
Effect of Porcine Colostral Exosomes on T Cells in the Peripheral Blood of Suckling Piglets

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Isolation of Exosomes from Porcine Colostrum and Mature Milk
2.3. Isolation of Piglet PBMCs
2.4. Dynamin Inhibition in Piglet PBMCs
2.5. Culture of Piglet PBMCs with Milk-Derived Exosomes
2.6. Analysis of Cell Populations in Piglet PBMCs Using Flow Cytometry
2.7. Measurement of IFN-γ, IL-2 and IL-12 in Culture Medium
2.8. Statistical Analysis
3. Results
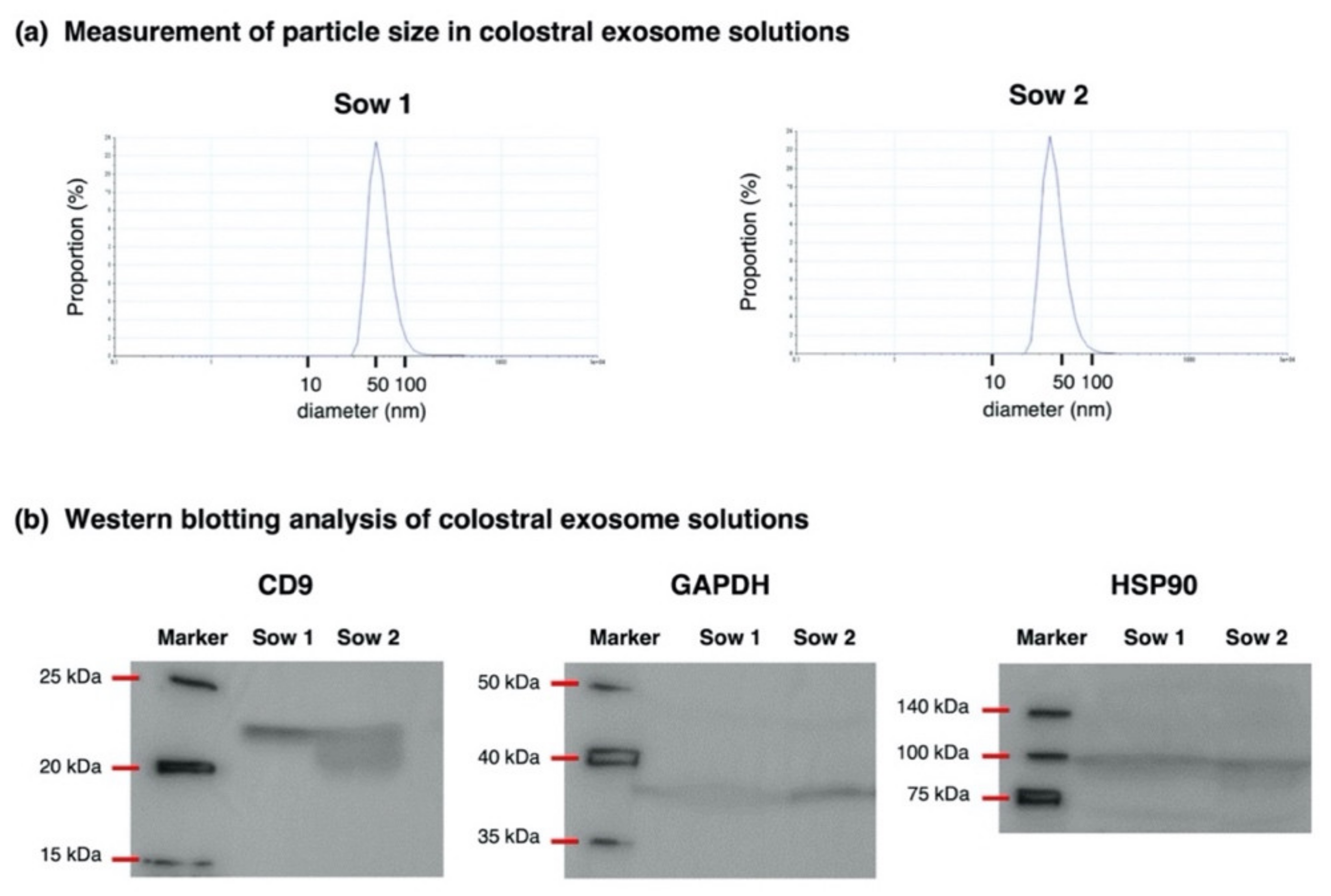
3.1. Validation of Exosome Isolation from Porcine Colostrum
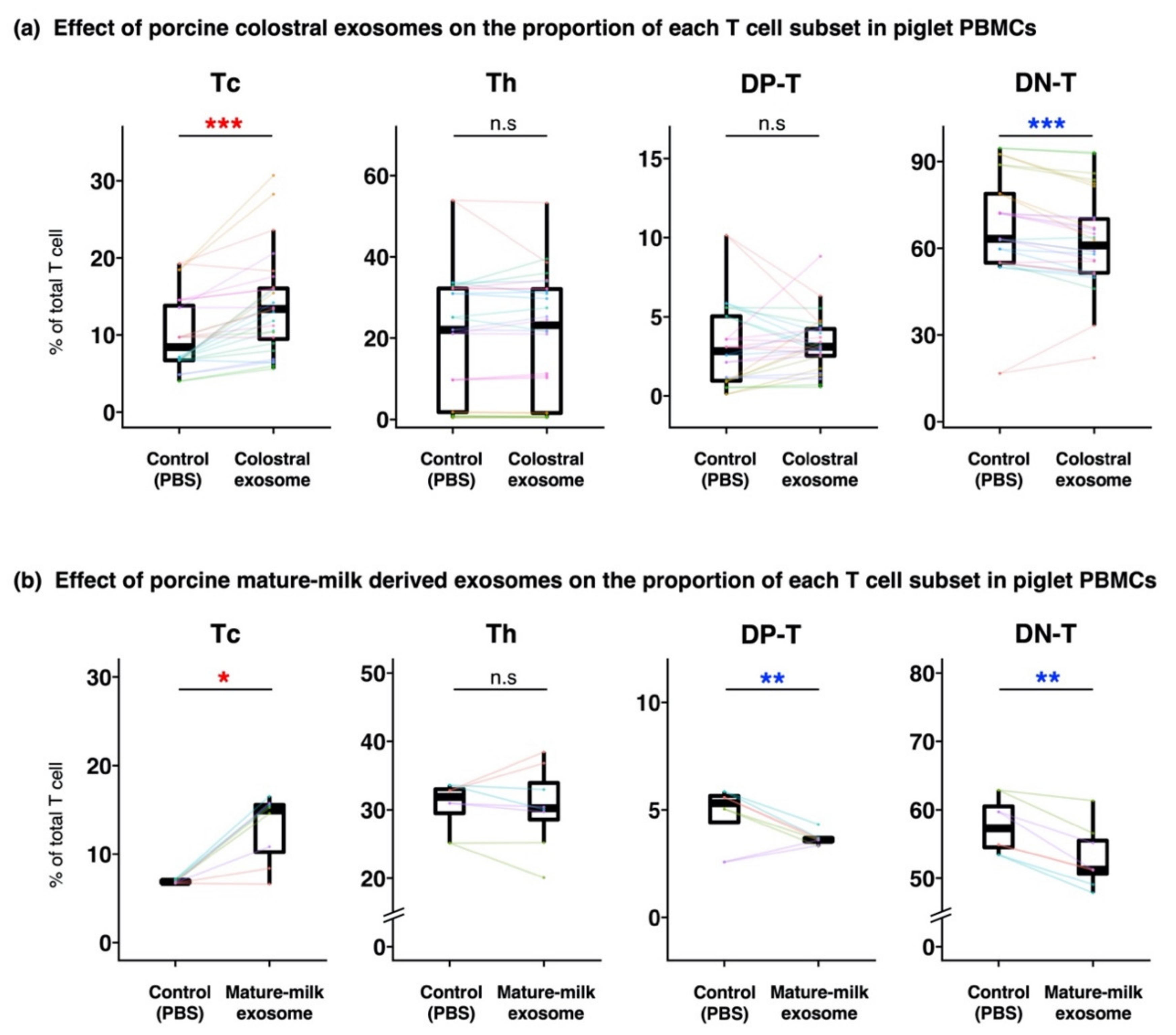
3.2. Compositions of T Cells in Piglet PBMCs Cultured with Milk-Derived Exosomes
3.3. Concentration of IFN-γ, IL-2 and IL-12 in Culture Medium
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Inoue, R.; Tsukahara, T. Composition and physiological functions of the porcine colostrum. Anim. Sci. J. 2021, 92, e13618. [Google Scholar] [CrossRef] [PubMed]
- Rooke, J.A.; Bland, I.M. The acquisition of passive immunity in the new-born piglet. Livest. Prod. Sci. 2002, 78, 13–23. [Google Scholar] [CrossRef]
- Ward, L.A.; Rich, E.D.; Besser, T.E. Role of maternally derived circulating antibodies in protection of neonatal swine against porcine group a Rotavirus. J. Infect. Dis. 1996, 174, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, S.; Tsukahara, T.; Imaoka, T.; Nakanishi, N.; Ushida, K.; Inoue, R. The effect of colostrum ingestion during the first 24 hours of life on early postnatal development of piglet immune systems. Anim. Sci. J. 2016, 87, 1511–1515. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Xie, M.-Y.; Sun, J.-J.; Ye, R.-S.; Cheng, X.; Sun, R.-P.; Wei, L.-M.; Li, M.; Lin, D.-L.; Jiang, Q.-Y.; et al. Porcine milk-derived exosomes promote proliferation of intestinal epithelial cells. Sci. Rep. 2016, 6, 33862. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Li, M.; Wang, T.; Liang, Y.; Zhong, Z.; Wang, X.; Zhou, Q.; Chen, L.; Lang, Q.; He, Z.; et al. Lactation-related microRNA expression profiles of porcine breast milk exosomes. PLoS ONE 2012, 7, e43691. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell. Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Alsaweed, M.; Hartmann, P.E.; Geddes, D.T.; Kakulas, F. MicroRNAs in breastmilk and the lactating breast: Potential immunoprotectors and developmental regulators for the infant and the mother. Int. J. Environ. Res. Public Health 2015, 12, 13981–14020. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Wang, H.; Luo, J.; Xie, M.; Zhao, Z.; Chen, X.; Wang, D.; Sun, J.; Xi, Q.; Chen, T.; et al. Porcine milk-derived small extracellular vesicles promote intestinal immunoglobulin production through pIgR. Animals 2021, 11, 1522. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zang, X.; Wang, M.; Li, Z.; Qiao, M.; Hu, H.; Chen, D. Exosome-based nanocarriers as bio-inspired and versatile vehicles for drug delivery: Recent advances and challenges. J. Mater. Chem. B 2019, 7, 2421–2433. [Google Scholar] [CrossRef]
- Sinkora, M.; Butler, J.E. The ontogeny of the porcine immune system. Dev. Comp. Immunol. 2009, 33, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Borghetti, P.; Angelis, E.D.; Saleri, R.; Cavalli, V.; Cacchioli, A.; Corradi, A.; Mocchegiani, E.; Martelli, P. Peripheral T lymphocyte changes in neonatal piglets: Relationship with growth hormone (GH), prolactin (PRL) and cortisol changes. Vet. Immunol. Immunop. 2006, 110, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Juul-Madsen, H.R.; Jensen, K.H.; Nielsen, J.; Damgaard, B.M. Ontogeny and characterization of blood leukocyte subsets and serum proteins in piglets before and after weaning. Vet. Immunol. Immunop. 2010, 133, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.J.; Sangild, P.T.; Zhang, Y.Q.; Zhang, S.H. Chapter 5 Bio-active compounds in porcine colostrum and milk and their effects on intestinal development in neonatal pigs. In Biology of Growing Animals; Zabielski, R., Gregory, P.C., Weström, B., Salek, E., Eds.; Elsevier: Amsterdam, The Netherlands, 2002; p. 10. [Google Scholar] [CrossRef]
- Ogawa, S.; Tsukahara, T.; Nishibayashi, R.; Nakatani, M.; Okutani, M.; Nakanishi, N.; Ushida, K.; Inoue, R. Shotgun proteomic analysis of porcine colostrum and mature milk. Anim. Sci. J. 2014, 85, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.F.; Blees, T.; Shakeri, F.; Buness, A.; Sylvester, M.; Savoini, G.; Agazzi, A.; Mrljak, V.; Sauerwein, H. Comparative proteome profiling in exosomes derived from porcine colostrum versus mature milk reveals distinct functional proteomes. J. Proteom. 2021, 249, 104338. [Google Scholar] [CrossRef] [PubMed]
- Kalia, V.; Sarkar, S. Regulation of effector and memory CD8 T cell differentiation by IL-2-A balancing act. Front. Immunol. 2018, 9, 2987. [Google Scholar] [CrossRef] [PubMed]
- Alspach, E.; Lussier, D.M.; Schreiber, R.D. Interferon γ and its important roles in promoting and inhibiting spontaneous and therapeutic cancer immunity. Csh. Perspect. Biol. 2019, 11, a028480. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, J.; Schmidt, C.; Mescher, M. The roles of IL-12 in providing a third signal for clonal expansion of naive CD8 T cells. J. Immunol. 2002, 169, 6842–6849. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Milk-Derived Exosomes | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sow ID | Sow 1 | Sow 2 | Sow 3 | Sow 4 | Sow 5 | Sow 6 | Sow 7 | |||||
| Source of PBMCs | Breed 1 | LW | LW | LW | LW | L | L | LW | ||||
| Piglet ID | Breed 1 | Location 2 | Age | Sex | Location 2 | Kyoto | Kyoto | Kanagawa | Kanagawa | Kanagawa | Kanagawa | Kyoto |
| Experiment 1 | ||||||||||||
| Piglet 1 | LWD | Kyoto | 21 d | Female | a | a | ||||||
| Piglet 2 | LWD | Kyoto | 20 d | Male | a | a | ||||||
| Piglet 3 | LWD | Kyoto | 20 d | Female | a | a | ||||||
| Piglet 4 | LWD | Kagawa | 29 d | Female | a | a | ||||||
| Piglet 5 | LWD | Kagawa | 29 d | Female | a | a | ||||||
| Experiment 2 | ||||||||||||
| Piglet 6 | LWD | Kanagawa | 30 d | Female | a, c | a, c | ||||||
| Piglet 7 | LWD | Kanagawa | 30 d | Female | a, b, c | a, b, c | ||||||
| Piglet 8 | LWD | Kanagawa | 25 d | Male | a, b, c | a, b, c | ||||||
| Piglet 9 | LWD | Kanagawa | 25 d | Female | a, b, c | a, b, c | ||||||
| Experiment 3 | ||||||||||||
| Piglet 10 | LD | Kanagawa | 30 d | Female | a | a | ||||||
| Piglet 11 | LD | Kanagawa | 27 d | Female | a | a | ||||||
| Piglet 12 | LD | Kanagawa | 15 d | Male | a, d | a, d | a | |||||
| Piglet 13 | LD | Kanagawa | 30 d | Male | a | a | a, d | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miura, H.; Jimbo, I.; Oda, M.; Noguchi, M.; Kawasaki, K.; Osada-Oka, M.; Tsukahara, T.; Inoue, R. Effect of Porcine Colostral Exosomes on T Cells in the Peripheral Blood of Suckling Piglets. Animals 2022, 12, 2172. https://doi.org/10.3390/ani12172172
Miura H, Jimbo I, Oda M, Noguchi M, Kawasaki K, Osada-Oka M, Tsukahara T, Inoue R. Effect of Porcine Colostral Exosomes on T Cells in the Peripheral Blood of Suckling Piglets. Animals. 2022; 12(17):2172. https://doi.org/10.3390/ani12172172
Chicago/Turabian StyleMiura, Hiroto, Itsuki Jimbo, Machi Oda, Michiko Noguchi, Kiyonori Kawasaki, Mayuko Osada-Oka, Takamitsu Tsukahara, and Ryo Inoue. 2022. "Effect of Porcine Colostral Exosomes on T Cells in the Peripheral Blood of Suckling Piglets" Animals 12, no. 17: 2172. https://doi.org/10.3390/ani12172172
APA StyleMiura, H., Jimbo, I., Oda, M., Noguchi, M., Kawasaki, K., Osada-Oka, M., Tsukahara, T., & Inoue, R. (2022). Effect of Porcine Colostral Exosomes on T Cells in the Peripheral Blood of Suckling Piglets. Animals, 12(17), 2172. https://doi.org/10.3390/ani12172172