.L.; Zheng, C.; Liu, Y.; Feng, T. Progress in research on the porcine placental barrier. Animal Husbandry & Veterinary Medicine. 2021, 53, 148–154. (In Chinese) [Google Scholar]
- Dimasuay, K.G.; Boeuf, P.; Powell, T.L.; Jansson, T. Placental responses to changes in the maternal environment determine fetal growth. Front Physiol. 2016, 7, 12. [Google Scholar] [CrossRef]
- Strauss, J.F.; Martinez, F.; Kiriakidou, M. Placental steroid hormone synthesis: Unique features and unanswered questions. Biol. Reprod. 1996, 54, 303–311. [Google Scholar] [CrossRef]
- Cui, X.; Sun, J.; Liang, C.; Zheng, Q.; Yang, X.; Liu, S.; Yan, Q. Progesterone promotes embryo adhesion by upregulating c-Fos/c-Jun transcription factor-mediated poFUT1 expression. Biol. Reprod. 2019, 101, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Almeida, F.; Dias, A. Pregnancy in pigs: The journey of an early life. Domest. Anim. Endocrinol. 2022, 78, 106656. [Google Scholar] [CrossRef] [PubMed]
- Song, H.B. Prospective Study of Dynamic Monitoring of Serum β-hCG and Progesterone to Predict Early Pregnancy Outcome. Master’s Thesis, Qingdao University, Qingdao, China, January 2020. [Google Scholar]
- Solano, M.E.; Arck, P.C. Steroids, Pregnancy and Fetal Development. Front. Immunol. 2019, 10, 3017. [Google Scholar] [CrossRef]
- Johansson, E.D.; Jonasson, L.E. Progesterone levels in amniotic fluid and plasma from women. I. Levels during normal pregnancy. Acta. Obstet. Gynecol. Scand. 1971, 50, 339–343. [Google Scholar] [CrossRef]
- Virgo, B.B.; Bellward, G.D. Serum progesterone levels in the pregnant and postpartum laboratory mouse. Endocrinology 1974, 95, 1486–1490. [Google Scholar] [CrossRef]
- Nilsson, E.E.; Skinner, M.K. Progesterone regulation of primordial follicle assembly in bovine fetal ovaries. Mol. Cell. Endocrinol. 2009, 313, 9–16. [Google Scholar] [CrossRef]
- Liu, X.J.; Zhang, D.W.; Mai, H.M.; Luo, J.Y.; Chen, H.M. Serum progesterone and insulin-like growth factor in predicting adverse outcomes in early pregnancy. Med. Innov. China 2019, 16, 30–33. (In Chinese) [Google Scholar]
- Breier, G.; Albrecht, U.; Sterrer, S.; Risau, W. Expression of vascular endothelial growth factor during embryonic angiogenesis and endothelial cell differentiation. Development 1992, 114, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Rätsep, M.T.; Felker, A.M.; Kay, V.R.; Tolusso, L.; Hofmann, A.P.; Croy, B.A. Uterine natural killer cells: Supervisors of vasculature construction in early decidua basalis. Reproduction 2015, 149, R91–R102. [Google Scholar] [CrossRef]
- Jewson, M.; Purohit, P.; Lumsden, M.A. Progesterone and abnormal uterine bleeding/menstrual disorders. Best Pract. Res. Clin. Obst. Gynaecol. 2020, 69, 62–73. [Google Scholar] [CrossRef]
- Su, Y.; Li, Q.; Zhang, Q.; Li, Z.; Yao, X.; Guo, Y.; Xiao, L.; Wang, X.; Ni, H. Exosomes derived from placental trophoblast cells regulate endometrial epithelial receptivity in dairy cows during pregnancy. J. Reprod. Dev. 2022, 68, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Obr, A.E.; Grimm, S.L.; Bishop, K.A.; Pike, J.W.; Lydon, J.P.; Edwards, D.P. Progesterone receptor and Stat5 signaling cross talk through RANKL in mammary epithelial cells. Mol. Endocrinol. 2013, 27, 1808–1824. [Google Scholar] [CrossRef]
- Bazer, F.W.; Burghardt, R.C.; Johnson, G.A.; Spencer, T.E.; Wu, G. Mechanisms for the establishment and maintenance of pregnancy: Synergies from scientific collaborations. Biol. Reprod. 2018, 99, 225–241. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Xie, L.; Xu, Z.; Hao, M.; Yang, B.; Shan, W.; Wang, Y.; Lv, Q.; Chen, X. NrCAM secreted by endometrial stromal cells enhances the progestin sensitivity of endometrial cancer cells through epigenetic modulation of PRB. Cancer Gene Ther. 2022. [Google Scholar] [CrossRef] [PubMed]
- Leonhardt, S.A.; Edwards, D.P. Mechanism of action of progesterone antagonists. Exp. Biol. Med. 2002, 227, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Feinshtein, V.; Ben-Zvi, Z.; Sheiner, E.; Amash, A.; Sheizaf, B.; Holcberg, G. Progesterone levels in cesarean and normal delivered term placentas. Arch. Gynecol. Obstet. 2010, 281, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Lissauer, D.; Eldershaw, S.A.; Inman, C.F.; Coomarasamy, A.; Moss, P.A.; Kilby, M.D. Progesterone promotes maternal-fetal tolerance by reducing human maternal T-cell polyfunctionality and inducing a specific cytokine profile. Eur. J. Immunol. 2015, 45, 2858–2872. [Google Scholar] [CrossRef]
- Lissauer, D.M.; Piper, K.P.; Moss, P.A.; Kilby, M.D. Fetal microchimerism: The cellular and immunological legacy of pregnancy. Expert Rev. Mol. Med. 2009, 11, e33. [Google Scholar] [CrossRef]
- Cai, S.; Zhu, J.; Zeng, X.; Ye, Q.; Ye, C.; Mao, X.; Zhang, S.; Qiao, S.; Zeng, X. Maternal N-Carbamylglutamate Supply during Early Pregnancy Enhanced Pregnancy Outcomes in Sows through Modulations of Targeted Genes and Metabolism Pathways. J. Agric. Food Chem. 2018, 66, 5845–5852. [Google Scholar] [CrossRef]
- Ding, H.; Yang, Y.; Wei, S.; Spicer, L.J.; Kenéz, Á.; Xu, W.; Liu, Y.; Feng, T. Influence of N-acetylcysteine on steroidogenesis and gene expression in porcine placental trophoblast cells. Theriogenology 2021, 161, 49–56. [Google Scholar] [CrossRef]
- Liu, Y.; Bai, J.H.; Xu, X.L.; Chen, Z.L.; Spicer, L.J.; Feng, T. Effects of N-carbamylglutamate and L-arginine on gonadotrophin-releasing hormone (GnRH) gene expression and secretion in GT1-7 cells. Reprod. Fertil. Dev. 2018, 30, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Schütz, L.F.; Morrell, B.C.; Perego, M.C.; Spicer, L.J. Effects of N-carbamylglutamate and L-arginine on steroidogenesis and gene expression in bovine granulosa cells. Anim. Reprod. Sci. 2018, 188, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, W.; Wang, S.; Liu, H.; Zhang, D.; Wang, Y.; Ji, H. Swine-derived probiotic lactobacillus plantarum modulates porcine intestinal endogenous host defense peptide synthesis through TLR2/MAPK/AP-1 signaling pathway. Front. Immunol. 2019, 10, 2691. [Google Scholar] [CrossRef] [PubMed]
- Chermuła, B.; Jeseta, M.; Sujka-Kordowska, P.; Konwerska, A.; Jankowski, M.; Kranc, W.; Kocherova, I.; Celichowski, P.; Antosik, P.; Bukowska, D.; et al. Genes regulating hormone stimulus and response to protein signaling revealed differential expression pattern during porcine oocyte in vitro maturation, confirmed by lipid concentration. Histochem. Cell Biol. 2020, 154, 77–95. [Google Scholar] [CrossRef]
- Kang, H.G.; Lee, S.; Jeong, P.S.; Kim, M.J.; Park, S.H.; Joo, Y.E.; Park, S.H.; Song, B.S.; Kim, S.U.; Kim, M.K.; et al. Lycopene improves in vitro development of porcine embryos by reducing oxidative stress and apoptosis. Antioxidants 2021, 10, 230. [Google Scholar] [CrossRef]
- Zhu, Z.; Pan, Q.; Zhao, W.; Wu, X.; Yu, S.; Shen, Q.; Zhang, J.; Yue, W.; Peng, S.; Li, N.; et al. BCL2 enhances survival of porcine pluripotent stem cells through promoting FGFR2. Cell Prolif. 2021, 54, e12932. [Google Scholar] [CrossRef]
- Mathew, D.J.; Sellner, E.M.; Green, J.C.; Okamura, C.S.; Anderson, L.L.; Lucy, M.C.; Geisert, R.D. Uterine progesterone receptor expression, conceptus development, and ovarian function in pigs treated with RU 486 during early pregnancy. Biol. Reprod. 2011, 84, 130–139. [Google Scholar] [CrossRef]
- Formby, B.; Wiley, T.S. Bcl-2, survivin and variant CD44 v7-v10 are downregulated and p53 is upregulated in breast cancer cells by progesterone: Inhibition of cell growth and induction of apoptosis. Mol. Cell Biochem. 1999, 202, 53–61. [Google Scholar] [CrossRef]
- Li, M.Q.; Xie, F.; Shi, J.W.; Yang, H.L.; Lai, Z.Z.; Shen, H.H.; Ruan, L.Y.; Wang, Y.; Qiu, X.M. Aspirin enhances the protective effect of progesterone on trophoblast cell from oxidative stress and apoptosis. Reprod. Dev. Med. 2021, 5, 1–8. [Google Scholar]
- Pei, J.; Liu, Z.; Wang, C.; Chu, N.; Liu, L.; Tang, Y.; Liu, H.; Xiang, Q.; Cheng, H.; Li, M.; et al. Progesterone attenuates sirt1-deficiency-mediated pre-eclampsia. Biomolecules 2022, 12, 422. [Google Scholar] [CrossRef]
- Formby, B.; Wiley, T.S. Progesterone inhibits growth and induces apoptosis in breast cancer cells: Inverse effects on Bcl-2 and p53. Ann. Clin. Lab Sci. 1998, 28, 360–369. [Google Scholar] [PubMed]
- Sasaki, G.; Zubair, M.; Ishii, T.; Mitsui, T.; Hasegawa, T.; Auchus, R.J. The contribution of serine 194 phosphorylation to steroidogenic acute regulatory protein function. Mol. Endocrinol. 2014, 28, 1088–1096. [Google Scholar] [CrossRef] [PubMed]
- Payne, A.H.; Hales, D.B. Overview of steroidogenic enzymes in the pathway from cholesterol to active steroid hormones. Endocr. Rev. 2004, 25, 947–970. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Wan, W.; Zhu, K.; Pan, M.; Zhao, X.; Ma, B.; Wei, Q. Effects of 4-vinylcyclohexene diepoxide on the cell cycle, apoptosis, and steroid hormone secretion of goat ovarian granulosa cells. Vitr. Cell Dev. Biol. Anim. 2022, 58, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez Castaño, P.; Parween, S.; Pandey, A.V. Bioactivity of curcumin on the cytochrome p450 enzymes of the steroidogenic pathway. Int. J. Mol. Sci. 2019, 20, 4606. [Google Scholar] [CrossRef]
- Salem, H.F. Sustained-release progesterone nanosuspension following intramuscular injection in ovariectomized rats. Int. J. Nanomed. 2010, 5, 943–954. [Google Scholar] [CrossRef]
- Sun, D.; Cui, Y.; Jin, B.; Zhang, X.; Yang, X.; Gao, C. Effects of the yangjing capsule extract on steroidogenesis and apoptosis in mouse leydig cells. Evid Based Complement Alternat. Med. 2012, 2012, 985457. [Google Scholar] [CrossRef]
- Fox, C.W.; Zhang, L.; Sohni, A.; Doblado, M.; Wilkinson, M.F.; Chang, R.J.; Duleba, A.J. Inflammatory Stimuli Trigger Increased Androgen Production and Shifts in Gene Expression in Theca-Interstitial Cells. Endocrinology 2019, 160, 2946–2958. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Ji, M.; Wen, X.; Chen, D.; Huang, F.; Guan, X.; Tian, J.; Xie, J.; Shao, J.; Wang, J.; et al. Effects of midazolam on the development of adult leydig cells from stem cells in vitro. Front Endocrinol. 2021, 12, 765251. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, F.; Ye, L.; Zirkin, B.; Chen, H. Steroidogenesis in Leydig cells: Effects of aging and environmental factors. Reproduction. 2017, 154, R111–R122. [Google Scholar] [CrossRef]
- Lavoie, H.A.; King, S.R. Transcriptional regulation of steroidogenic genes: STARD1, CYP11A1 and HSD3B. Exp. Biol. Med. 2009, 234, 880–907. [Google Scholar] [CrossRef] [PubMed]
- Noyola-Martínez, N.; Halhali, A.; Zaga-Clavellina, V.; Olmos-Ortiz, A.; Larrea, F.; Barrera, D. A time-course regulatory and kinetic expression study of steroid metabolizing enzymes by calcitriol in primary cultured human placental cells. J. Steroid Biochem. Mol. Biol. 2017, 167, 98–105. [Google Scholar] [CrossRef]
- Lu, X.; Ma, P.; Kong, L.; Wang, X.; Jiang, L. Vitamin K2 inhibits hepatocellular carcinoma cell proliferation by binding to 17β-hydroxysteroid dehydrogenase 4. Front. Oncol. 2021, 11, 757603. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Duan, A.; Ma, X.; Liang, S.; Deng, T. Knockdown of CYP19A1 in buffalo follicular granulosa cells results in increased progesterone secretion and promotes cell proliferation. Front. Vet. Sci. 2020, 7, 539496. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Huang, J.; Gu, X.; Li, L.; Han, J. Downregulation of aromatase plays a dual role in preeclampsia. Mol. Hum. Reprod. 2021, 27, gaab013. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Tasaki, T.; Tsukamoto, M.; Wang, K.Y.; Azuma, K. Deficiency of Wnt10a causes female infertility via the β-catenin/Cyp19a1 pathway in mice. Int. J. Med. Sci. 2022, 19, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Shu, C.; Han, S.; Xu, P.; Wang, Y.; Cheng, T.; Hu, C. Estrogen and preeclampsia: Potential of estrogens as therapeutic agents in preeclampsia. Drug Des. Devel. Ther. 2021, 15, 2543–2550. [Google Scholar] [CrossRef]
- Marquardt, R.M.; Kim, T.H.; Shin, J.H.; Jeong, J.W. Progesterone and Estrogen Signaling in the Endometrium: What Goes Wrong in Endometriosis? Int. J. Mol. Sci. 2019, 20, 3822. [Google Scholar] [CrossRef]
- Jo, Y.S.; Lee, G.S.; Nam, S.Y.; Kim, S.J. Progesterone Inhibits Leptin-Induced Invasiveness of BeWo Cells. Int. J. Mol. Sci. 2015, 12, 773–779. [Google Scholar] [CrossRef]
- Horita, K.; Inase, N.; Miyake, S.; Formby, B.; Toyoda, H.; Yoshizawa, Y. Progesterone induces apoptosis in malignant mesothelioma cells. Anticancer Res. 2001, 21, 3871–3874. [Google Scholar]
- Sugino, N.; Okuda, K. Species-related differences in the mechanism of apoptosis during structural luteolysis. J. Reprod. Dev. 2007, 53, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.M.; Cory, S. The Bcl-2 protein family: Arbiters of cell survival. Science 1998, 281, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- De Falco, M.; Fedele, V.; Cobellis, L.; Mastrogiacomo, A.; Giraldi, D.; Leone, S.; De Luca, L.; Laforgia, V.; De Luca, A. Pattern of expression of cyclin D1/CDK4 complex in human placenta during gestation. Cell Tissue Res. 2004, 317, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Nuzzo, A.M.; Giuffrida, D.; Masturzo, B.; Mele, P.; Piccoli, E.; Eva, C.; Todros, T.; Rolfo, A. Altered expression of G1/S phase cell cycle regulators in placental mesenchymal stromal cells derived from preeclamptic pregnancies with fetal-placental compromise. Cell Cycle 2017, 16, 200–212. [Google Scholar] [CrossRef]
- Zhou, Q.; Acharya, G. Editorial: Placental hormones and pregnancy-related endocrine disorders. Front Endocrinol. 2022, 13, 905829. [Google Scholar] [CrossRef]
- Alvarado-Flores, F.; Kaneko-Tarui, T.; Beyer, W.; Katz, J.; Chu, T.; Catalano, P.; Sadovsky, Y.; Hivert, M.F.; O’Tierney-Ginn, P. Placental miR-3940-3p is associated with maternal insulin resistance in late pregnancy. J. Clin. Endocrinol. Metab. 2021, 106, 3526–3535. [Google Scholar] [CrossRef]
- Shin, Y.Y.; An, S.M.; Jeong, J.S.; Yang, S.Y.; Lee, G.S.; Hong, E.J.; Jeung, E.B.; Kim, S.C.; An, B.S. Comparison of steroid hormones in three different preeclamptic models. Mol. Med. Rep. 2021, 23, 252. [Google Scholar] [CrossRef]
- Lawrenz, B.; Melado, L.; Fatemi, H. Premature progesterone rise in ART-cycles. Reprod. Biol. 2018, 18, 1–4. [Google Scholar] [CrossRef]
- Huang, B.; Ren, X.; Wu, L.; Zhu, L.; Xu, B.; Li, Y.; Ai, J.; Jin, L. Elevated Progesterone Levels on the Day of Oocyte Maturation May Affect Top Quality Embryo IVF Cycles. PLoS ONE 2016, 11, e0145895. [Google Scholar] [CrossRef]
- Adda-Herzog, E.; Poulain, M.; de Ziegler, D.; Ayoubi, J.M.; Fanchin, R. Premature progesterone elevation in controlled ovarian stimulation: To make a long story short. Fertil. Steril. 2018, 109, 563–570. [Google Scholar] [CrossRef]
- Kalakota, N.R.; George, L.C.; Morelli, S.S.; Douglas, N.C.; Babwah, A.V. Towards an Improved Understanding of the Effects of Elevated Progesterone Levels on Human Endometrial Receptivity and Oocyte/Embryo Quality during Assisted Reproductive Technologies. Cells 2022, 11, 1405. [Google Scholar] [CrossRef] [PubMed]
- Bazer, F.W.; Slayden, O.D. Progesterone-induced gene expression in uterine epithelia: A myth perpetuated by conventional wisdom. Biol. Reprod. 2008, 79, 1008–1009. [Google Scholar] [CrossRef] [PubMed]
- Diep, C.H.; Ahrendt, H.; Lange, C.A. Progesterone induces progesterone receptor gene (PGR) expression via rapid activation of protein kinase pathways required for cooperative estrogen receptor alpha (ER) and progesterone receptor (PR) genomic action at ER/PR target genes. Steroids 2016, 114, 48–58. [Google Scholar] [CrossRef] [PubMed]





| Antibody | Vendor | Code | Host Species | Dilution |
|---|---|---|---|---|
| CASP3 | Abclonal | A2156 | Rabbit | 1:1000 |
| Bax | Cell Signaling Technology | 2772T | Rabbit | 1:1000 |
| CCND2 | Proteintech | 10934-1-AP | Rabbit | 1:1000 |
| CYP11A1 | Absin | abs120402 | Rabbit | 1:1000 |
| CYP19A1 | Absin | 122200 | Rabbit | 1:1000 |
| StAR | Proteintech | 12225-1-AP | Rabbit | 1:1000 |
| GAPDH | Abclonal | AC002 | Mouse | 1:5000 |
| β–Actin | Proteintech | 66009-1 | Mouse | 1:5000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Ding, H.; Yang, Y.; Liu, Y.; Cao, X.; Feng, T. Progesterone Induces Apoptosis and Steroidogenesis in Porcine Placental Trophoblasts. Animals 2022, 12, 2704. https://doi.org/10.3390/ani12192704
Liu Y, Ding H, Yang Y, Liu Y, Cao X, Feng T. Progesterone Induces Apoptosis and Steroidogenesis in Porcine Placental Trophoblasts. Animals. 2022; 12(19):2704. https://doi.org/10.3390/ani12192704
Chicago/Turabian StyleLiu, Yueshuai, Hongxiang Ding, Yuze Yang, Yan Liu, Xin Cao, and Tao Feng. 2022. "Progesterone Induces Apoptosis and Steroidogenesis in Porcine Placental Trophoblasts" Animals 12, no. 19: 2704. https://doi.org/10.3390/ani12192704
APA StyleLiu, Y., Ding, H., Yang, Y., Liu, Y., Cao, X., & Feng, T. (2022). Progesterone Induces Apoptosis and Steroidogenesis in Porcine Placental Trophoblasts. Animals, 12(19), 2704. https://doi.org/10.3390/ani12192704