
Developmental Programming of Fertility in Cattle—Is It a Cause for Concern?


Abstract
Simple Summary
Abstract
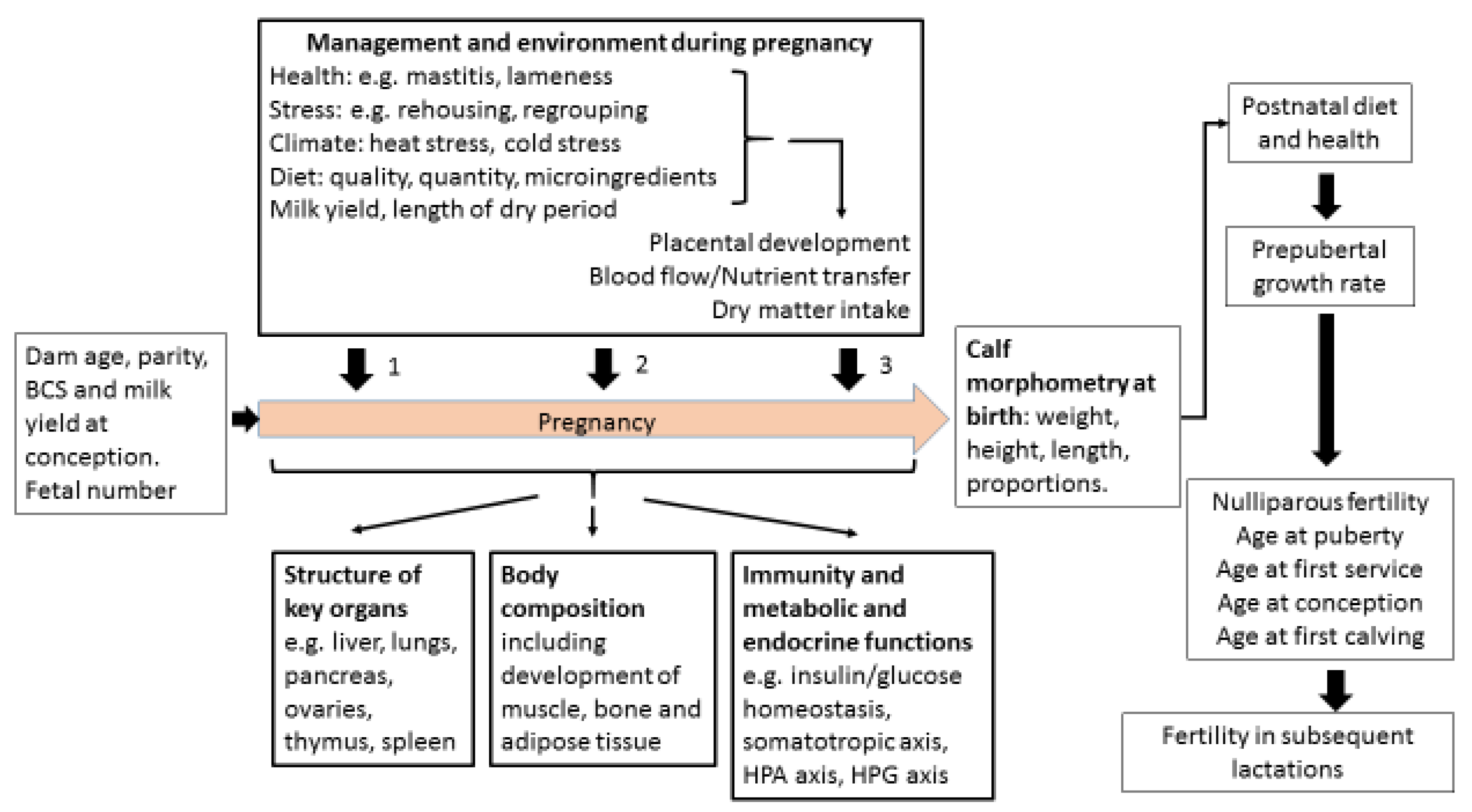
1. Introduction
2. Fetal Origins of Adult Disease
2.1. Background
2.2. Links to Fertility
2.3. Placental Development
2.4. Dam Parity and Milk Yield
2.5. Climate
3. Impact of Fetal Programming on Size at Birth and Post-Natal Growth
3.1. Size at Birth
3.2. Postnatal Growth
4. Influence of Fetal Programming on the Development of Specific Organs and Tissues
4.1. Muscle and Adipose Tissue
4.2. Skeleton
4.3. Ovaries
4.4. Gastrointestinal Tract and Lungs
4.5. Immune System
4.6. Anogenital Distance
5. Influence of Fetal Programming on Postnatal Metabolism and Endocrinology
5.1. Insulin and Glucose
5.2. Somatotropic Axis
5.3. Hypothalamic-Pituitary-Adrenal (HPA) Axis
5.4. Hypothalamic-Pituitary-Gonadal (HPG) Axis
6. Follow-Up Studies Investigating Fetal Programming and Fertility
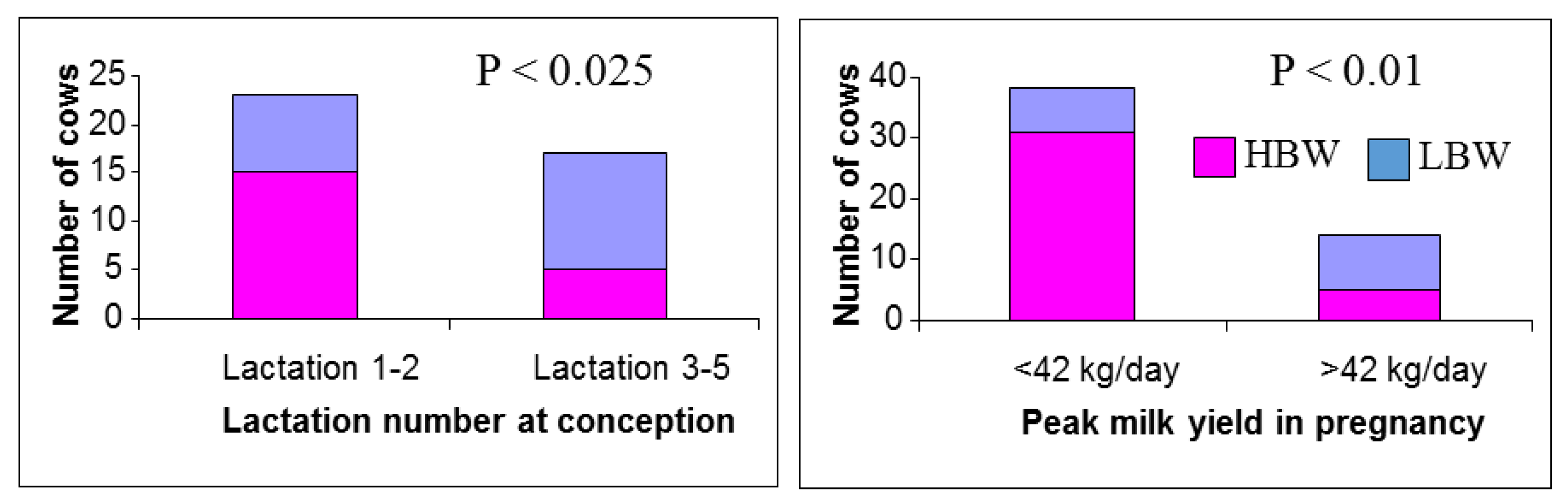
6.1. Dam Parity and Milk Yield
6.2. Dam Nutrition
6.3. Dam Heat Stress
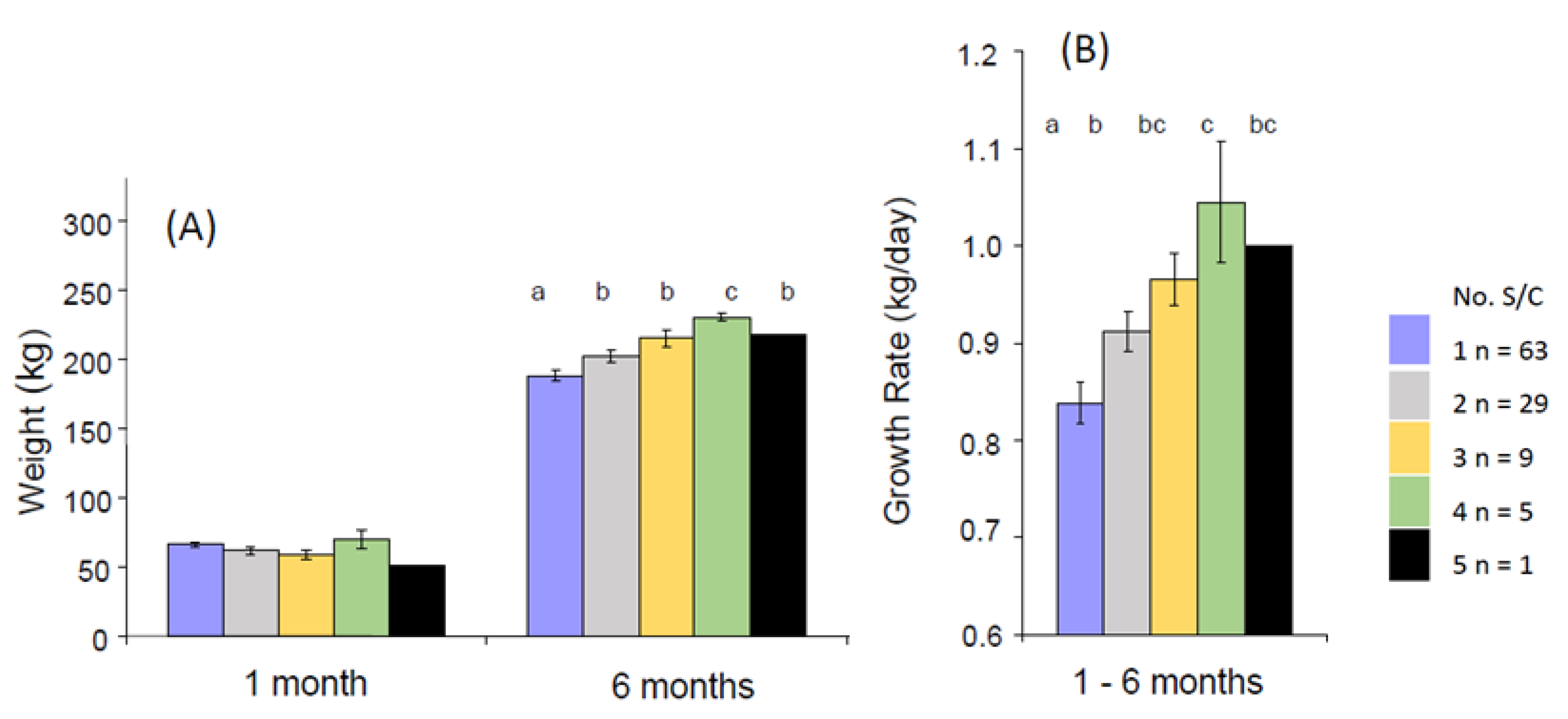
6.4. Calf Morphology
6.5. Anogenital Distance
6.6. Metabolic Imbalance
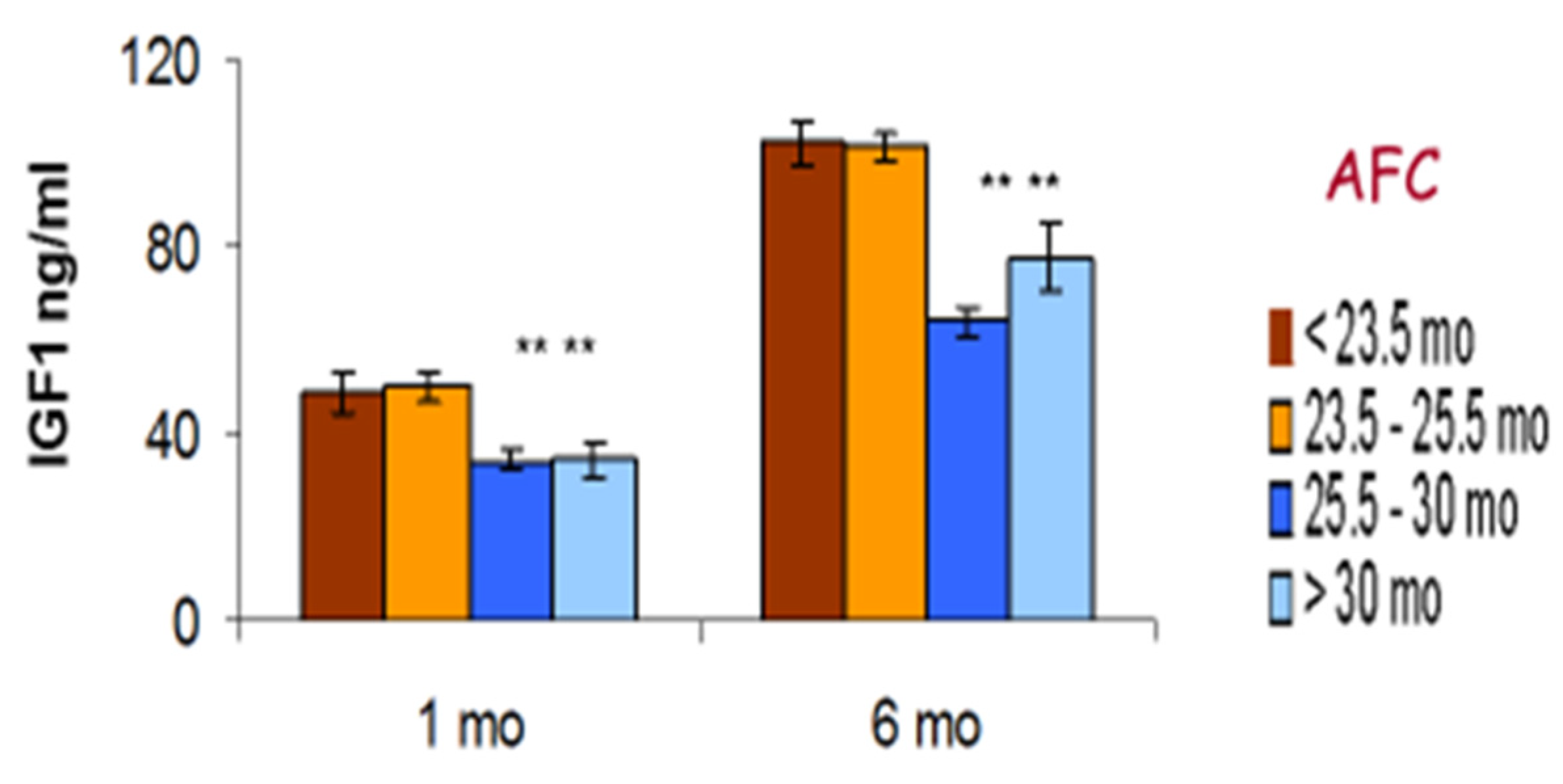
6.7. Somatotropic Axis
6.8. Disease
7. Epigenetic Modifications in Sperm
8. Assisted Reproductive Technologies
9. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. List of Abbreviations Used in the Text
References
- Barker, D.J. Fetal origins of coronary heart disease. BMJ 1995, 311, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Maloney, C.A.; Rees, W.D. Gene-nutrient interactions during fetal development. Reproduction 2005, 130, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.W.; Wakeland, A.K.; Parast, M.M. Trophoblast lineage specification, differentiation and their regulation by oxygen tension. J. Endocrinol. 2018, 236, R43–R56. [Google Scholar] [CrossRef]
- Leese, H.J.; Brison, D.R.; Sturmey, R.G. The quiet embryo hypothesis: 20 years on. Front. Physiol. 2022, 13, 899485. [Google Scholar] [CrossRef]
- Widdowson, E.M.; McCance, R.A. A review: New thoughts on growth. Pediatr. Res. 1975, 9, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.; Sinclair, K.D.; McEvoy, T.G. Nutritional effects on foetal growth. Anim. Sci. 1999, 68, 315–331. [Google Scholar] [CrossRef]
- Hales, C.N.; Barker, D.J. The thrifty phenotype hypothesis. Br. Med. Bull. 2001, 60, 5–20. [Google Scholar] [CrossRef]
- McMillen, I.C.; Adams, M.B.; Ross, J.T.; Coulter, C.L.; Simonetta, G.; Owens, J.A.; Robinson, J.S.; Edwards, L.J. Fetal growth restriction: Adaptations and consequences. Reproduction 2001, 122, 195–204. [Google Scholar] [CrossRef]
- Ozanne, S.E.; Fernandez-Twinn, D.; Hales, C.N. Fetal growth and adult diseases. Semin. Perinatol. 2004, 28, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Pinborg, A.; Loft, A.; Romundstad, L.B.; Wennerholm, U.B.; Söderström-Anttila, V.; Bergh, C.; Aittomäki, K. Epigenetics and assisted reproductive technologies. Acta Obstet. Gynecol. Scand. 2016, 95, 10–15. [Google Scholar] [CrossRef]
- Goyal, D.; Limesand, S.W.; Goyal, R. Epigenetic responses and the developmental origins of health and disease. J. Endocrinol. 2019, 242, T105–T119. [Google Scholar] [CrossRef]
- Morgan, H.L.; Watkins, A.J. Transgenerational impact of environmental change. Adv. Exp. Med. Biol. 2019, 1200, 71–89. [Google Scholar] [CrossRef]
- Yao, S.; Lopez-Tello, J.; Sferruzzi-Perri, A.N. Developmental programming of the female reproductive system-a review. Biol. Reprod. 2021, 104, 745–770. [Google Scholar] [CrossRef] [PubMed]
- Bauman, D.E.; Currie, W.B. Partitioning of nutrients during pregnancy and lactation—A review of mechanisms involving homeostasis and homeorhesis. J. Dairy Sci. 1980, 63, 1514–1529. [Google Scholar] [CrossRef]
- Ingvartsen, K.L.; Andersen, J.B. Integration of metabolism and intake regulation: A review focusing on periparturient animals. J. Dairy Sci. 2000, 83, 1573–1597. [Google Scholar] [CrossRef]
- Mellor, D.J. Nutritional and placental determinants of foetal growth rate in sheep and consequences for the newborn lamb. Br. Vet. J. 1983, 139, 307–324. [Google Scholar] [CrossRef]
- Brown, W.E.; Bradford, B.J. Invited review: Mechanisms of hypophagia during disease. J. Dairy Sci. 2021, 104, 9418–9436. [Google Scholar] [CrossRef]
- Greenwood, P.L.; Bell, A.W. Consequences of intra-uterine growth retardation for postnatal growth, metabolism and pathophysiology. Reprod. Suppl. 2003, 61, 195–206. [Google Scholar] [CrossRef][Green Version]
- Chavatte-Palmer, P.; Velazquez, M.A.; Jammes, H.; Duranthon, V. Review: Epigenetics, developmental programming and nutrition in herbivores. Animal 2018, 12, s363–s371. [Google Scholar] [CrossRef]
- Wallace, J.M. Competition for nutrients in pregnant adolescents: Consequences for maternal, conceptus and offspring endocrine systems. J. Endocrinol. 2019, 242, T1–T19. [Google Scholar] [CrossRef]
- Brickell, J.S.; Wathes, D.C. A descriptive study of the survival of Holstein-Friesian heifers through to third calving on English dairy farms. J. Dairy Sci. 2011, 94, 1831–1838. [Google Scholar] [CrossRef]
- Funston, R.N.; Larson, D.M.; Vonnahme, K.A. Effects of maternal nutrition on conceptus growth and offspring performance: Implications for beef cattle production. J. Anim. Sci. 2010, 88 (Suppl. 13), E205–E215. [Google Scholar] [CrossRef]
- Prior, R.L.; Laster, D.B. Development of the bovine fetus. J. Anim. Sci. 1979, 48, 1546–1553. [Google Scholar] [CrossRef] [PubMed]
- McCrabb, G.; Egan, A.; Hosking, B. Maternal undernutrition during mid-pregnancy in sheep: Variable effects on placental growth. J. Agric. Sci. 1992, 118, 127–132. [Google Scholar] [CrossRef]
- Osgerby, J.C.; Gadd, T.S.; Wathes, D.C. The effects of maternal nutrition and body condition on placental and foetal growth in the ewe. Placenta 2003, 24, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Heasman, L.; Clarke, L.; Stephenson, T.J.; Symonds, M.E. The influence of maternal nutrient restriction in early to mid-pregnancy on placental and fetal development in sheep. Proc. Nutr. Soc. 1999, 58, 283–288. [Google Scholar] [CrossRef]
- Wathes, D.C.; Reynolds, T.S.; Robinson, R.S.; Stevenson, K.R. Role of the insulin-like growth factor system in uterine function and placental development in ruminants. J. Dairy Sci. 1998, 81, 1778–1789. [Google Scholar] [CrossRef]
- Mericq, V.; Martinez-Aguayo, A.; Uauy, R.; Iñiguez, G.; Van der Steen, M.; Hokken-Koelega, A. Long-term metabolic risk among children born premature or small for gestational age. Nat. Rev. Endocrinol. 2017, 13, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Vonnahme, K.A.; Zhu, M.J.; Borowicz, P.P.; Geary, T.W.; Hess, B.W.; Reynolds, L.P.; Caton, J.S.; Means, W.J.; Ford, S.P. Effect of early gestational undernutrition on angiogenic factor expression and vascularity in the bovine placentome. J. Anim. Sci. 2007, 85, 2464–2472. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, L.P.; Borowicz, P.P.; Vonnahme, K.A.; Johnson, M.L.; Grazul-Bilska, A.T.; Wallace, J.M.; Caton, J.S.; Redmer, D.A. Animal models of placental angiogenesis. Placenta 2005, 26, 689–708. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.J.; Du, M.; Hess, B.W.; Means, W.J.; Nathanielsz, P.W.; Ford, S.P. Maternal nutrient restriction upregulates growth signaling pathway in the cotyledonary artery of cow placentomes. Placenta 2007, 28, 361–368. [Google Scholar] [CrossRef]
- Redifer, C.A.; Duncan, N.B.; Meyer, A.M. Factors affecting placental size in beef cattle: Maternal and fetal influences. Theriogenology 2021, 174, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, T.M.; Micke, G.C.; Magalhaes, R.S.; Phillips, N.J.; Perry, V.E. Dietary protein during gestation affects placental development in heifers. Theriogenology 2009, 72, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Kamal, M.M.; Van Eetvelde, M.; Vandaele, L.; Opsomer, G. Environmental and maternal factors associated with gross placental morphology in dairy cattle. Reprod. Domest. Anim. 2017, 52, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Wathes, D.C.; Brickell, J.S.; Bourne, N.E.; Swali, A.; Cheng, Z. Factors influencing heifer survival and fertility on commercial dairy farms. Animal 2008, 2, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Coffey, M.P.; Hickey, J.; Brotherstone, S. Genetic aspects of growth of Holstein-Friesian dairy cows from birth to maturity. J. Dairy Sci. 2006, 89, 322–329. [Google Scholar] [CrossRef]
- Frisancho, A.R.; Matos, J.; Leonard, W.R.; Yaroch, L.A. Developmental and nutritional determinants of pregnancy outcome among teenagers. Am. J. Phys. Anthropol. 1985, 66, 247–261. [Google Scholar] [CrossRef]
- Van Eetvelde, M.; Kamal, M.M.; Hostens, M.; Vandaele, L.; Fiems, L.O.; Opsomer, G. Evidence for placental compensation in cattle. Animal 2016, 10, 1342–1350. [Google Scholar] [CrossRef]
- Klewitz, J.; Struebing, C.; Rohn, K.; Goergens, A.; Martinsson, G.; Orgies, F.; Probst, J.; Hollinshead, F.; Bollwein, H.; Sieme, H. Effects of age, parity, and pregnancy abnormalities on foal birth weight and uterine blood flow in the mare. Theriogenology 2015, 83, 721–729. [Google Scholar] [CrossRef]
- Lundborg, G.K.; Oltenacu, P.A.; Maizon, D.O.; Svensson, E.C.; Liberg, P.G. Dam-related effects on heart girth at birth, morbidity and growth rate from birth to 90 days of age in Swedish dairy calves. Prev. Vet. Med. 2003, 60, 175–190. [Google Scholar] [CrossRef]
- Swali, A.; Wathes, D.C. Influence of primiparity on size at birth, growth, the somatotropic axis and fertility in dairy heifers. Anim. Reprod. Sci. 2007, 102, 122–136. [Google Scholar] [CrossRef] [PubMed]
- Akbarinejad, V.; Gharagozlou, F.; Vojgani, M.; Bagheri Amirabadi, M.B. Nulliparous and primiparous cows produce less fertile female offspring with lesser concentration of anti-Müllerian hormone (AMH) as compared with multiparous cows. Anim. Reprod. Sci. 2018, 197, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Kamal, M.M.; Van Eetvelde, M.; Depreester, E.; Hostens, M.; Vandaele, L.; Opsomer, G. Age at calving in heifers and level of milk production during gestation in cows are associated with the birth size of Holstein calves. J. Dairy Sci. 2014, 97, 5448–5458. [Google Scholar] [CrossRef] [PubMed]
- Swali, A.; Wathes, D.C. Influence of the dam and sire on size at birth and subsequent growth, milk production and fertility in dairy heifers. Theriogenology 2006, 66, 1173–1184. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.P.; Lee, J.M.; Macdonald, K.A.; Roche, J.R. Body condition score and body weight effects on dystocia and stillbirths and consequent effects on postcalving performance. J. Dairy Sci. 2007, 90, 4201–4211. [Google Scholar] [CrossRef] [PubMed]
- González-Recio, O.; Ugarte, E.; Bach, A. Trans-generational effect of maternal lactation during pregnancy: A Holstein cow model. PLoS ONE 2012, 7, e51816. [Google Scholar] [CrossRef]
- Banos, G.; Brotherstone, S.; Coffey, M.P. Prenatal maternal effects on body condition score, female fertility, and milk yield of dairy cows. J. Dairy Sci. 2007, 90, 3490–3499. [Google Scholar] [CrossRef]
- Monteiro, A.P.; Tao, S.; Thompson, I.M.; Dahl, G.E. Effect of heat stress during late gestation on immune function and growth performance of calves: Isolation of altered colostral and calf factors. J. Dairy Sci. 2014, 97, 6426–6439. [Google Scholar] [CrossRef]
- Tao, S.; Monteiro, A.P.; Thompson, I.M.; Hayen, M.J.; Dahl, G.E. Effect of late-gestation maternal heat stress on growth and immune function of dairy calves. J. Dairy Sci. 2012, 95, 7128–7136. [Google Scholar] [CrossRef]
- Ghaffari, M.H. Developmental programming: Prenatal and postnatal consequences of hyperthermia in dairy cows and calves. Domest. Anim. Endocrinol. 2022, 80, 106723. [Google Scholar] [CrossRef]
- Walton, A.; Hammond, J. The maternal effects on growth and conformation in shire horse-shetland pony crosses. Proc. R. Soc. Lond. 1938, 125, B125311–B125335. [Google Scholar] [CrossRef]
- Wilsher, S.; Allen, W.R. The effects of maternal age and parity on placental and fetal development in the mare. Equine Vet. J. 2003, 35, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M. Quantitative Genetic Analysis of Mortality in Danish Holstein Calves. Ph.D. Thesis, The Royal Veterinary and Agricultural University, Copenhagen, Denmark, 2004. [Google Scholar]
- Yin, T.; König, S. Genetic parameters for body weight from birth to calving and associations between weights with test-day, health, and female fertility traits. J. Dairy Sci. 2018, 101, 2158–2170. [Google Scholar] [CrossRef] [PubMed]
- Phocas, F.; Laloe, D. Genetic parameters for birth and weaning traits in French specialized beef cattle breeds. Livest. Prod. Sci. 2004, 89, 121–128. [Google Scholar] [CrossRef]
- Long, N.M.; Prado-Cooper, M.J.; Krehbiel, C.R.; Wettemann, R.P. Effects of nutrient restriction of bovine dams during early gestation on postnatal growth and regulation of plasma glucose. J. Anim. Sci. 2010, 88, 3262–3268. [Google Scholar] [CrossRef]
- Mossa, F.; Carter, F.; Walsh, S.W.; Kenny, D.A.; Smith, G.W.; Ireland, J.L.; Hildebrandt, T.B.; Lonergan, P.; Ireland, J.J.; Evans, A.C. Maternal undernutrition in cows impairs ovarian and cardiovascular systems in their offspring. Biol. Reprod. 2013, 88, 92. [Google Scholar] [CrossRef] [PubMed]
- Noya, A.; Casasús, I.; Ferrer, J.; Sanz, A. Long-term effects of maternal subnutrition in early pregnancy on cow-calf performance, immunological and physiological profiles during the next lactation. Animals 2019, 9, 936. [Google Scholar] [CrossRef]
- Micke, G.C.; Sullivan, T.M.; Soares Magalhaes, R.J.; Rolls, P.J.; Norman, S.T.; Perry, V.E.A. Heifer nutrition during early-and mid-pregnancy alters fetal growth trajectory and birth weight. Anim. Reprod. Sci. 2010, 117, 1–10. [Google Scholar] [CrossRef]
- Carstens, G.E.; Johnson, D.E.; Holland, M.D.; Odde, K.G. Effects of prepartum protein nutrition and birth weight on basal metabolism in bovine neonates. J. Anim. Sci. 1987, 65, 745–751. [Google Scholar] [CrossRef]
- Martin, J.L.; Vonnahme, K.A.; Adams, D.C.; Lardy, G.P.; Funston, R.N. Effects of dam nutrition on growth and reproductive performance of heifer calves. J. Anim. Sci. 2007, 85, 841–847. [Google Scholar] [CrossRef]
- Corah, L.R.; Dunn, T.G.; Kaltenbach, C.C. Influence of prepartum nutrition on the reproductive performance of beef females and the performance of their progeny. J. Anim. Sci. 1975, 41, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Long, J.M.; Trubenbach, L.A.; Hobbs, K.C.; Poletti, A.E.; Steinhauser, C.B.; Pryor, J.H.; Long, C.R.; Wickersham, T.A.; Sawyer, J.E.; Miller, R.K.; et al. Maternal nutrient restriction in late pregnancy programs postnatal metabolism and pituitary development in beef heifers. PLoS ONE 2021, 16, e0249924. [Google Scholar] [CrossRef] [PubMed]
- NRC. Nutrient Requirements of Beef Cattle, 7th ed.; Natl. Acad. Press: Washington, DC, USA, 2000. [Google Scholar]
- Cho, W.K.; Suh, B.K. Catch-up growth and catch-up fat in children born small for gestational age. Korean J. Pediatr. 2016, 59, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zanelli, S.A.; Rogol, A.D. Short children born small for gestational age outcomes in the era of growth hormone therapy. Growth Horm. IGF Res. 2018, 38, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Bieswal, F.; Ahn, M.T.; Reusens, B.; Holvoet, P.; Raes, M.; Rees, W.D.; Remacle, C. The importance of catch-up growth after early malnutrition for the programming of obesity in male rat. Obesity 2006, 14, 1330–1343. [Google Scholar] [CrossRef]
- Clayton, P.E.; Cianfarani, S.; Czernichow, P.; Johannsson, G.; Rapaport, R.; Rogol, A. Management of the child born small for gestational age through to adulthood: A consensus statement of the International Societies of Pediatric Endocrinology and the Growth Hormone Research Society. J. Clin. Endocrinol. Metab. 2007, 92, 804–810. [Google Scholar] [CrossRef]
- Gardner, D.S.; Tingey, K.; Van Bon, B.W.M.; Ozanne, S.E.; Wilson, V.; Dandrea, J.; Keisler, D.H.; Stephenson, T.; Symonds, M.E. Programming of glucose-insulin metabolism in adult sheep after maternal undernutrition. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R947–R954. [Google Scholar] [CrossRef]
- Du, M.; Tong, J.; Zhao, J.; Underwood, K.R.; Zhu, M.; Ford, S.P.; Nathanielsz, P.W. Fetal programming of skeletal muscle development in ruminant animals. J. Anim. Sci. 2010, 88 (Suppl. E), E51–E60. [Google Scholar] [CrossRef]
- Khanal, P.; Nielsen, M.O. Impacts of prenatal nutrition on animal production and performance: A focus on growth and metabolic and endocrine function in sheep. J. Anim. Sci. Biotechnol. 2017, 8, 75. [Google Scholar] [CrossRef]
- Hoffman, M.L.; Peck, K.N.; Forella, M.E.; Fox, A.R.; Govoni, K.E.; Zinn, S.A. The effects of poor maternal nutrition during gestation on postnatal growth and development of lambs. J. Anim. Sci. 2016, 94, 789–799. [Google Scholar] [CrossRef]
- Greenwood, P.L.; Hunt, A.S.; Hermanson, J.W.; Bell, A.W. Effects of birth weight and postnatal nutrition on neonatal sheep: II. Skeletal muscle growth and development. J. Anim. Sci. 2000, 78, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Symonds, M.E.; Stephenson, T. Maternal nutrition and endocrine programming of fetal adipose tissue development. Biochem. Soc. Trans. 1999, 27, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Ford, S.P.; Hess, B.W.; Schwope, M.M.; Nijland, M.J.; Gilbert, J.S.; Vonnahme, K.A.; Means, W.J.; Han, H.; Nathanielsz, P.W. Maternal undernutrition during early to mid-gestation in the ewe results in altered growth, adiposity, and glucose tolerance in male offspring. J. Anim. Sci. 2007, 85, 1285–1294. [Google Scholar] [CrossRef] [PubMed]
- Karunaratne, J.F.; Ashton, C.J.; Stickland, N.C. Fetal programming of fat and collagen in porcine skeletal muscles. J. Anat. 2005, 207, 763–768. [Google Scholar] [CrossRef]
- Manapurath, R.; Gadapani, B.; Pereira-da-Silva, L. Body composition of infants born with intrauterine growth restriction: A systematic review and meta-analysis. Nutrients 2022, 14, 1085. [Google Scholar] [CrossRef]
- Ibáñez, L.; Ong, K.; Dunger, D.B.; de Zegher, F. Early development of adiposity and insulin resistance after catch-up weight gain in small-for-gestational-age children. J. Clin. Endocrinol. Metab. 2006, 91, 2153–2158. [Google Scholar] [CrossRef]
- Osgerby, J.C.; Wathes, D.C.; Howard, D.; Gadd, T.S. The effect of maternal undernutrition on ovine fetal growth. J. Endocrinol. 2002, 173, 131–141. [Google Scholar] [CrossRef]
- Tygesen, M.P.; Harrison, A.P.; Therkildsen, M. The effect of maternal nutrition restriction during late gestation on muscle, bone and meat parameters in five month old lambs. Livest. Sci. 2007, 110, 230–241. [Google Scholar] [CrossRef]
- Heasman, L.; Clarke, L.; Firth, K.; Stephenson, T.; Symonds, M.E. Influence of restricted maternal nutrition in early to mid gestation on placental and fetal development at term in sheep. Pediatr. Res. 1998, 44, 546–551. [Google Scholar] [CrossRef]
- Sullivan, T.M.; Micke, G.C.; Perkins, N.; Martin, G.B.; Wallace, C.R.; Gatford, K.L.; Owens, J.A.; Perry, V.E. Dietary protein during gestation affects maternal insulin-like growth factor, insulin-like growth factor binding protein, leptin concentrations, and fetal growth in heifers. J. Anim. Sci. 2009, 87, 3304–3316. [Google Scholar] [CrossRef]
- Maresca, S.; Lopez Valiente, S.; Rodriguez, A.M.; Long, N.M.; Pavan, E.; Quintans, G. Effect of protein restriction of bovine dams during late gestation on offspring postnatal growth, glucose-insulin metabolism and IGF-1 concentration. Livest. Sci. 2018, 212, 120–126. [Google Scholar] [CrossRef]
- Clothier, J.; Small, A.; Hinch, G.; Brown, W.Y. Prematurity and dysmaturity are associated with reduced height and shorter distal limb length in horses. J. Equine Vet. Sci. 2020, 91, 103129. [Google Scholar] [CrossRef] [PubMed]
- Zuckerman, S. The number of oocytes in the mature ovary. Recent. Prog. Horm. Res. 1951, 6, 63–108. [Google Scholar]
- Borwick, S.C.; Rhind, S.M.; McMillen, S.R.; Racey, P.A. Effect of undernutrition of ewes from the time of mating on fetal ovarian development in mid gestation. Reprod. Fertil. Dev. 1997, 9, 711–715. [Google Scholar] [CrossRef]
- Rae, M.T.; Palassio, S.; Kyle, C.E.; Brooks, A.N.; Lea, R.G.; Miller, D.W.; Rhind, S.M. Effect of maternal undernutrition during pregnancy on early ovarian development and subsequent follicular development in sheep fetuses. Reproduction 2001, 122, 915–922. [Google Scholar] [CrossRef]
- Lea, R.G.; Andrade, L.P.; Rae, M.T.; Hannah, L.T.; Kyle, C.E.; Murray, J.F.; Rhind, S.M.; Miller, D.W. Effects of maternal undernutrition during early pregnancy on apoptosis regulators in the ovine fetal ovary. Reproduction 2006, 131, 113–124. [Google Scholar] [CrossRef][Green Version]
- Grazul-Bilska, A.T.; Caton, J.S.; Arndt, W.; Burchill, K.; Thorson, C.; Boroczyk, E.; Bilski, J.J.; Redmer, D.A.; Reynolds, L.P.; Vonnahme, K.A. Cellular proliferation and vascularization in ovine fetal ovaries: Effects of undernutrition and selenium in maternal diet. Reproduction 2009, 137, 699–707. [Google Scholar] [CrossRef]
- Nwachukwu, C.U.; Woad, K.J.; Barnes, N.; Gardner, D.S.; Robinson, R.S. Maternal protein restriction affects fetal ovary development in sheep. Reprod. Fertil. 2021, 2, 161–171. [Google Scholar] [CrossRef]
- Nilsson, E.E.; Skinner, M.K. Progesterone regulation of primordial follicle assembly in bovine fetal ovaries. Mol. Cell. Endocrinol. 2009, 313, 9–16. [Google Scholar] [CrossRef]
- Sullivan, T.M.; Micke, G.C.; Greer, R.M.; Irving-Rodgers, H.F.; Rodgers, R.J.; Perry, V.E. Dietary manipulation of Bos indicus x heifers during gestation affects the reproductive development of their heifer calves. Reprod. Fertil. Dev. 2009, 21, 773–784. [Google Scholar] [CrossRef]
- Ireland, J.J.; Zielak-Steciwko, A.E.; Jimenez-Krassel, F.; Folger, J.; Bettegowda, A.; Scheetz, D.; Walsh, S.; Mossa, F.; Knight, P.G.; Smith, G.W.; et al. Variation in the ovarian reserve is linked to alterations in intrafollicular estradiol production and ovarian biomarkers of follicular differentiation and oocyte quality in cattle. Biol. Reprod. 2009, 80, 954–964. [Google Scholar] [CrossRef] [PubMed]
- Mossa, F.; Walsh, S.W.; Butler, S.T.; Berry, D.P.; Carter, F.; Lonergan, P.; Smith, G.W.; Ireland, J.J.; Evans, A.C. Low numbers of ovarian follicles ≥3 mm in diameter are associated with low fertility in dairy cows. J. Dairy Sci. 2012, 95, 2355–2361. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, E.S.; Bisinotto, R.S.; Lima, F.S.; Greco, L.F.; Morrison, A.; Kumar, A.; Thatcher, W.W.; Santos, J.E. Plasma anti-Müllerian hormone in adult dairy cows and associations with fertility. J. Dairy Sci. 2014, 97, 6888–6900. [Google Scholar] [CrossRef] [PubMed]
- Cushman, R.A.; Allan, M.F.; Kuehn, L.A.; Snelling, W.M.; Cupp, A.S.; Freetly, H.C. Evaluation of antral follicle count and ovarian morphology in crossbred beef cows: Investigation of influence of stage of the estrous cycle, age, and birth weight. J. Anim. Sci. 2009, 87, 1971–1980. [Google Scholar] [CrossRef]
- Tenley, S.C.; Gomes, R.S.; Rosasco, S.L.; Northrop, E.J.; Rich, J.J.; McNeel, A.K.; Summers, A.F.; Miles, J.R.; Chase, C.C., Jr.; Lents, C.A.; et al. Maternal age influences the number of primordial follicles in the ovaries of yearling Angus heifers. Anim. Reprod. Sci. 2019, 200, 105–112. [Google Scholar] [CrossRef]
- Gobikrushanth, M.; Purfield, D.C.; Colazo, M.G.; Butler, S.T.; Wang, Z.; Ambrose, D.J. The relationship between serum anti-Müllerian hormone concentrations and fertility, and genome-wide associations for anti-Müllerian hormone in Holstein cows. J. Dairy Sci. 2018, 101, 7563–7574. [Google Scholar] [CrossRef]
- Avila, C.G.; Harding, R.; Rees, S.; Robinson, P.M. Small intestinal development in growth-retarded fetal sheep. J. Pediatr. Gastroenterol. Nutr. 1989, 8, 507–515. [Google Scholar] [CrossRef]
- McMullen, S.; Osgerby, J.C.; Milne, J.S.; Wallace, J.M.; Wathes, D.C. The effects of acute nutrient restriction in the mid-gestational ewe on maternal and fetal nutrient status, the expression of placental growth factors and fetal growth. Placenta 2005, 26, 25–33. [Google Scholar] [CrossRef]
- McMillen, I.C.; Robinson, J.S. Developmental origins of the metabolic syndrome: Prediction, plasticity, and programming. Physiol. Rev. 2005, 85, 571–633. [Google Scholar] [CrossRef]
- Alexander, G. Birth weight of lambs: Influences and consequences. In Size at Birth; Elliott, K., Knight., J., Eds.; Elsevier: Amsterdam, The Netherlands, 1974; pp. 213–239. [Google Scholar]
- Clarke, L.; Firth, K.; Heasman, L.; Juniper, D.T.; Budge, H.; Stephenson, T.; Symonds, M.E. Influence of relative size at birth on growth and glucose homeostasis in twin lambs during juvenile life. Reprod. Fertil. Dev. 2000, 12, 69–73. [Google Scholar] [CrossRef]
- Hyatt, M.A.; Budge, H.; Walker, D.; Stephenson, T.; Symonds, M.E. Effects of maternal parity and late gestational nutrition on mRNA abundance for growth factors in the liver of postnatal sheep. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R1934–R1942. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, M.A.; Gopalakrishnan, G.S.; Bispham, J.; Gentili, S.; McMillen, I.C.; Rhind, S.M.; Rae, M.T.; Kyle, C.E.; Brooks, A.N.; Jones, C.; et al. Maternal nutrient restriction in early pregnancy programs hepatic mRNA expression of growth-related genes and liver size in adult male sheep. J. Endocrinol. 2007, 192, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Fenwick, M.A.; Fitzpatrick, R.; Kenny, D.A.; Diskin, M.G.; Patton, J.; Murphy, J.J.; Wathes, D.C. Interrelationships between negative energy balance (NEB) and IGF regulation in liver of lactating dairy cows. Domest. Anim. Endocrinol. 2008, 34, 31–44. [Google Scholar] [CrossRef]
- Desai, M.; Hales, C.N. Role of fetal and infant growth in programming metabolism in later life. Biol. Rev. Camb. Philos. Soc. 1997, 72, 329–348. [Google Scholar] [CrossRef] [PubMed]
- Harding, R.; Hooper, S.B. Regulation of lung expansion and lung growth before birth. J. Appl. Physiol. 1996, 81, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Sherer, D.M.; Davis, J.M.; Woods, J.R., Jr. Pulmonary hypoplasia: A review. Obstet. Gynecol. Surv. 1990, 45, 792–803. [Google Scholar]
- Harding, R.; Higgins, G.C. The influence of oligohydramnios on thoracic dimensions in the sheep. J. Dev. Physiol. 1991, 16, 355–361. [Google Scholar]
- Jakubowska, A.E.; Billings, K.; Johns, D.P.; Hooper, S.B.; Harding, R. Respiratory function in lambs following prolonged oligohydramnios during late gestation. Pediatr. Res. 1993, 34, 611–617. [Google Scholar] [CrossRef]
- Wright, R.J. Perinatal stress and early life programming of lung structure and function. Biol. Psychol. 2010, 84, 46–56. [Google Scholar] [CrossRef]
- Joss-Moore, L.A.; Albertine, K.H.; Lane, R.H. Epigenetics and the developmental origins of lung disease. Mol. Genet. Metab. 2011, 104, 61–66. [Google Scholar] [CrossRef]
- Harding, R.; Cock, M.L.; Louey, S.; Joyce, B.J.; Davey, M.G.; Albuquerque, C.A.; Hooper, S.B.; Maritz, G.S. The compromised intra-uterine environment: Implications for future lung health. Clin. Exp. Pharmacol. Physiol. 2000, 27, 965–974. [Google Scholar] [CrossRef]
- Long, N.M.; Prado-Cooper, M.J.; Krehbiel, C.R.; DeSilva, U.; Wettemann, R.P. Effects of nutrient restriction of bovine dams during early gestation on postnatal growth, carcass and organ characteristics, and gene expression in adipose tissue and muscle. J. Anim. Sci. 2010, 88, 3251–3261. [Google Scholar] [CrossRef] [PubMed]
- Bronte, V.; Pittet, M.J. The spleen in local and systemic regulation of immunity. Immunity 2013, 39, 806–818. [Google Scholar] [CrossRef] [PubMed]
- Gasisova, A.I.; Atkenova, A.B.; Ahmetzhanova, N.B.; Murzabekova, L.M.; Bekenova, A.C. Morphostructure of immune system organs in cattle of different age. Anat. Histol. Embryol. 2017, 46, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Armengaud, J.B.; Dennebouy, Z.; Labes, D.; Fumey, C.; Wilson, A.; Candotti, F.; Yzydorczyk, C.; Simeoni, U. Intra-uterine growth restriction induced by maternal low-protein diet causes long-term alterations of thymic structure and function in adult male rat offspring. Br. J. Nutr. 2020, 123, 892–900. [Google Scholar] [CrossRef]
- Cromi, A.; Ghezzi, F.; Raffaelli, R.; Bergamini, V.; Siesto, G.; Bolis, P. Ultrasonographic measurement of thymus size in IUGR fetuses: A marker of the fetal immunoendocrine response to malnutrition. Ultrasound Obstet. Gynecol. 2009, 33, 421–426. [Google Scholar] [CrossRef]
- Fulford, A.J.; Moore, S.E.; Arifeen, S.E.; Persson, L.Å.; Neufeld, L.M.; Wagatsuma, Y.; Prentice, A.M. Disproportionate early fetal growth predicts postnatal thymic size in humans. J. Dev. Orig. Health Dis. 2013, 4, 223–231. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, F.; Nie, H.; Ma, T.; Wang, Z.; Wang, F.; Loor, J.J. Dietary N-carbamylglutamate and rumen-protected L-arginine supplementation during intrauterine growth restriction in undernourished ewes improve fetal thymus development and immune function. Reprod. Fertil. Dev. 2018, 30, 1522–1531. [Google Scholar] [CrossRef]
- Ahmed, B.M.S.; Younas, U.; Asar, T.O.; Monteiro, A.P.A.; Hayen, M.J.; Tao, S.; Dahl, G.E. Maternal heat stress reduces body and organ growth in calves: Relationship to immune status. JDS Commun. 2021, 2, 295–299. [Google Scholar] [CrossRef]
- Marrero, M.G.; Dado-Senn, B.; Field, S.L.; Yang, G.; Driver, J.P.; Laporta, J. Chronic heat stress delays immune system development and alters serotonin signaling in pre-weaned dairy calves. PLoS ONE 2021, 16, e0252474. [Google Scholar] [CrossRef]
- Stylianopoulou, F. Effect of maternal adrenocorticotropin injections on the differentiation of sexual behavior of the offspring. Horm. Behav. 1983, 17, 324–331. [Google Scholar] [CrossRef]
- Wolf, C.J.; Hotchkiss, A.; Ostby, J.S.; LeBlanc, G.A.; Gray, L.E., Jr. Effects of prenatal testosterone propionate on the sexual development of male and female rats: A dose-response study. Toxicol. Sci. 2002, 65, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Perlman, S.; Toledano, Y.; Kivilevitch, Z.; Halevy, N.; Rubin, E.; Gilboa, Y. Foetal sonographic anogenital distance is longer in polycystic ovary syndrome mothers. J. Clin. Med. 2020, 9, 2863. [Google Scholar] [CrossRef]
- Wooding, F.B.; Fowden, A.L.; Bell, A.W.; Ehrhardt, R.A.; Limesand, S.W.; Hay, W.W. Localisation of glucose transport in the ruminant placenta: Implications for sequential use of transporter isoforms. Placenta 2005, 26, 626–640. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.Q.; Keating, A.F. Expression and regulation of glucose transporters in the bovine mammary gland. J. Dairy Sci. 2007, 90 (Suppl. 1), E76–E86. [Google Scholar] [CrossRef] [PubMed]
- De Koster, J.D.; Opsomer, G. Insulin resistance in dairy cows. Vet. Clin. North Am. Food Anim. Pract. 2013, 29, 299–322. [Google Scholar] [CrossRef] [PubMed]
- Fowden, A.L.; Hill, D.J. Intra-uterine programming of the endocrine pancreas. Br. Med. Bull. 2001, 60, 123–142. [Google Scholar] [CrossRef] [PubMed]
- Odhiambo, J.F.; Pankey, C.L.; Ghnenis, A.B.; Ford, S.P. A review of maternal nutrition during pregnancy and impact on the offspring through development: Evidence from animal models of over- and undernutrition. Int. J. Environ. Res. Public Health 2020, 17, 6926. [Google Scholar] [CrossRef]
- Peterson, M.; Gauvin, M.; Pillai, S.; Jones, A.; McFadden, K.; Cameron, K.; Reed, S.; Zinn, S.; Govoni, K. Maternal under- and over-nutrition during gestation causes islet hypertrophy and sex-specific changes to pancreas DNA methylation in fetal sheep. Animals 2021, 11, 2531. [Google Scholar] [CrossRef]
- Carver, T.D.; Anderson, S.M.; Aldoretta, P.W.; Hay, W.W.J. Effect of low-level basal plus marked “pulsatile” hyperglycemia on insulin secretion in fetal sheep. Am. J. Physiol. Endocrinol. Metab. 1996, 271, E865–E871. [Google Scholar] [CrossRef]
- Long, J.M.; Trubenbach, L.A.; Pryor, J.H.; Long, C.R.; Wickersham, T.A.; Sawyer, J.E.; Satterfield, M.C. Maternal nutrient restriction alters endocrine pancreas development in fetal heifers. Domest. Anim. Endocrinol. 2021, 74, 106580. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, P.; Hunt, A.; Slepetis, R.; Finnerty, K.; Alston, C.; Beermann, D.; Bell, A.W. Effects of birth weight and postnatal nutrition on neonatal sheep. III. Regulation of energy metabolism. J. Anim. Sci. 2002, 80, 2850–2861. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, M.L.; Peck, K.N.; Wegrzyn, J.L.; Reed, S.A.; Zinn, S.A.; Govoni, K.E. Poor maternal nutrition during gestation alters the expression of genes involved in muscle development and metabolism in lambs. J. Anim. Sci. 2016, 94, 3093–3099. [Google Scholar] [CrossRef] [PubMed]
- Kamal, M.M.; Van Eetvelde, M.; Bogaert, H.; Hostens, M.; Vandaele, L.; Shamsuddin, M.; Opsomer, G. Environmental factors and dam characteristics associated with insulin sensitivity and insulin secretion in newborn Holstein calves. Animal 2015, 9, 1490–1499. [Google Scholar] [CrossRef] [PubMed]
- Van Eetvelde, M.; Opsomer, G. Innovative look at dairy heifer rearing: Effect of prenatal and post-natal environment on later performance. Reprod. Domest. Anim. 2017, 52 (Suppl. 3), 30–36. [Google Scholar] [CrossRef]
- Clark, R. The somatogenic hormones and insulin-like growth factor-1: Stimulators of lymphopoiesis and immune function. Endocr. Rev. 1997, 18, 157–179. [Google Scholar] [CrossRef]
- Heemskerk, V.H.; Daemen, M.A.; Buurman, W.A. Insulin-like growth factor-1 (IGF-1) and growth hormone (GH) in immunity and inflammation. Cytokine Growth Factor Rev. 1999, 10, 5–14. [Google Scholar] [CrossRef]
- Le Roith, D.; Bondy, C.; Yakar, S.; Liu, J.L.; Butler, A. The somatomedin hypothesis: 2001. Endocr. Rev. 2001, 22, 53–74. [Google Scholar] [CrossRef]
- Butler, S.T.; Marr, A.L.; Pelton, S.H.; Radcliff, R.P.; Lucy, M.C.; Butler, W.R. Insulin restores GH responsiveness during lactation-induced negative energy balance in dairy cattle: Effects on expression of IGF-I and GH receptor 1A. J. Endocrinol. 2003, 176, 205–217. [Google Scholar] [CrossRef]
- Taylor, V.J.; Cheng, Z.; Pushpakumara, P.G.; Beever, D.E.; Wathes, D.C. Relationships between the plasma concentrations of insulin-like growth factor-I in dairy cows and their fertility and milk yield. Vet. Rec. 2004, 155, 583–588. [Google Scholar] [CrossRef]
- Li, J.; Owens, J.A.; Owens, P.C.; Saunders, J.C.; Fowden, A.L.; Gilmour, R.S. The ontogeny of hepatic growth hormone receptor and insulin-like growth factor I gene expression in the sheep fetus during late gestation: Developmental regulation by cortisol. Endocrinology 1996, 137, 1650–1657. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, M.A.; Budge, H.; Walker, D.; Stephenson, T.; Symonds, M.E. Ontogeny and nutritional programming of the hepatic growth hormone-insulin-like growth factor-prolactin axis in the sheep. Endocrinology 2007, 148, 4754–4760. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hammon, H.M.; Zbinden, Y.; Sauerwein, H.; Breier, B.H.; Blum, J.W.; Donkin, S.S. The response of the hepatic insulin-like growth factor system to growth hormone and dexamethasone in calves. J. Endocrinol. 2003, 179, 427–435. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gluckman, P.D.; Sizonenko, S.V.; Bassett, N.S. The transition from fetus to neonate--an endocrine perspective. Acta Paediatr. Suppl. 1999, 88, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, R.P.; Greenwood, P.L.; Bell, A.W.; Boisclair, Y.R. Nutritional regulation of the genes encoding the acid-labile subunit and other components of the circulating insulin-like growth factor system in the sheep. J. Anim. Sci. 2000, 78, 2681–2689. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.K.; Breier, B.H.; Harding, J.E.; Veldhuis, J.D.; Gluckman, P.D. The fetal somatotropic axis during long term maternal undernutrition in sheep: Evidence for nutritional regulation in utero. Endocrinology 1995, 136, 1250–1257. [Google Scholar] [CrossRef]
- Gallaher, B.W.; Breier, B.H.; Keven, C.L.; Harding, J.E.; Gluckman, P.D. Fetal programming of insulin-like growth factor (IGF)-I and IGF-binding protein-3: Evidence for an altered response to undernutrition in late gestation following exposure to periconceptual undernutrition in the sheep. J. Endocrinol. 1998, 159, 501–508. [Google Scholar] [CrossRef]
- Heasman, L.; Brameld, J.; Mostvn, A.; Budge, H.; Dawson, J.; Buttery, P.; Stephenson, T.; Symonds, M.E. Maternal nutrient restriction during early to mid gestation alters the relationship between insulin-like growth factor I and bodyweight at term in fetal sheep. Reprod. Fertil. Dev. 2000, 12, 345–350. [Google Scholar] [CrossRef]
- Owens, J.A.; Kind, K.L.; Carbone, F.; Robinson, J.S.; Owens, P.C. Circulating insulin-like growth factors-I and -II and substrates in fetal sheep following restriction of placental growth. J. Endocrinol. 1994, 140, 5–13. [Google Scholar] [CrossRef]
- Holland, M.D.; Hossner, K.L.; Williams, S.E.; Wallace, C.R.; Niswender, G.D.; Odde, K.G. Serum concentrations of insulin-like growth factors and placental lactogen during gestation in cattle. I. Fetal profiles. Domest. Anim. Endocrinol. 1997, 14, 231–239. [Google Scholar] [CrossRef]
- Brickell, J.S.; McGowan, M.M.; Wathes, D.C. Effect of management factors and blood metabolites during the rearing period on growth in dairy heifers on UK farms. Domest. Anim. Endocrinol. 2009, 36, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.F.; Chancellor, N.; Burn, C.C.; Wathes, D.C. Analysis of pre-weaning feeding policies and other risk factors influencing growth rates in calves on 11 commercial dairy farms. Animal 2018, 12, 1413–1423. [Google Scholar] [CrossRef] [PubMed]
- Fowden, A.L.; Vaughan, O.R.; Murray, A.J.; Forhead, A.J. Metabolic consequences of glucocorticoid exposure before birth. Nutrients 2022, 14, 2304. [Google Scholar] [CrossRef] [PubMed]
- Edwards, L.J.; McMillen, I.C. Impact of maternal undernutrition during the periconceptional period, fetal number, and fetal sex on the development of the hypothalamo-pituitary adrenal axis in sheep during late gestation. Biol. Reprod. 2002, 66, 1562–1569. [Google Scholar] [CrossRef] [PubMed]
- Huber, E.; Notaro, U.S.; Recce, S.; Rodríguez, F.M.; Ortega, H.H.; Salvetti, N.R.; Rey, F. Fetal programming in dairy cows: Effect of heat stress on progeny fertility and associations with the hypothalamic-pituitary-adrenal axis functions. Anim. Reprod. Sci. 2020, 216, 106348. [Google Scholar] [CrossRef]
- Zambrano, E.; Guzmán, C.; Rodríguez-González, G.L.; Durand-Carbajal, M.; Nathanielsz, P.W. Fetal programming of sexual development and reproductive function. Mol. Cell. Endocrinol. 2014, 382, 538–549. [Google Scholar] [CrossRef]
- Bloomfield, F.H.; Oliver, M.H.; Hawkins, P.; Holloway, A.C.; Campbell, M.; Gluckman, P.D.; Harding, J.E.; Challis, J.R. Periconceptional undernutrition in sheep accelerates maturation of the fetal hypo-thalamic–pituitary–adrenal axis in late gestation. Endocrinology 2004, 145, 4278–4285. [Google Scholar] [CrossRef]
- Gardner, D.S.; Van Bon, B.W.; Dandrea, J.; Goddard, P.J.; May, S.F.; Wilson, V.; Stephenson, T.; Symonds, M.E. Effect of periconceptional undernutrition and gender on hypothalamic-pituitary-adrenal axis function in young adult sheep. J. Endocrinol. 2006, 190, 203–212. [Google Scholar] [CrossRef]
- Hawkins, P.; Steyn, C.; McGarrigle, H.H.G.; Calder, N.A.; Saito, T.; Stratford, L.L.; Noakes, D.E.; Hanson, M.A. Cardiovascular and hypothalamic–pituitary axis development in late gestation fetal sheep and young lambs following modest maternal nutrient restriction in early gestation. Reprod. Fertil. Dev. 2000, 12, 443–456. [Google Scholar] [CrossRef]
- Chadio, S.E.; Kotsampasi, B.; Papadomichelakis, G.; Deligeorgis, S.; Kalogiannis, D.; Menegatos, I.; Zervas, G. Impact of maternal undernutrition on the hypothalamic pituitary-adrenal axis responsiveness in sheep at different ages postnatal. J. Endocrinol. 2007, 192, 495–503. [Google Scholar] [CrossRef]
- Wallace, J.M.; Milne, J.S.; Green, L.R.; Aitken, R.P. Postnatal hypothalamic-pituitary-adrenal function in sheep is influenced by age and sex, but not by prenatal growth restriction. Reprod. Fertil Dev. 2011, 23, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Borwick, S.C.; Rae, M.T.; Brooks, J.; McNeilly, A.S.; Racey, P.A.; Rhind, S.M. Undernutrition of ewe lambs in utero and in early post-natal life does not affect hypothalamic-pituitary function in adulthood. Anim. Reprod. Sci. 2003, 77, 61–70. [Google Scholar] [CrossRef]
- Kotsampasi, B.; Chadio, S.; Papadomichelakis, G.; Deligeorgis, S.; Kalogiannis, D.; Menegatos, I.; Zervas, G. Effects of maternal undernutrition on the hypothalamic-pituitary-gonadal axis function in female sheep offspring. Reprod. Domest. Anim. 2009, 44, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Pryce, J.E.; Coffey, M.P.; Brotherstone, S.H.; Woolliams, J.A. Genetic relationships between calving interval and body condition score conditional on milk yield. J. Dairy Sci. 2002, 85, 1590–1595. [Google Scholar] [CrossRef]
- Gunn, R.G.; Sim, D.A.; Hunter, E.A. Effects of nutrition in utero and in early life on the subsequent lifetime reproductive performance of Scottish Blackface ewes in two management systems. Anim. Sci. 1995, 60, 223–230. [Google Scholar] [CrossRef]
- Rae, M.T.; Kyle, C.E.; Miller, D.W.; Hammond, A.J.; Brooks, A.N.; Rhind, S.M. The effects of undernutrition, in utero, on reproductive function in adult male and female sheep. Anim. Reprod. Sci. 2002, 72, 63–71. [Google Scholar] [CrossRef]
- Long, N.M.; Nijland, M.J.; Nathanielsz, P.W.; Ford, S.P. The effect of early to mid-gestational nutrient restriction on female offspring fertility and hypothalamic-pituitary-adrenal axis response to stress. J. Anim. Sci. 2010, 88, 2029–2037. [Google Scholar] [CrossRef]
- Funston, R.N.; Martin, J.L.; Adams, D.C.; Larson, D.M. Effects of winter grazing system and supplementation during late gestation on performance of beef cows and progeny. Proc. West. Sec. Am. Soc. Anim. Sci. 2008, 59, 102–105. [Google Scholar]
- Kipp, C.; Brügemann, K.; Zieger, P.; Mütze, K.; Möcklinghoff-Wicke, S.; König, S.; Halli, K. Across-generation effects of maternal heat stress during late gestation on production, female fertility and longevity traits in dairy cows. J. Dairy Res. 2021, 88, 147–153. [Google Scholar] [CrossRef]
- Succu, S.; Sale, S.; Ghirello, G.; Ireland, J.J.; Evans, A.C.O.; Atzori, A.S.; Mossa, F. Exposure of dairy cows to high environmental temperatures and their lactation status impairs establishment of the ovarian reserve in their offspring. J. Dairy Sci. 2020, 103, 11957–11969. [Google Scholar] [CrossRef]
- Nogalski, Z.; Mordas, W. Pelvic parameters in Holstein-Friesian and Jersey heifers in relation to their calving. Pak. Vet. J. 2012, 32, 507–510. [Google Scholar]
- Wall, E.; White, I.M.; Coffey, M.P.; Brotherstone, S. The relationship between fertility, rump angle, and selected type information in Holstein-Friesian cows. J. Dairy Sci. 2005, 88, 1521–1528. [Google Scholar] [CrossRef]
- Van Eetvelde, M.; Verdru, K.; de Jong, G.; van Pelt, M.L.; Meesters, M.; Opsomer, G. Researching 100 t cows: An innovative approach to identify intrinsic cow factors associated with a high lifetime milk production. Prev. Vet. Med. 2021, 193, 105392. [Google Scholar] [CrossRef] [PubMed]
- Nogalski, Z. Effect of selected factors on the course of parturition in Holstein-Friesian heifers. EJPAU 2002, 5, #03. Available online: http://www.ejpau.media.pl/volume5/issue2/animal/art-03.html (accessed on 26 September 2022).
- Heinrichs, A.J.; Zanton, G.I.; Lascano, G.J.; Jones, C.M. A 100-Year Review: A century of dairy heifer research. J. Dairy Sci. 2017, 100, 10173–10188. [Google Scholar] [CrossRef]
- Mee, J.F. Prevalence and risk factors for dystocia in dairy cattle: A review. Vet. J. 2008, 176, 93–101. [Google Scholar] [CrossRef]
- Wathes, D.C.; Bourne, N.; Cheng, Z.; Mann, G.E.; Taylor, V.J.; Coffey, M.P. Multiple correlation analyses of metabolic and endocrine profiles with fertility in primiparous and multiparous cows. J. Dairy Sci. 2007, 90, 1310–1325. [Google Scholar] [CrossRef]
- Swali, A.; Cheng, Z.; Bourne, N.; Wathes, D.C. Metabolic traits affecting growth rates of pre-pubertal calves and their relationship with subsequent survival. Domest. Anim. Endocrinol. 2008, 35, 300–313. [Google Scholar] [CrossRef]
- Taylor, V.J.; Beever, D.E.; Bryant, M.J.; Wathes, D.C. First lactation ovarian function in dairy heifers in relation to prepubertal metabolic profiles. J. Endocrinol. 2004, 180, 63–75. [Google Scholar] [CrossRef]
- Velazquez, M.A.; Spicer, L.J.; Wathes, D.C. The role of endocrine insulin-like growth factor-I (IGF-I) in female bovine reproduction. Domest. Anim. Endocrinol. 2008, 35, 325–342. [Google Scholar] [CrossRef]
- Brickell, J.S.; Bourne, N.; McGowan, M.M.; Wathes, D.C. Effect of growth and development during the rearing period on the subsequent fertility of nulliparous Holstein-Friesian heifers. Theriogenology 2009, 72, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Gardner, R.W.; Schuh, J.D.; Vargus, L.G. Accelerated growth and early breeding of Holstein heifers. J. Dairy Sci. 1977, 60, 1941–1948. [Google Scholar] [CrossRef]
- Abeni, F.; Calamari, L.; Stefanini, L.; Pirlo, G. Effects of daily gain in pre- and postpubertal replacement dairy heifers on body condition score, body size, metabolic profile, and future milk production. J. Dairy Sci. 2000, 83, 1468–1478. [Google Scholar] [CrossRef]
- Cabrera, V.E.; Fricke, P.M. Economics of Twin Pregnancies in Dairy Cattle. Animas 2021, 11, 552. [Google Scholar] [CrossRef] [PubMed]
- Kohler, H.-P.; Knudsen, L.B.; Skytthe, A.; Christensen, K. The fertility pattern of twins and the general population compared: Evidence from Danish cohorts 1945-64. Demogr. Res. 2002, 6, 383–408. [Google Scholar] [CrossRef]
- Kasimanickam, R.K.; Kasimanickam, V.R.; McCann, M.L. Difference in body weight at breeding affects reproductive performance in replacement beef heifers and carries consequences to next generation heifers. Animals 2021, 11, 2800. [Google Scholar] [CrossRef]
- Gobikrushanth, M.; Bruinjé, T.C.; Colazo, M.G.; Butler, S.T.; Ambrose, D.J. Characterization of anogenital distance and its relationship to fertility in lactating Holstein cows. J. Dairy Sci. 2017, 100, 9815–9823. [Google Scholar] [CrossRef]
- Carrelli, J.E.; Gobikrushanth, M.; Corpron, M.; Rajesh, I.; Sandberg, W.; Colazo, M.G.; Ahmadzadeh, A.; Oba, M.; Ambrose, D.J. Relationship of anogenital distance with fertility in nulliparous Holstein heifers. J. Dairy Sci. 2021, 104, 8256–8264. [Google Scholar] [CrossRef]
- Carrelli, J.E.; Gobikrushanth, M.; Corpron, M.; Sandberg, W.; Rajesh, I.; Ahmadzadeh, A.; Oba, M.; Ambrose, D.J. Associations between anogenital distance and measures of fertility in lactating North American Holstein cows: A validation study. J. Dairy Sci. 2022, 105, 6339–6352. [Google Scholar] [CrossRef]
- Gobikrushanth, M.; Purfield, D.C.; Kenneally, J.; Doyle, R.C.; Holden, S.A.; Martinez, P.M.; Canadas, E.R.; Bruinjé, T.C.; Colazo, M.G.; Ambrose, D.J.; et al. The relationship between anogenital distance and fertility, and genome-wide associations for anogenital distance in Irish Holstein-Friesian cows. J. Dairy Sci. 2019, 102, 1702–1711. [Google Scholar] [CrossRef]
- Rajesh, I.; Gobikrushanth, M.; Carrelli, J.E.; Oba, M.; Ambrose, D.J. Repeatability of anogenital distance measurements from birth to maturity and at different physio logical states in female Holstein cattle. J. Dairy Sci. 2022, 105, 2699–2707. [Google Scholar] [CrossRef] [PubMed]
- Grala, T.M.; Price, M.D.; Kuhn-Sherlock, B.; Burke, C.R.; Meier, S. Investigating anogenital distance and antral follicle count as novel markers of fertility within a herd of cows with positive or negative genetic merit for fertility traits. J. Dairy Sci. 2021, 104, 12939–12952. [Google Scholar] [CrossRef] [PubMed]
- Lucy, M.C. Mechanisms linking nutrition and reproduction in postpartum cows. Reprod. Suppl. 2003, 61, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Chagas, L.M.; Bass, J.J.; Blache, D.; Burke, C.R.; Kay, J.K.; Lindsay, D.R.; Lucy, M.C.; Martin, G.B.; Meier, S.; Rhodes, F.M.; et al. Invited review: New perspectives on the roles of nutrition and metabolic priorities in the subfertility of high-producing dairy cows. J. Dairy Sci. 2007, 90, 4022–4032. [Google Scholar] [CrossRef]
- Wathes, D.C. Mechanisms linking metabolic status and disease with reproductive outcome in the dairy cow. Reprod. Domest. Anim. 2012, 47 (Suppl. 4), 304–312. [Google Scholar] [CrossRef]
- Pascottini, O.B.; Leroy, J.L.; Opsomer, G. Maladaptation to the transition period and consequences on fertility of dairy cows. Reprod. Domest. Anim. 2022, 57, 21–32. [Google Scholar] [CrossRef]
- Smith, J.M.; Van Amburgh, M.E.; Diaz, M.C.; Lucy, M.C.; Bauman, D.E. Effect of nutrient intake on the development of the somatotropic axis and its responsiveness to GH in Holstein bull calves. J. Anim. Sci. 2002, 80, 1528–1537. [Google Scholar] [CrossRef]
- Terré, M.; Devant, M.; Bach, A. Performance and nitrogen metabolism of calves fed conventionally or following an enhanced-growth feeding program during the preweaning period. Livest. Sci. 2006, 105, 109–119. [Google Scholar] [CrossRef]
- Butler, W.R. Nutritional interactions with reproductive performance in dairy cattle. Anim. Reprod. Sci. 2000, 60, 44–9457. [Google Scholar] [CrossRef]
- Gobikrushanth, M.; Purfield, D.C.; Colazo, M.G.; Wang, Z.; Butler, S.T.; Ambrose, D.J. The relationship between serum insulin-like growth factor-1 (IGF-1) concentration and reproductive performance, and genome-wide associations for serum IGF-1 in Holstein cows. J. Dairy Sci. 2018, 101, 9154–9167. [Google Scholar] [CrossRef]
- Rhoads, R.P.; Kim, J.W.; Leury, B.J.; Baumgard, L.H.; Segoale, N.; Frank, S.J.; Bauman, D.E.; Boisclair, Y.R. Insulin increases the abundance of the growth hormone receptor in liver and adipose tissue of periparturient dairy cows. J. Nutr. 2004, 134, 1020–1027. [Google Scholar] [CrossRef] [PubMed]
- Taylor, V.J.; Beever, D.E.; Bryant, M.J.; Wathes, D.C. Pre-pubertal measurements of the somatotropic axis as predictors of milk production in Holstein-Friesian dairy cows. Domest. Anim. Endocrinol. 2006, 31, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Givens, M.D.; Marley, M.S. Infectious causes of embryonic and fetal mortality. Theriogenology 2008, 70, 270–285. [Google Scholar] [CrossRef]
- Kossaibati, M.A.; Esslemont, R.J. The costs of production diseases in dairy herds in England. Vet. J. 1997, 154, 41–51. [Google Scholar] [CrossRef]
- Cheng, Z.; Buggiotti, L.; Salavati, M.; Marchitelli, C.; Palma-Vera, S.; Wylie, A.; Takeda, H.; Tang, L.; Crowe, M.A.; Wathes, D.C. GplusE consortium. Global transcriptomic profiles of circulating leucocytes in early lactation cows with clinical or subclinical mastitis. Mol. Biol. Rep. 2021, 48, 4611–4623. [Google Scholar] [CrossRef] [PubMed]
- Windeyer, M.C.; Leslie, K.E.; Godden, S.M.; Hodgins, D.C.; Lissemore, K.D.; LeBlanc, S.J. Factors associated with morbidity, mortality, and growth of dairy heifer calves up to 3 months of age. Prev. Vet. Med. 2014, 113, 231–240. [Google Scholar] [CrossRef]
- Paré, J.; Thurmond, M.C.; Gardner, I.A.; Picanso, J.P. Effect of birthweight, total protein, serum IgG and packed cell volume on risk of neonatal diarrhea in calves on two California dairies. Can. J. Vet. Res. 1993, 57, 241–246. [Google Scholar] [PubMed]
- McCorquodale, C.E.; Sewalem, A.; Miglior, F.; Kelton, D.; Robinson, A.; Koeck, A.; Leslie, K.E. Short communication: Analysis of health and survival in a population of Ontario Holstein heifer calves. J. Dairy Sci. 2013, 96, 1880–1885. [Google Scholar] [CrossRef]
- Glover, I.D.; Barrett, D.C.; Reyher, K.K. Little association between birth weight and health of preweaned dairy calves. Vet. Rec. 2019, 184, 477. [Google Scholar] [CrossRef]
- Johnson, K.F.; Chancellor, N.; Wathes, D.C. A cohort study risk factor analysis for endemic disease in pre-weaned dairy heifer calves. Animals 2021, 11, 378. [Google Scholar] [CrossRef]
- Brickell, J.S.; McGowan, M.M.; Pfeiffer, D.U.; Wathes, D.C. Mortality in Holstein-Friesian calves and replacement heifers, in relation to body weight and IGF-I concentration, on 19 farms in England. Animal 2009, 3, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Haisan, J.; Oba, M.; Ambrose, D.J.; Steele, M.A. Short communication: The effects of offering a high or low plane of milk preweaning on insulin-like growth factor and insulin-like growth factor binding proteins in dairy heifer calves. J. Dairy Sci. 2018, 101, 11441–11446. [Google Scholar] [CrossRef] [PubMed]
- Donovan, G.A.; Dohoo, I.R.; Montgomery, D.M.; Bennett, F.L. Calf and disease factors affecting growth in female Holstein calves in Florida, USA. Prev. Vet. Med. 1998, 33, 1–10. [Google Scholar] [CrossRef]
- Quigley, J.D.; Wolfe, T.A.; Elsasser, T.H. Effects of additional milk replacer feeding on calf health, growth, and selected blood metabolites in calves. J. Dairy Sci. 2006, 89, 207–216. [Google Scholar] [CrossRef]
- Fox, C.J.; Hammerman, P.S.; Thompson, C.B. Fuel feeds function: Energy metabolism and the T-cell response. Nat. Rev. Immunol. 2005, 5, 844–852. [Google Scholar] [CrossRef] [PubMed]
- Hurst, T.S.; Neves, R.C.; Boerman, J.P. Early life indicators of first lactation milk yield and the effect of treatment for bovine respiratory disease on survivability and risk of pregnancy in Holstein dairy cattle. Vet. J. 2022, 282, 105826. [Google Scholar] [CrossRef] [PubMed]
- Correa, M.T.; Curtis, C.R.; Erb, H.N.; White, M.E. Effect of calfhood morbidity on age at first calving in New York Holstein herds. Prev. Vet. Med. 1988, 6, 253–262. [Google Scholar] [CrossRef]
- Heinrichs, A.J.; Heinrichs, B.S.; Harel, O.; Rogers, G.W.; Place, N.T. A prospective study of calf factors affecting age, body size, and body condition score at first calving of Holstein dairy heifers. J. Dairy Sci. 2005, 88, 2828–2835. [Google Scholar] [CrossRef]
- Bach, A. Associations between several aspects of heifer development and dairy cow survivability to second lactation. J. Dairy Sci. 2011, 94, 1052–1057. [Google Scholar] [CrossRef]
- Svensson, C.; Hultgren, J. Associations between housing, management, and morbidity during rearing and subsequent first-lactation milk production of dairy cows in southwest Sweden. J. Dairy Sci. 2008, 91, 1510–1518. [Google Scholar] [CrossRef]
- Dahlen, C.R.; Borowicz, P.P.; Ward, A.K.; Caton, J.S.; Czernik, M.; Palazzese, L.; Loi, P.; Reynolds, L.P. Programming of embryonic development. Int. J. Mol. Sci. 2021, 22, 11668. [Google Scholar] [CrossRef] [PubMed]
- Veenendaal, M.V.; Painter, R.C.; de Rooij, S.R.; Bossuyt, P.M.; van der Post, J.A.; Gluckman, P.D.; Hanson, M.A.; Roseboom, T.J. Transgenerational effects of prenatal exposure to the 1944-45 Dutch famine. BJOG 2013, 120, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Carone, B.R.; Fauquier, L.; Habib, N.; Shea, J.M.; Hart, C.E.; Li, R.; Bock, C.; Li, C.; Gu, H.; Zamore, P.D.; et al. Paternally induced transgenerational environmental reprogramming of metabolic gene expression in mammals. Cell 2010, 143, 1084–1096. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.B.; Kamal, M.M.; Rijsselaere, T.; Vandaele, L.; Shamsuddin, M.; Van Soom, A. Altered chromatin condensation of heat-stressed spermatozoa perturbs the dynamics of DNA methylation reprogramming in the paternal genome after in vitro fertilisation in cattle. Reprod. Fertil. Dev. 2014, 26, 1107–1116. [Google Scholar] [CrossRef]
- Bonilla, L.; Block, J.; Denicol, A.C.; Hansen, P.J. Consequences of transfer of an in vitro-produced embryo for the dam and resultant calf. J. Dairy Sci. 2014, 97, 229–239. [Google Scholar] [CrossRef]
- Mullaart, E.; Ludema, G.; Zijlstra, A.; Veldhuisen, J. No difference in health and fertility characteristics between offspring arising from in vitro production or multiple ovulation embryo transfer. Reprod. Fertil. Dev. 2021, 34, 267–268. [Google Scholar] [CrossRef]
- Rabaglino, M.B.; Bojsen-Møller Secher, J.; Sirard, M.-A.; Hyttel, P.; Kadarmideen, H.N. Epigenomic and transcriptomic analyses reveal early activation of the HPG axis in in vitro-produced male dairy calves. FASEB J. 2021, 35, e21882. [Google Scholar] [CrossRef]
- Rabaglino, M.B.; Secher, J.B.; Hyttel, P.; Kadarmideen, H.N. In vitro and in vivo-produced male dairy calves show molecular differences in the hepatic and muscular energy regulation. Biol. Reprod. 2022, ioac131. [Google Scholar] [CrossRef]
- Hagan, B.A.; Moro-Mendez, J.; Cue, R.I. Realized genetic selection differentials in Canadian Holstein dairy herds. J. Dairy Sci. 2020, 103, 1651–1666. [Google Scholar] [CrossRef]
- Brito, L.F.; Bedere, N.; Douhard, F.; Oliveira, H.R.; Arnal, M.; Peñagaricano, F.; Schinckel, A.P.; Baes, C.F.; Miglior, F. Review: Genetic selection of high-yielding dairy cattle toward sustainable farming systems in a rapidly changing world. Animal 2021, 15 (Suppl. 1), 100292. [Google Scholar] [CrossRef]
- Calus, M.P.; Bijma, P.; Veerkamp, R.F. Evaluation of genomic selection for replacement strategies using selection index theory. J. Dairy Sci. 2015, 98, 6499–6509. [Google Scholar] [CrossRef] [PubMed]
- Newton, J.E.; Hayes, B.J.; Pryce, J.E. The cost-benefit of genomic testing of heifers and using sexed semen in pasture-based dairy herds. J. Dairy Sci. 2018, 101, 6159–6173. [Google Scholar] [CrossRef] [PubMed]
- Dallago, G.M.; Wade, K.M.; Cue, R.I.; McClure, J.T.; Lacroix, R.; Pellerin, D.; Vasseur, E. Keeping dairy cows for longer: A critical literature review on dairy cow longevity in high milk-producing countries. Animals 2021, 11, 808. [Google Scholar] [CrossRef] [PubMed]
- Boulton, A.C.; Rushton, J.; Wathes, D.C. An empirical analysis of the cost of rearing dairy heifers from birth to first calving and the time taken to repay these costs. Animal 2017, 11, 1372–1380. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dam Previous 305 d Milk Yield (kg) | <7500 | 7500–10,000 | >10,000 | p 1 |
|---|---|---|---|---|
| n | 98 | 166 | 85 | |
| Age at first service (d) | 536 ± 18.3 a | 461 ± 6.8 b | 500 ± 12.4 a | 0.000 |
| Age at conception (d) | 558 ± 16.5 a | 513 ± 9.0 b | 541 ± 17.5 a | 0.034 |
| Age at first calving (d) | 839 ± 16.5 a | 791 ± 9.0 b | 828 ± 17.9 a | 0.018 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wathes, D.C. Developmental Programming of Fertility in Cattle—Is It a Cause for Concern? Animals 2022, 12, 2654. https://doi.org/10.3390/ani12192654
Wathes DC. Developmental Programming of Fertility in Cattle—Is It a Cause for Concern? Animals. 2022; 12(19):2654. https://doi.org/10.3390/ani12192654
Chicago/Turabian StyleWathes, D. Claire. 2022. "Developmental Programming of Fertility in Cattle—Is It a Cause for Concern?" Animals 12, no. 19: 2654. https://doi.org/10.3390/ani12192654
APA StyleWathes, D. C. (2022). Developmental Programming of Fertility in Cattle—Is It a Cause for Concern? Animals, 12(19), 2654. https://doi.org/10.3390/ani12192654