
Morpho-Functional Traits in Pura Raza Menorquina Horses: Genetic Parameters and Relationship with Coat Color Variables

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
3. Results and Discussion
3.1. Descriptive Statistics of the Data
3.2. Genetic Parameters
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Folla, F.; Sartori, C.; Mancin, E.; Pigozzi, G.; Mantovani, R. Genetic Parameters of Linear Type Traits Scored at 30 Months in Italian Heavy Draught Horse. Animals 2020, 10, 1099. [Google Scholar] [CrossRef] [PubMed]
- Gómez; Molina, A.; Sánchez-Guerrero, M.; Valera, M. Prediction of adult conformation traits from shape characteristics of Pura Raza Español foals. Livest. Sci. 2021, 253, 104701. [Google Scholar] [CrossRef]
- Forest, A. Son importance avant l’achat en fonction d’un service recherché et pour le choix des reproducteurs. In La Conformation d’un Cheval, Proceedings of the 7th Colloque sur le Cheval, Saint Hyacinthe, QC, Canada, 27 April 1996; Conseil des Productions Animales du Québec (CPAQ): Quebec City, QC, Canada, 1996; pp. 83–93. [Google Scholar]
- Mawdsley, A.; Kelly, E.P.; Smith, F.H.; Brophy, P.O. Linear assessment of the Thoroughbred horse: An approach to conformation evaluation. Equine Vet.-J. 1996, 28, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Bruns, E.; Bierbaum, M.; Frese, D.; Haring, H.J.F. Die Entwicklung von Selektionskriterienfür die Reitpferdezucht. IV. Schätzungrelativerökonomischer Gewichtanhand von Auktionsergebnissen. Züchtungskunde 1978, 50, 93–100. [Google Scholar]
- Schwark, H.J.; Petzold, P.; Nörenberg, I. UntersuchungenzurAuswahl von Selektionskriterienbei der Weiterentwicklung der Pferdezucht der DDR. Arch. Tierz. 1988, 31, 279–289. [Google Scholar]
- Sánchez-Guerrero, M.J.; Negro-Rama, S.; Demyda-Peyras, S.; Solé-Berga, M.; Azor-Ortiz, P.J.; Valera-Córdoba, M. Morphological and genetic diversity of Pura Raza Español horse with regard to the coal colour. Anim. Sci. J. 2019, 90, 14–22. [Google Scholar] [CrossRef]
- Pruvost, M.; Bellone, R.; Benecke, N.; Sandoval-Castellanos, E.; Cieslak, M.; Kuznetsova, T.; Morales-Muñiz, A.; O’Connor, T.; Reissmann, M.; Hofreiter, M.; et al. Genotypes of predomestic horses match phenotypes painted in Paleolithic works of cave art. Proc. Natl. Acad. Sci. USA 2011, 108, 18626–18630. [Google Scholar] [CrossRef] [PubMed]
- Novotná, A.; Svitáková, A.; Veselá, Z.; Vostrý, L. Estimation of genetic parameters for linear type traits in the population of sport horses in the Czech Republic. Livest. Sci. 2017, 202, 1–6. [Google Scholar] [CrossRef]
- Koenen, E.; van Veldhuizen, A.; Brascamp, E. Genetic parameters of linear scored conformation traits and their relation to dressage and show-jumping performance in the Dutch Warmblood Riding Horse population. Livest. Prod. Sci. 1995, 43, 85–94. [Google Scholar] [CrossRef]
- Schroderus, E.; Ojala, M. Estimates of genetic parameters for conformation measures and scores in Finnhorse and Standardbred foals. J. Anim. Breed. Genet. 2010, 127, 395–403. [Google Scholar] [CrossRef]
- Solé, M.; Cervantes, I.; Gutiérrez, J.P.; Gomez, M.D.; Valera, M. Estimation of genetic parameters for morphological and functional traits in a Menorca horse population. Span. J. Agric. Res. 2014, 12, 125. [Google Scholar] [CrossRef]
- MAPA. Census of Pura Raza Menorquina Horses. 2022. Available online: https://www.mapa.gob.es/es/ganaderia/temas/zootecnia/razas-ganaderas/razas/catalogo-razas/equino-caballar/menorquina/default.aspx (accessed on 15 March 2022).
- MAPA. Purebred Menorca Horse Breeding Program. 2022. Available online: https://www.mapa.gob.es/es/ganaderia/temas/zootecnia/en_programa_de_cria_prme_definitivo_tcm30-561612.PDF (accessed on 15 March 2022).
- Giontella, A.; Sarti, F.M.; Biggio, G.P.; Giovannini, S.; Cherchi, R.; Pieramati, C.; Silvestrelli, M. Genetic Parameters and Inbreeding Effect of Morphological Traits in Sardinian Anglo Arab Horse. Animals 2020, 10, 791. [Google Scholar] [CrossRef] [PubMed]
- Hofmanova, B.; Kohoutova, P.; Vostrý, L.; Vydrova, H.V.; Majzlik, I. Quantitative aspects of coat color in old kladruber black horses. Poljoprivreda 2015, 21, 224–227. [Google Scholar] [CrossRef]
- SAS, Statistical Analysis Software. Users’ Guide Statistics Version 9.4; SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Misztal, I.; Tsuruta, S.; Lourenco, D.; Aguilar, I.; Legarra, A.; Vitezica, Z. Manual for BLUPF90 Family of Programs; University of Georgia: Athens, GA, USA, 2018; p. 142. [Google Scholar]
- Plummer, M.; Best, N.; Cowles, K.; Vines, K. CODA: Convergence diagnosis and output analysis for MCMC. R News. 2006, 6, 7–11. [Google Scholar]
- Gutierrez, J.P.; Goyache, F. A note on ENDOG: A computer program for analysing pedigree information. J. Anim. Breed. Genet. 2005, 122, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Kuhnke, S.; Bär, K.; Bosch, P.; Rensing, M.; König, U.; Borstel, V. Evaluation of a novel system for linear conformation, gait and personality trait scoring and automatic ranking of horses at breed shows: A pilot study in America Quarter Horses. J. Equine Vet. Sci. 2019, 78, 53–59. [Google Scholar] [CrossRef]
- Vostrý, L.; Capkova, Z.; Andrejsová, L.; Mach, K.; Majzlík, I. Linear type trait analysis in the Coldblood breeds: Czech-Moravian Belgian horse and Silesian Noriker. Slovak J. Anim. Sci. 2009, 42, 99–106. [Google Scholar]
- Jakubec, V.; Pejfková, M.; Volenec, J.; Majzlík, I.; Vostrý, L. Analysis of linear description of type traits in the varieties and studs of the Old Kladrub horse. Czech J. Anim. Sci. 2007, 52, 299–307. [Google Scholar] [CrossRef]
- Jakubec, V.; Vostrý, L.; Schlote, W.; Majzlík, I.; Mach, K. Selection in the genetic resource: Genetic variation of the linear described type traits in the Old Kladrub horse. Arch. Anim. Breed. 2009, 52, 343–355. [Google Scholar] [CrossRef]
- Sánchez, M.; Gómez, M.; Molina, A.; Valera, M. Genetic analyses for linear conformation traits in Pura Raza Español horses. Livest. Sci. 2013, 157, 57–64. [Google Scholar] [CrossRef]
- Sánchez, M.J.; Gómez, M.D.; Molina, A.; Valera, M. Assessment scores in morphological competitions of Pura Raza Español Horse. Int. J. Agric. Biol. 2014, 16, 557–563. [Google Scholar]
- Sánchez, M.J.; Azor, P.J.; Molina, A.; Parkin, T.; Rivero, J.L.L.; Valera, M. Prevalence, risk factors and genetic parameters of cresty neck in Pura Raza Español horses. Equine Vet. J. 2017, 49, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Bowling, A.T. Dominant Inheritance of Overo Spotting in Paint Horses. J. Hered. 1994, 85, 222–224. [Google Scholar] [CrossRef] [PubMed]
- Corbi-Botto, C.M.; Sadaba, S.A.; Francisco, E.I.; Kalemkerian, P.B.; Lirón, J.P.; Villegas-Castagnasso, E.E.; Giovambattista, G.; Peral-García, P.; Díaz, S. Genetic variability of Appaloosa horses: A study of a closed breeding population from Argentina. Front. Agric. Sci. Eng. 2014, 1, 175–178. [Google Scholar] [CrossRef]
- Sponenberg, D.P. Equine Color Genetics; A Blackwell Publishing Company: Hoboken, NJ, USA; Iowa State Press: Ames, IA, USA, 1996. [Google Scholar]
- Hofmanová, B.; Vostrý, L.; Vostrá-Vydrová, H.; Dokoupilová, A.; Majzlík, I. Estimation of genetic and non-genetic effects in-fluencing coat color in black horses. Czech J. Anim. Sci. 2019, 64, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Pasternak, M.; Krupiński, J.; Gurgul, A.; Bugno-Poniewierska, M. Genetic, historical and breeding aspects of the occurrence of the tobiano pattern and white markings in the Polish population of Hucul horses—A review. J. Appl. Anim. Res. 2020, 48, 21–27. [Google Scholar] [CrossRef]
- Grilz-Seger, G.; Dobretsberger, M.; Brem, G.; Druml, T. Study on distribution of coat color related alleles and white markings in a closed population of Lipizzan horses. Zuchtungskunde 2020, 92, 76–86. [Google Scholar]
- Rieder, S.; Hagger, C.; Obexer-Ruff, G.; Leeb, T.; Poncet, P.-A. Genetic Analysis of White Facial and Leg Markings in the Swiss Franches-Montagnes Horse Breed. J. Hered. 2008, 99, 130–136. [Google Scholar] [CrossRef]
- Woolf, C.M. Multifactorial Inheritance of Common White Markings in the Arabian Horse. J. Hered. 1990, 81, 250–256. [Google Scholar] [CrossRef]
- Stachurska, A.; Ussing, A.P. White markings in horses. Med. Wet. 2012, 68, 74–78. [Google Scholar]
- Fegraeus, K.J.; Velie, B.D.; Axelsson, J.; Ang, R.; Hamilton, N.A.; Andersson, L.; Meadows, J.; Lindgren, G. A potential regulatory region near the EDN3 gene may control both harness racing performance and coat color variation in horses. Physiol. Rep. 2018, 6, e13700. [Google Scholar] [CrossRef]
- Bellone, R.R. Pleiotropic effects of pigmentation genes in horses. Anim. Genet. 2010, 41, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Azeem, N.M.; Emeash, H.H. Relationship of horse temperament with breed, age, sex and body characteristics: A questionnaire-based study. J. Basic Appl. 2021, 10, 45. [Google Scholar] [CrossRef]
- Vostrý, L.; Mach, K.; Přibyl, J. Selection of a suitable data set and model for the genetic evaluation of the linear description of conformation and type description in Old Kladruber horses. Arch. Anim. Breed. 2012, 55, 105–112. [Google Scholar] [CrossRef]
- Rustin, M.; Janssens, S.; Buys, N.; Gengler, N. Multi-trait animal model estimation of genetic parameters for linear type and gait traits in the Belgian warmblood horse. J. Anim. Breed. Genet. 2009, 126, 378–386. [Google Scholar] [CrossRef]
- Ripolles, M.; Sánchez-Guerrero, M.; Perdomo-González, D.; Azor, P.; Valera, M. Survey of Risk Factors and Genetic Characterization of Ewe Neck in a World Population of Pura Raza Español Horses. Animals 2020, 10, 1789. [Google Scholar] [CrossRef]
- Ducro, B.J.; Bovenhuis, H.; Back, W. Heritability of foot conformation and its relationship to sports performance in a Dutch Warmblood horse population. Equine Vet. J. 2009, 41, 139–143. [Google Scholar] [CrossRef]
- Finn, J.L.; Haase, B.; Willet, C.E.; van Rooy, D.; Chew, T.; Wade, C.M.; Hamilton, N.A.; Velie, B.D. The relationship between coat colour phenotype ad equine behavior: A pilot study. Appl. Anim. Behav. Sci. 2016, 174, 66–69. [Google Scholar] [CrossRef]
- Jacobs, L.N.; Staiger, E.A.; Albright, J.D.; Brooks, S. Data from: The MC1R and ASIP coat color loci may impact behavior in the horse. J. Hered. 2016, 107, 214–219. [Google Scholar] [CrossRef]
- Stachurska, A.; Pięta, M.; Łojek, J.; Szulowska, J. Performance in racehorses of various colours. Livest. Sci. 2007, 106, 282–286. [Google Scholar] [CrossRef]
- Bartolomé, E.; Goyache, F.; Molina, A.; Cervantes, I.; Valera, M.; Gutiérrez, J.P. Pedigree estimation of the subpopulation contribution to the total gene diversity: The horse coat colour case. Animal 2010, 4, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Druml, T.; Baumung, R.; Sölkner, J. Mophological analysis and effect of selection for conformation in the Noriker draught horse population. Livest. Sci. 2008, 115, 118–128. [Google Scholar] [CrossRef]
- Negro, S.; Imsland, F.; Valera, M.; Molina, A.; Solé, M.; Andersson, L. Association analysis of KIT, MUTF and PAX3 variants with White markings in Spanish horses. Anim. Genet. 2017, 48, 349–352. [Google Scholar] [CrossRef] [PubMed]
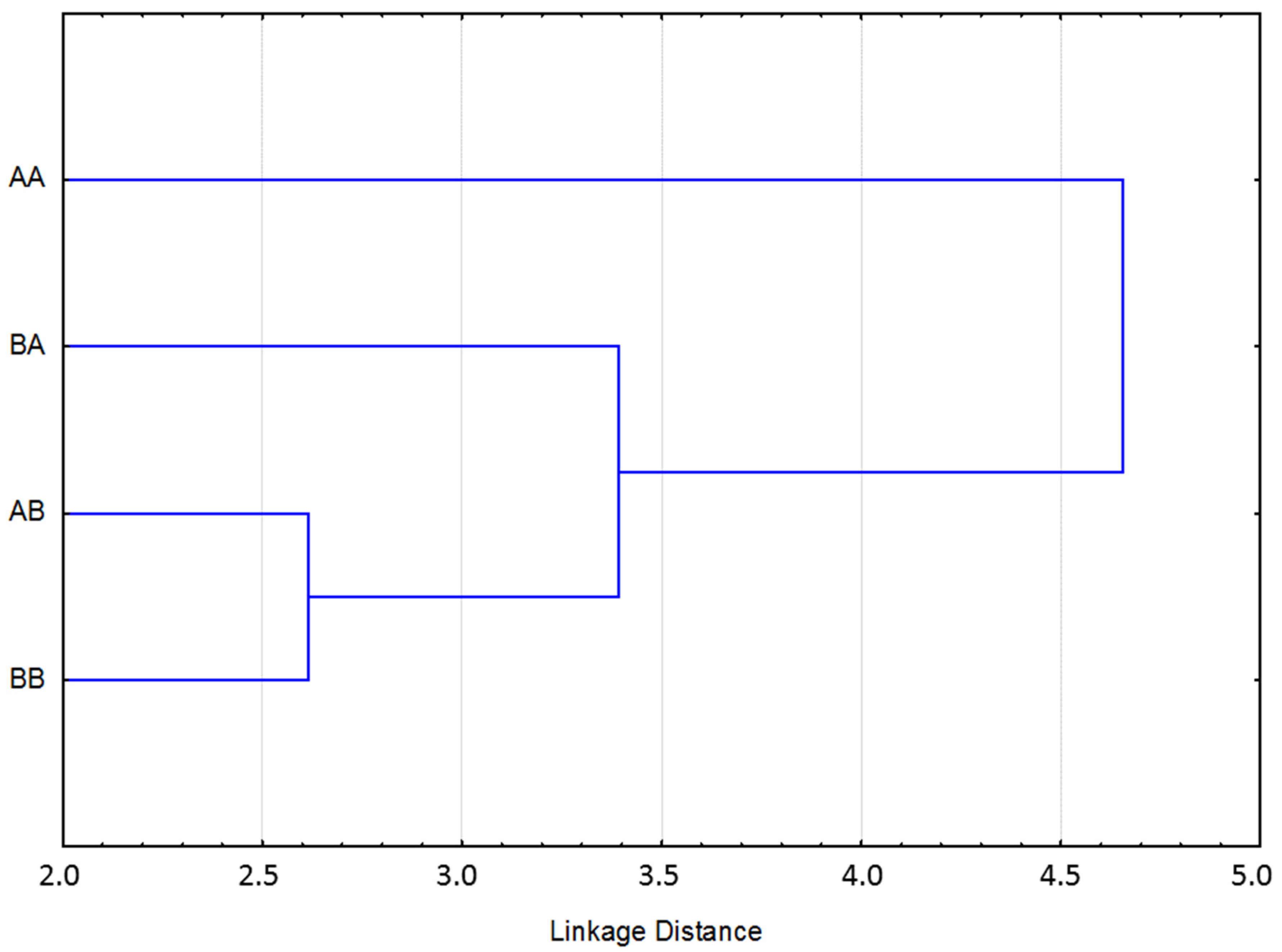

{kind=link}
| Trait | Class | Trait | Class | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 7 | 1 | 7 | ||||||
| Color | 1 | Quality of black coat | Pale | Very dark | Morphological traits | 24 | Chest width | Very narrow | Very wide |
| 2 | Percentage of white marks | Absence | Excess | 25 | Thorax width | Very narrow | Very wide | ||
| Morphological traits | 3 | Head width | Very narrow | Very wide | 26 | Forearm length | Very short | Very long | |
| 4 | Head length | Very short | Very long | 27 | Cannon bone length | Very short | Very long | ||
| 5 | Head depth | Shallow | Very deep | 28 | Fore knee perimeter | Very narrow | Very wide | ||
| 6 | Head profile | Concave | Convex | 29 | Cannon bone perimeter | Very narrow | Very wide | ||
| 7 | Head expression | Not much | Very much | 30 | Forelimb side view | Camped under | Camped out | ||
| 8 | Neck length | Very short | Very long | 31 | Fore hoof-pastern axis side view | Horizontal | Vertical | ||
| 9 | Head-neck junction | Very covered | Very marked | 32 | Fore-hoof slope | Horizontal | Vertical | ||
| 10 | Upper neck line | Concave | Convex | 33 | Form of hoof | Cylindrical | Bell-shaped | ||
| 11 | Lower neck line | Concave | Convex | 34 | Fore-hoof front view | Toe-in | Toe-out | ||
| 12 | Height of withers | Very short | Very high | 35 | Forelimb front view | Closed | Open | ||
| 13 | Shape of withers | Not prominent | Very prominent | 36 | Hock side view | Closed | Open | ||
| 14 | Shoulder length | Very short | Very long | 37 | Hock rear view | Closed | Open | ||
| 15 | Shoulder angle slope | Very horizontal | Very vertical | Functional traits | 38 | Walking activity | Very little | Hasty | |
| 16 | Thorax depth | Shallow | Very deep | 39 | Walking clarity | Very little | Much | ||
| 17 | Back length | Very short | Very long | 40 | Walking amplitude | Very little | Wide | ||
| 18 | Loin length | Very short | Very long | 41 | Walking suppleness | Very little | Much | ||
| 19 | Shape of back-loin line | Very concave | Slightly convex | 42 | Trotting amplitude | Very little | Wide | ||
| 20 | Croup length | Very short | Very long | 43 | Trotting suppleness | Very little | Much | ||
| 21 | Croup angle | Very horizontal | Very vertical | 44 | Trotting impulsion | Very little | Much | ||
| 22 | Buttock length | Very short | Very long | 45 | Trotting equilibrium | Downhill | Uphill | ||
| 23 | Withers-croup equilibrium | Downhill | Uphill | 46 | Trotting suspension | Very little | Much | ||
| Trait | Mean ± s.e. | CV (%) | Range | Trait | Mean ± s.e. | CV (%) | Range | ||
|---|---|---|---|---|---|---|---|---|---|
| CT | 1 | 4.80 ± 0.046 | 26.49 | 7 | MT | 24 | 5.01 ± 0.046 | 25.63 | 7 |
| 2 | 1.51 ± 0.036 | 64.95 | 7 | 25 | 4.83 ± 0.039 | 22.69 | 6 | ||
| MT | 3 | 5.23 ± 0.042 | 22.45 | 7 | 26 | 4.01 ± 0.027 | 18.78 | 6 | |
| 4 | 4.80 ± 0.038 | 21.72 | 6 | 27 | 3.55 ± 0.025 | 19.68 | 5 | ||
| 5 | 4.48 ± 0.033 | 20.30 | 6 | 28 | 3.67 ± 0.029 | 21.80 | 6 | ||
| 6 | 4.49 ± 0.032 | 19.53 | 7 | 29 | 3.21 ± 0.025 | 21.43 | 5 | ||
| 7 | 4.82 ± 0.032 | 18.68 | 6 | 30 | 3.50 ± 0.022 | 17.36 | 5 | ||
| 8 | 4.84 ± 0.037 | 21.27 | 7 | 31 | 4.16 ± 0.029 | 19.10 | 6 | ||
| 9 | 3.91 ± 0.040 | 28.11 | 7 | 32 | 3.98 ± 0.027 | 18.50 | 5 | ||
| 10 | 5.16 ± 0.028 | 15.31 | 6 | 33 | 3.42 ± 0.027 | 21.81 | 6 | ||
| 11 | 4.12 ± 0.030 | 20.52 | 6 | 34 | 4.30 ± 0.035 | 22.38 | 6 | ||
| 12 | 4.76 ± 0.043 | 25.32 | 6 | 35 | 4.03 ± 0.024 | 16.79 | 6 | ||
| 13 | 4.40 ± 0.052 | 32.89 | 7 | 36 | 4.40 ± 0.035 | 22.23 | 6 | ||
| 14 | 4.95 ± 0.034 | 19.10 | 6 | 37 | 4.00 ± 0.038 | 26.51 | 7 | ||
| 15 | 3.95 ± 0.042 | 29.18 | 7 | FT | 38 | 4.04 ± 0.031 | 21.42 | 7 | |
| 16 | 4.66 ± 0.037 | 21.81 | 6 | 39 | 4.71 ± 0.038 | 22.20 | 6 | ||
| 17 | 3.99 ± 0.029 | 19.87 | 5 | 40 | 4.35 ± 0.034 | 21.62 | 7 | ||
| 18 | 3.45 ± 0.030 | 23.77 | 6 | 41 | 4.38 ± 0.037 | 23.19 | 7 | ||
| 19 | 3.85 ± 0.030 | 21.36 | 7 | 42 | 4.59 ± 0.036 | 21.76 | 7 | ||
| 20 | 4.23 ± 0.031 | 20.53 | 7 | 43 | 4.33 ± 0.038 | 24.08 | 7 | ||
| 21 | 4.48 ± 0.042 | 25.77 | 7 | 44 | 4.70 ± 0.041 | 24.02 | 7 | ||
| 22 | 3.52 ± 0.037 | 29.52 | 7 | 45 | 4.50 ± 0.037 | 22.85 | 6 | ||
| 23 | 3.89 ± 0.039 | 28.06 | 7 | 46 | 4.52 ± 0.042 | 25.45 | 7 |
| Tr | σ2u (s.d.) | σ2pe (s.d.) | σ2e (s.d.) | h2 (s.d.) | Tr | σ2u (s.d.) | σ2pe (s.d.) | σ2e (s.d.) | h2 (s.d.) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CT | 1 | 0.47 (0.146) | 0.32 (0.115) | 0.51 (0.004) | 0.36 (0.098) | MT | 24 | 0.41 (0.137) | 0.36 (0.114) | 0.57 (0.039) | 0.30 (0.091) |
| 2 | 0.22 (0.146) | 0.64 (0.125) | 0.08 (0.001) | 0.23 (0.142) | 25 | 0.30 (0.090) | 0.19 (0.073) | 0.43 (0.030) | 0.32 (0.086) | ||
| MT | 3 | 0.36 (0.067) | 0.05 (0.039) | 0.53 (0.035) | 0.38 (0.057) | 26 | 0.15 (0.056) | 0.14 (0.047) | 0.27 (0.019) | 0.26 (0.088) | |
| 4 | 0.45 (0.105) | 0.12 (0.071) | 0.46 (0.031) | 0.44 (0.083) | 27 | 0.10 (0.035) | 0.10 (0.033) | 0.28 (0.019) | 0.21 (0.068) | ||
| 5 | 0.24 (0.055) | 0.07 (0.042) | 0.47 (0.032) | 0.30 (0.062) | 28 | 0.11 (0.042) | 0.09 (0.038) | 0.31 (0.021) | 0.21 (0.077) | ||
| 6 | 0.43 (0.076) | 0.07 (0.046) | 0.24 (0.017) | 0.58 (0.073) | 29 | 0.09 (0.034) | 0.10 (0.032) | 0.25 (0.018) | 0.19 (0.071) | ||
| 7 | 0.10 (0.061) | 0.16 (0.058) | 0.45 (0.031) | 0.14 (0.081) | 30 | 0.03 (0.025) | 0.16 (0.029) | 0.18 (0.012) | 0.09 (0.065) | ||
| 8 | 0.36 (0.105) | 0.13 (0.078) | 0.47 (0.032) | 0.37 (0.094) | 31 | 0.14 (0.064) | 0.17 (0.055) | 0.29 (0.020) | 0.23 (0.098) | ||
| 9 | 0.31 (0.105) | 0.19 (0.089) | 0.65 (0.045) | 0.27 (0.082) | 32 | 0.13 (0.046) | 0.10 (0.041) | 0.31 (0.021) | 0.24 (0.079) | ||
| 10 | 0.15 (0.057) | 0.21 (0.049) | 0.19 (0.013) | 0.28 (0.093) | 33 | 0.15 (0.066) | 0.16 (0.055) | 0.27 (0.019) | 0.26 (0.104) | ||
| 11 | 0.15 (0.080) | 0.14 (0.066) | 0.37 (0.026) | 0.23 (0.111) | 34 | 0.26 (0.116) | 0.46 (0.101) | 0.25 (0.017) | 0.26 (0.109) | ||
| 12 | 0.27 (0.132) | 0.51 (0.122) | 0.59 (0.040) | 0.19 (0.090) | 35 | 0.05 (0.042) | 0.22 (0.042) | 0.21 (0.014) | 0.11 (0.083) | ||
| 13 | 0.37 (0.151) | 0.58 (0.142) | 0.80 (0.055) | 0.21 (0.079) | 36 | 0.09 (0.072) | 0.49 (0.081) | 0.41 (0.028) | 0.10 (0.070) | ||
| 14 | 0.28 (0.077) | 0.11 (0.059) | 0.46 (0.031) | 0.33 (0.079) | 37 | 0.23 (0.113) | 0.51 (0.106) | 0.37 (0.026) | 0.20 (0.094) | ||
| 15 | 0.13 (0.082) | 0.21 (0.080) | 0.73 (0.050) | 0.12 (0.074) | FT | 38 | 0.06 (0.050) | 0.26 (0.054) | 0.35 (0.024) | 0.09 (0.070) | |
| 16 | 0.32 (0.065) | 0.06 (0.043) | 0.39 (0.026) | 0.41 (0.069) | 39 | 0.09 (0.065) | 0.35 (0.074) | 0.56 (0.039) | 0.09 (0.063) | ||
| 17 | 0.05 (0.036) | 0.13 (0.041) | 0.41 (0.028) | 0.09 (0.059) | 40 | 0.31 (0.119) | 0.23 (0.093) | 0.39 (0.027) | 0.33 (0.113) | ||
| 18 | 0.09 (0.034) | 0.04 (0.029) | 0.41 (0.027) | 0.16 (0.060) | 41 | 0.33 (0.118) | 0.19 (0.093) | 0.51 (0.035) | 0.32 (0.102) | ||
| 19 | 0.07 (0.038) | 0.18 (0.041) | 0.30 (0.021) | 0.12 (0.065) | 42 | 0.21 (0.089) | 0.34 (0.080) | 0.34 (0.024) | 0.23 (0.092) | ||
| 20 | 0.17 (0.067) | 0.13 (0.058) | 0.46 (0.032) | 0.23 (0.081) | 43 | 0.20 (0.087) | 0.27 (0.077) | 0.43 (0.030) | 0.22 (0.088) | ||
| 21 | 0.25 (0.099) | 0.35 (0.092) | 0.57 (0.039) | 0.21 (0.078) | 44 | 0.30 (0.129) | 0.37 (0.108) | 0.42 (0.029) | 0.28 (0.106) | ||
| 22 | 0.08 (0.054) | 0.19 (0.058) | 0.55 (0.038) | 0.10 (0.063) | 45 | 0.25 (0.096) | 0.23 (0.082) | 0.48 (0.033) | 0.26 (0.090) | ||
| 23 | 0.26 (0.119) | 0.43 (0.109) | 0.54 (0.038) | 0.21 (0.090) | 46 | 0.23 (0.129) | 0.48 (0.116) | 0.44 (0.031) | 0.20 (0.103) |
| Trait | rg1 (s.d.) | rg2 (s.d.) | Trait | rg1 (s.d.) | rg2 (s.d.) | ||
|---|---|---|---|---|---|---|---|
| MT | 3 | −0.114 (0.209) | −0.065 (0.410) | MT | 25 | 0.099 (0.243) | −0.155 (0.465) |
| 4 | 0.186 (0.206) | −0.443 (0.372) | 26 | 0.015 (0.268) | −0.177 (0.504) | ||
| 5 | −0.090 (0.224) | 0.295 (0.377) | 27 | 0.047 (0.275) | 0.084 (0.479) | ||
| 6 | 0.609 (0.184) | −0.204 (0.382) | 28 | −0.349 (0.430) | 0.617 (0.402) | ||
| 7 | 0.154 (0.355) | −0.030 (0.584) | 29 | −0.738 (0.225) | 0.448 (0.444) | ||
| 8 | 0.337 (0.234) | −0.404 (0.399) | 30 | −0.170 (0.270) | −0.175 (0.484) | ||
| 9 | −0.325 (0.270) | 0.074 (0.503) | 31 | 0.366 (0.255) | −0.269 (0.524) | ||
| 10 | 0.816 (0.171) | −0.434 (0.393) | 32 | 0.226 (0.272) | 0.069 (0.505) | ||
| 11 | −0.441 (0.302) | 0.227 (0.534) | 33 | 0.094 (0.445) | −0.807 (0.235) | ||
| 12 | −0.493 (0.323) | 0.469 (0435) | 34 | 0.311 (0.418) | −0.603 (0.414) | ||
| 13 | −0.767 (0.216) | 0.577 (0.372) | 35 | 0.025 (0.325) | 0.076 (0.560) | ||
| 14 | 0.187 (0.219) | 0.224 (0.430) | 36 | 0.310 (0.428) | 0.098 (0.598) | ||
| 15 | −0.419 (0.327) | 0.638 (0.365) | 37 | 0.185 (0.398) | −0.110 (0.574) | ||
| 16 | 0.269 (0.203) | 0.329 (0.349) | FT | 38 | 0.123 (0.244) | 0.396 (0.420) | |
| 17 | 0.618 (0.298) | 0.336 (0.529) | 39 | 0.318 (0.231) | −0.574 (0.316) | ||
| 18 | 0.292 (0.299) | −0.045 (0.573) | 40 | 0.325 (0.247) | −0.078 (0.492) | ||
| 19 | −0.224 (0.340) | −0.379 (0.484) | 41 | 0.243 (0.257) | −0.206 (0.475) | ||
| 20 | 0.307 (0.263) | −0.295 (0.479) | 42 | 0.376 (0.226) | −0.115 (0.502) | ||
| 21 | −0.459 (0.296) | 0.139 (0.483) | 43 | 0.224 (0.254) | −0.199 (0.482) | ||
| 22 | 0.229 (0.346) | −0.576 (0.405) | 44 | 0.241 (0.296) | −0.379 (0.498) | ||
| 23 | −0.371 (0.320) | 0.159 (0.518) | 45 | 0.099 (0.243) | −0.155 (0.465) | ||
| 24 | 0.411 (0.227) | 0.014 (0.446) | 46 | 0.015 (0.268) | −0.177 (0.504) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perdomo-González, D.I.; García de Paredes, R.d.l.A.; Valera, M.; Bartolomé, E.; Gómez, M.D. Morpho-Functional Traits in Pura Raza Menorquina Horses: Genetic Parameters and Relationship with Coat Color Variables. Animals 2022, 12, 2319. https://doi.org/10.3390/ani12182319
Perdomo-González DI, García de Paredes RdlA, Valera M, Bartolomé E, Gómez MD. Morpho-Functional Traits in Pura Raza Menorquina Horses: Genetic Parameters and Relationship with Coat Color Variables. Animals. 2022; 12(18):2319. https://doi.org/10.3390/ani12182319
Chicago/Turabian StylePerdomo-González, Davinia I., Rocío de las Aguas García de Paredes, Mercedes Valera, Ester Bartolomé, and María Dolores Gómez. 2022. "Morpho-Functional Traits in Pura Raza Menorquina Horses: Genetic Parameters and Relationship with Coat Color Variables" Animals 12, no. 18: 2319. https://doi.org/10.3390/ani12182319
APA StylePerdomo-González, D. I., García de Paredes, R. d. l. A., Valera, M., Bartolomé, E., & Gómez, M. D. (2022). Morpho-Functional Traits in Pura Raza Menorquina Horses: Genetic Parameters and Relationship with Coat Color Variables. Animals, 12(18), 2319. https://doi.org/10.3390/ani12182319