
Effects of Decreasing Fishmeal as Main Source of Protein on Growth, Digestive Physiology, and Gut Microbiota of Olive Flounder (Paralichthys olivaceus)

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Experimental Fish and Sampling
2.3. Quantitative Analysis of Plasma GH and IGF-1
2.4. IGF-1 Gene Transcript Analysis
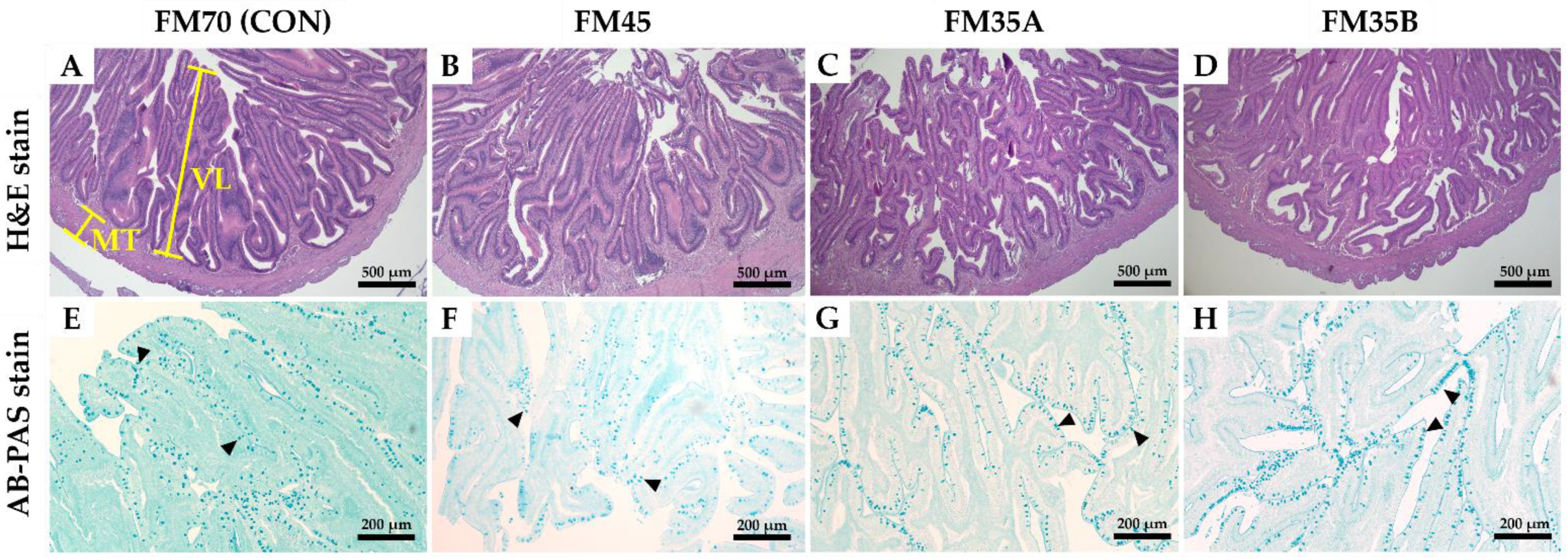
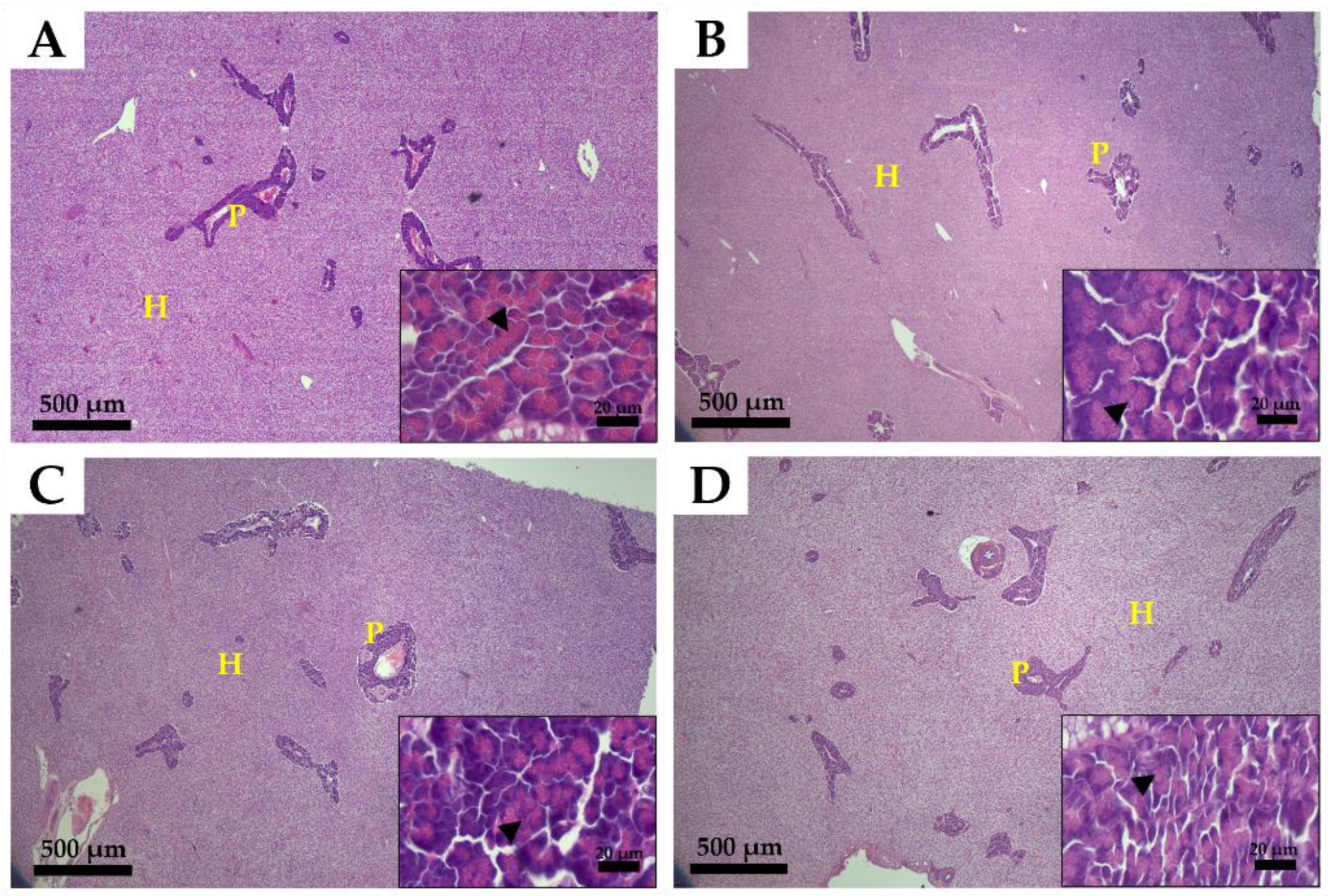
2.5. Histological Analysis
2.6. Digestive Enzyme Analysis
2.7. Gut Microbiota Analysis
2.8. Statistical Analysis
3. Results
3.1. Growth Performance
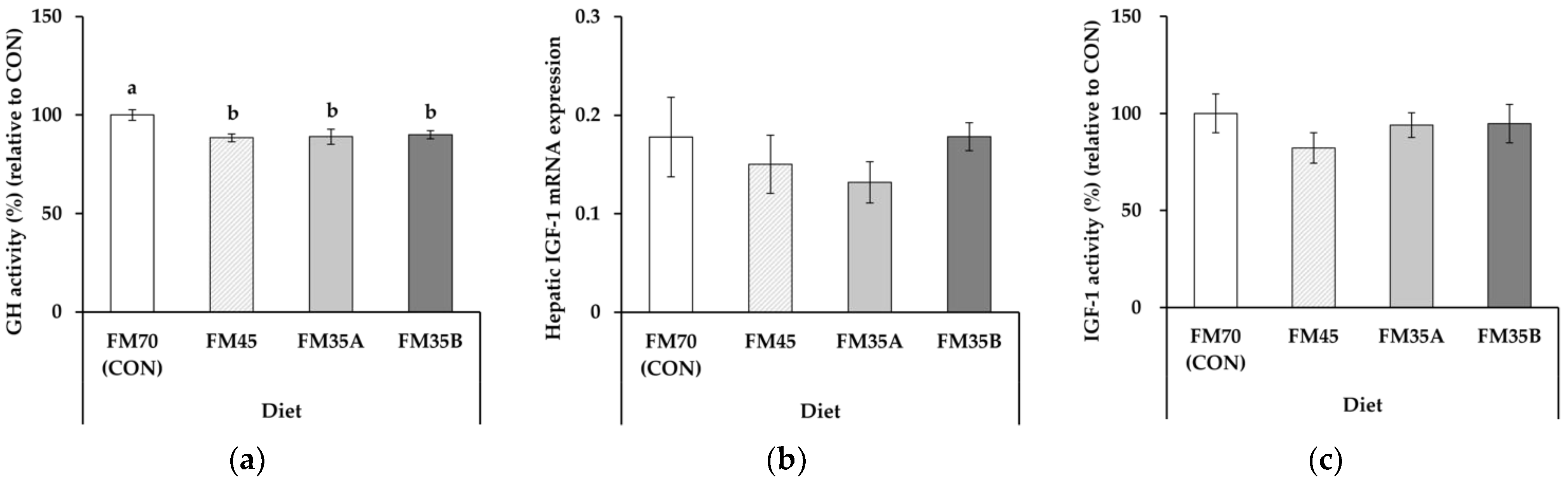
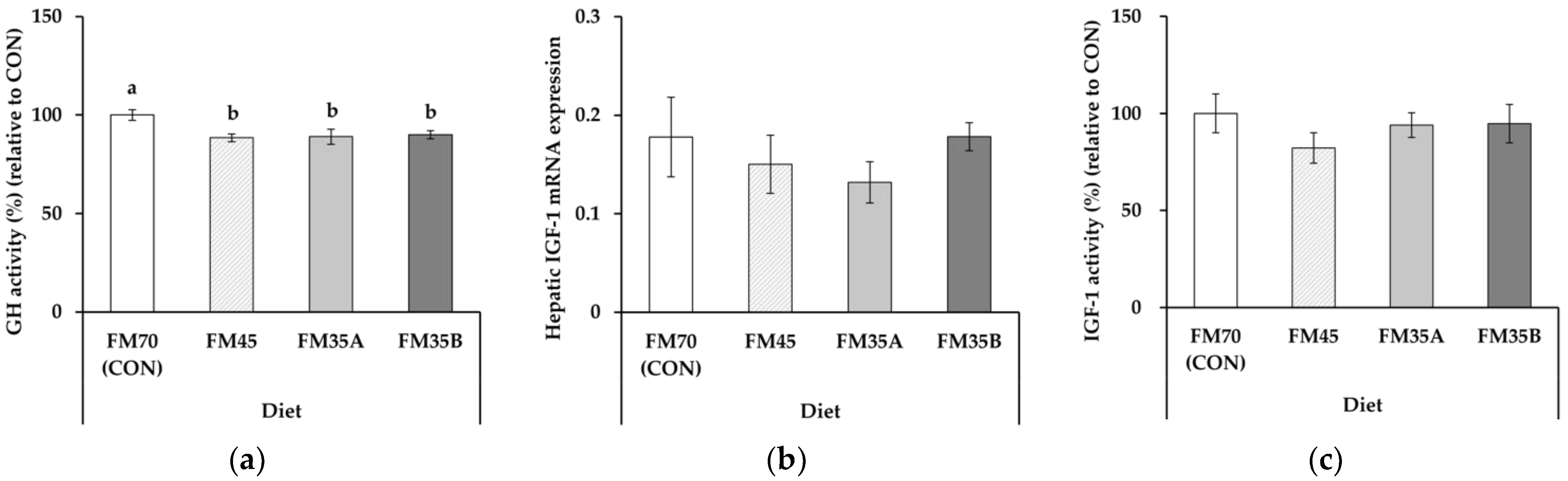
3.2. Growth-Related Factors
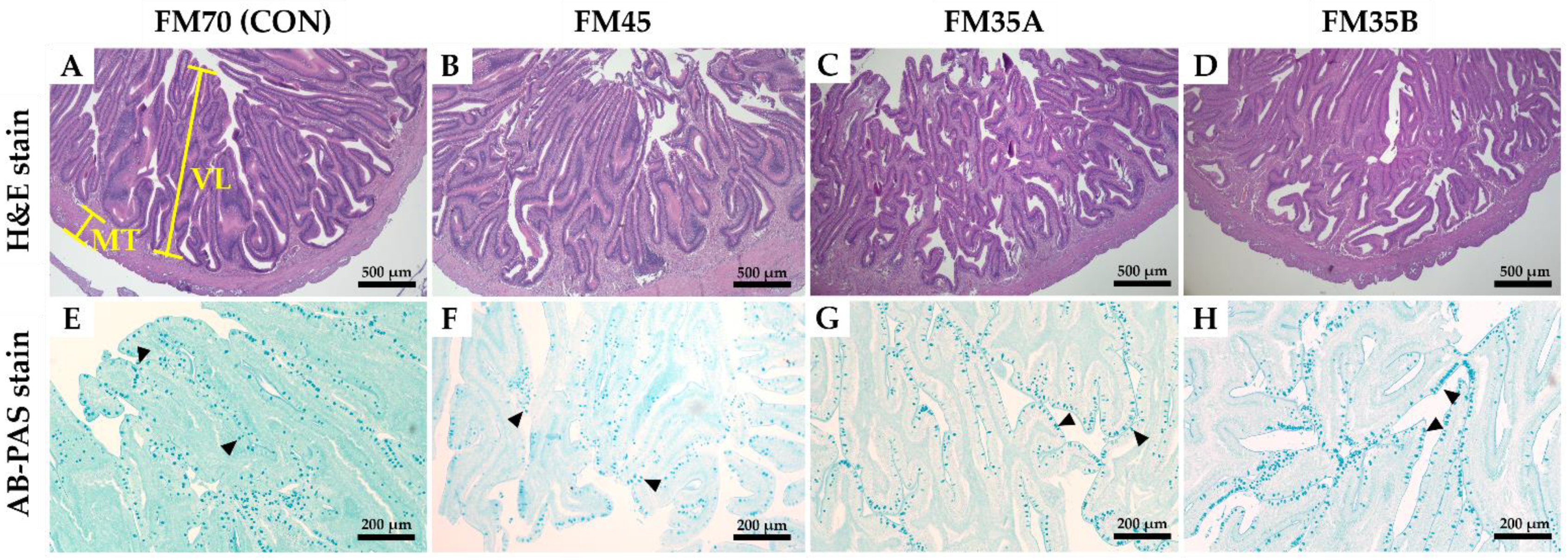
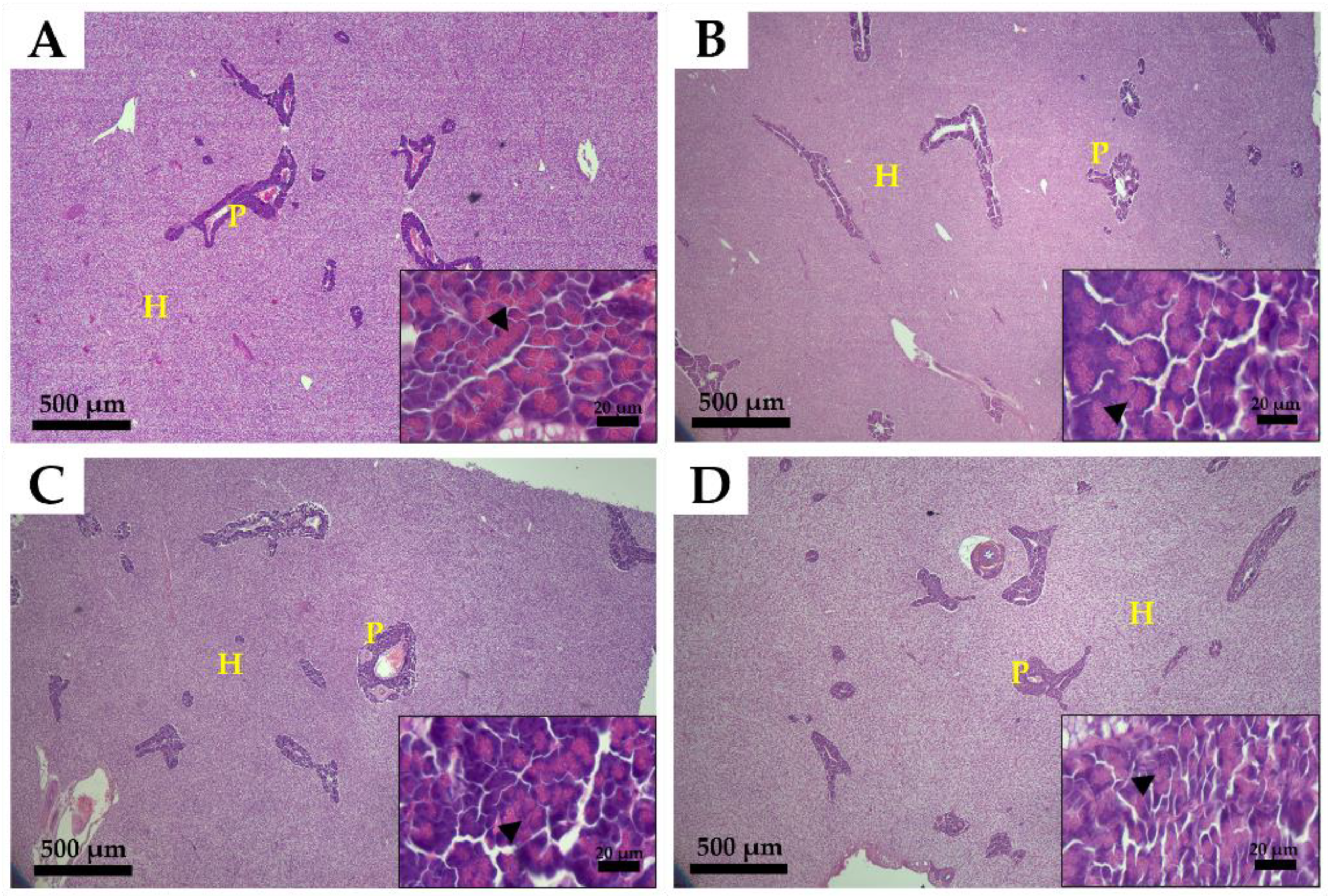
3.3. Histological Analysis of the Intestine and Liver
3.4. Digestive Enzyme Activity
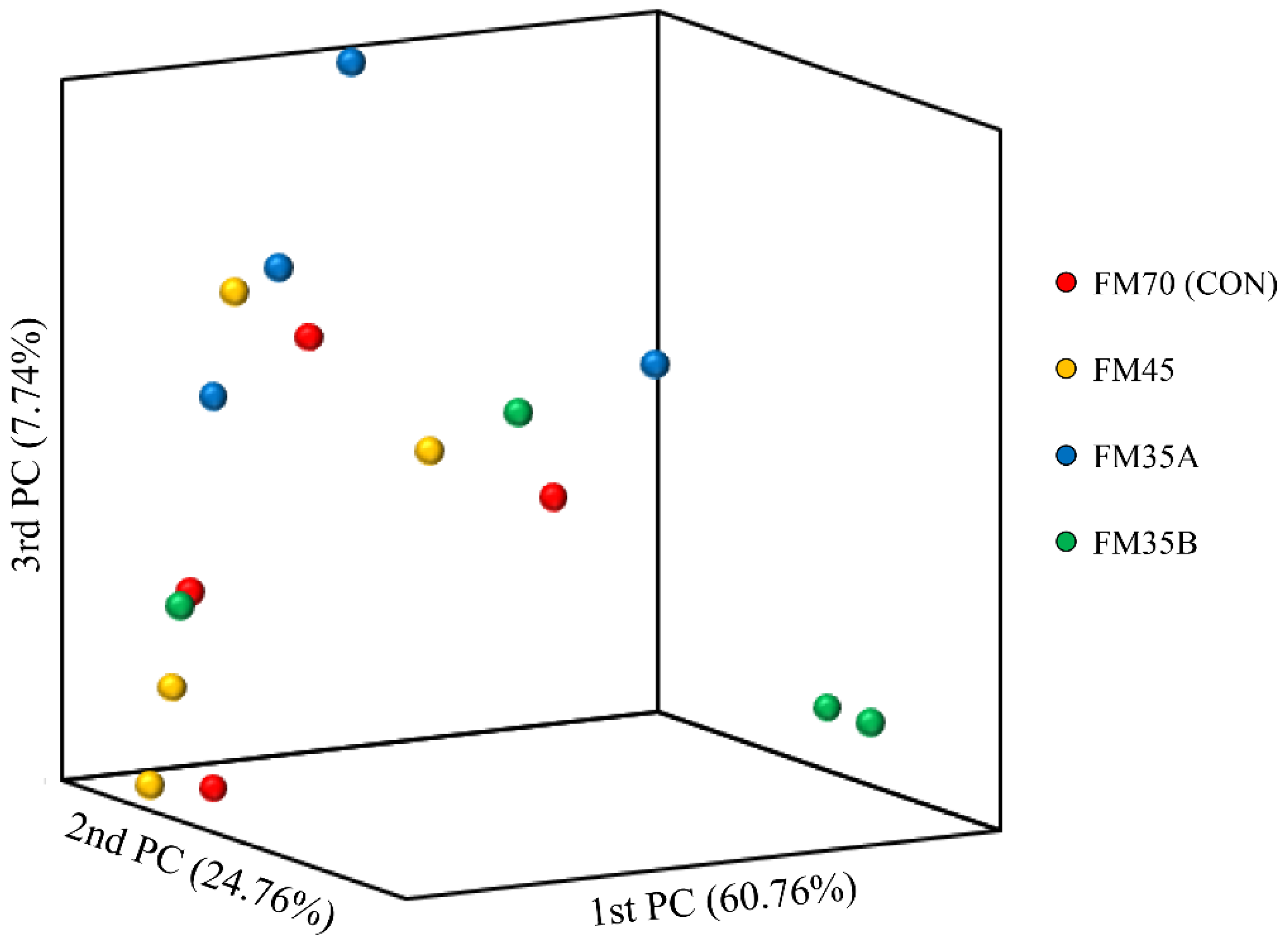
3.5. Diversity of Gut Microbiota
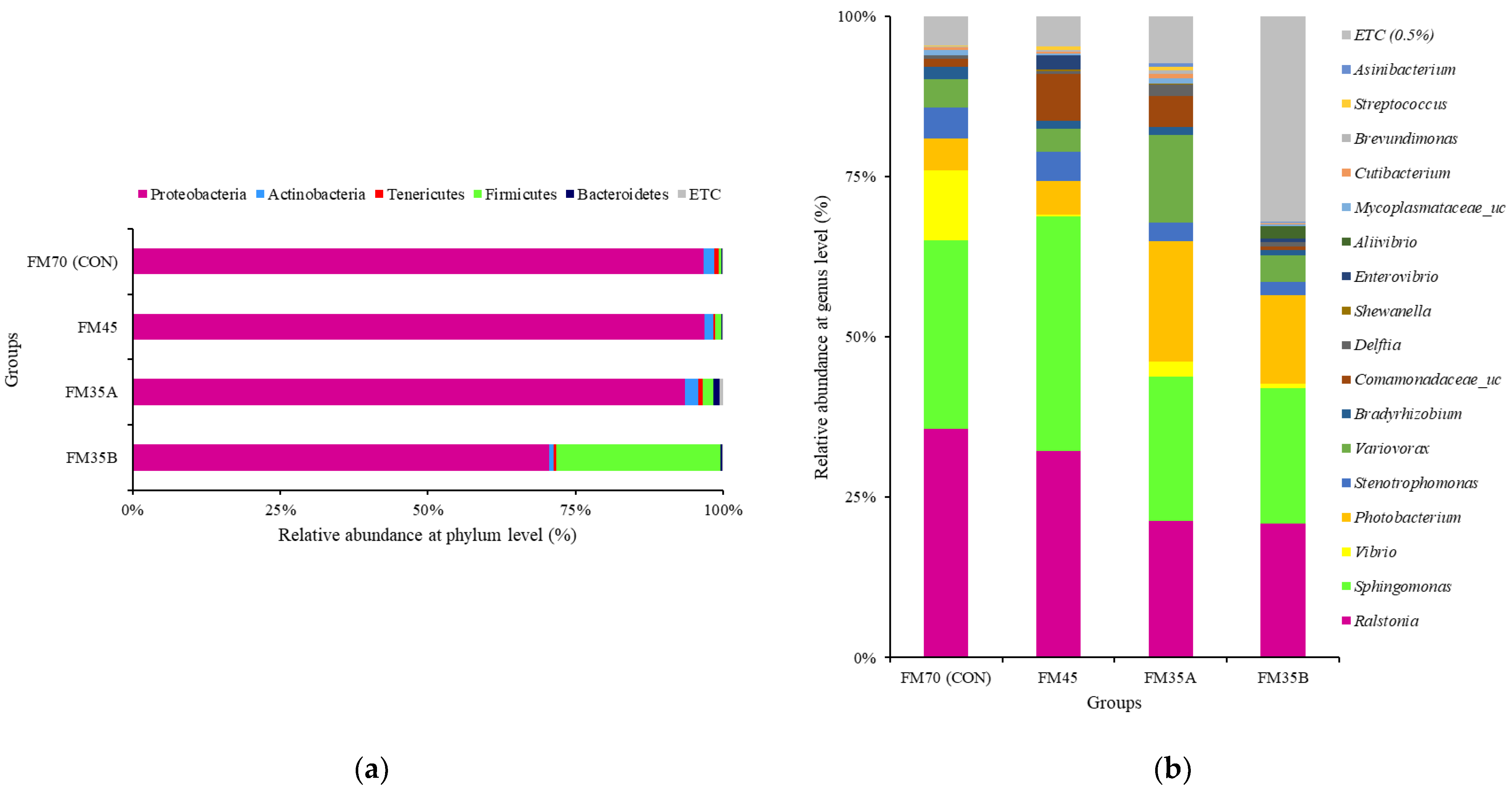
3.6. Gut Microbiota Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- KOSIS. Korean Statistical Information Service. 2022. Available online: https://Kosis.kr/eng/ (accessed on 24 March 2022).
- Hamidoghli, A.; Won, S.; Lee, S.; Lee, S.; Farris, N.W.; Bai, S.C. Nutrition and feeding of olive flounder Paralichthys olivaceus: A Review. Rev. Fish. Sci. Aquac. 2020, 28, 340–357. [Google Scholar] [CrossRef]
- Tacon, A.G.; Metian, M. Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects. Aquaculture 2008, 285, 146–158. [Google Scholar] [CrossRef]
- Shepherd, C.J.; Jackson, A.J. Global fishmesal and fish-oil supply: Inputs, outputs and marketsa. J. Fish Biol. 2013, 83, 1046–1066. [Google Scholar] [CrossRef]
- Choi, S.M.; Wang, X.; Park, G.J.; Lim, S.R.; Kim, K.W.; Bai, S.C.; Shin, I.S. Dietary dehulled soybean meal as a replacement for fish meal in fingerling and growing olive flounder Paralichthys olivaceus (Temminck et Schlegel). Aquac. Res. 2004, 35, 410–418. [Google Scholar] [CrossRef]
- Wei, Z.; Kangsen, M.; Baigang, Z.; Yangjiang, H.; Yu, Y. A study on the meat and bone meal or poultry by-product meal as protein substitutes of fishmeal in concentrated diets for Paralichthys olivaceus. J. Ocean Univ. China 2006, 5, 63–66. [Google Scholar] [CrossRef]
- Ye, J.; Liu, X.; Wang, Z.; Wang, K. Effect of partial fish meal replacement by soybean meal on the growth performance and biochemical indices of juvenile Japanese flounder Paralichthys olivaceus. Aquac. Int. 2011, 19, 143–153. [Google Scholar] [CrossRef]
- Kim, S.S.; Oh, D.H.; Cho, S.J.; Seo, S.H.; Han, H.S.; Lee, K.J. Evaluation of acid-concentrated soybean meal as a fishmeal replacement and its digestibility in diets for juvenile olive flounder Paralichthys olivaceus. Korean J. Fish. Aquat. Sci. 2014, 47, 824–831. [Google Scholar] [CrossRef]
- Jeong, S.M.; Khosravi, S.; Mauliasari, I.R.; Lee, B.J.; You, S.G.; Lee, S.M. Nutritional evaluation of cricket, Gryllus bimaculatus, meal as fish meal substitute for olive flounder, Paralichthys olivaceus, juveniles. J. World Aquac. Soc. 2021, 52, 859–880. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Khalil, R.H.; Metwally, A.A.; Shakweer, M.S.; Khallaf, M.A.; Abdel-Latif, H.M. Effects of black soldier fly (Hermetia illucens L.) larvae meal on growth performance, organs-somatic indices, body composition, and hemato-biochemical variables of European sea bass, Dicentrarchus labrax. Aquaculture 2020, 522, 735136. [Google Scholar] [CrossRef]
- Green, T.J.; Smullen, R.; Barnes, A.C. Dietary soybean protein concentrate-induced intestinal disorder in marine farmed Atlantic salmon, Salmo salar is associated with alterations in gut microbiota. Vet. Microbiol. 2013, 166, 286–292. [Google Scholar] [CrossRef]
- Luthada-Raswiswi, R.W.; O’Brien, G.; Mukaratirwa, S. Fishmeal replacement with animal protein source (Crocodylus niloticus Meat Meal) in diets of mozambique tilapia (Oreochromis mossambicus) of Different Size Groups. Appl. Sci. 2022, 12, 7211. [Google Scholar] [CrossRef]
- Parma, L.; Candela, M.; Soverini, M.; Turroni, S.; Consolandi, C.; Brigidi, P.; Mandrioli, L.; Sirri, R.; Fontanillas, R.; Gatta, P.P.; et al. Next-generation sequencing characterization of the gut bacterial community of gilthead sea bream (Sparus aurata, L.) fed low fishmeal based diets with increasing soybean meal levels. Anim. Feed Sci. Technol. 2016, 222, 204–216. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, C.; Shin, J.; Lee, K.J. Effect of Dietary Supplementation with Citrus By-product as a Vitamin C Replacement for Juvenile Olive Flounder Paralichthys olivaceus at Low Water Temperatures. Korean J. Fish. Aquat. Sci. 2017, 50, 15–24. [Google Scholar] [CrossRef]
- Hasan, M.T.; Jang, W.J.; Lee, B.J.; Kim, K.W.; Hur, S.W.; Lim, S.G.; Bai, S.C.; Kong, I.S. Heat-killed Bacillus sp. SJ-10 probiotic acts as a growth and humoral innate immunity response enhancer in olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2019, 88, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Dong, X.; Tan, B.; Du, T.; Zhang, S.; Yang, Y.; Chi, S.; Yang, Q.; Liu, H. Effects of fish meal replacement by low-gossypol cottonseed meal on growth performance, digestive enzyme activity, intestine histology and inflammatory gene expression of silver sillago (Sillago sihama Forsskál) (1775). Aquac. Nutr. 2020, 26, 1724–1735. [Google Scholar] [CrossRef]
- Cetinkaya, C.; Sisman, A.R.; Kiray, M.; Camsari, U.M.; Gencoglu, C.; Baykara, B.; Aksu, I.; Uysal, N. Positive effects of aerobic exercise on learning and memory functioning, which correlate with hippocampal IGF-1 increase in adolescent rats. Neurosci. Lett. 2013, 549, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, E.N.; Björnsson, B.T.; Valdés, J.A.; Einarsdottir, I.E.; Lorca, B.; Alvarez, M.; Molina, A. IGF-I/PI3K/Akt and IGF-I/MAPK/ERK pathways in vivo in skeletal muscle are regulated by nutrition and contribute to somatic growth in the fine flounder. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2011, 300, R1532–R1542. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; He, Y.; Chi, S.; Tan, B.; Lin, S.; Dong, X.; Yang, Q.; Liu, H.; Zhang, S. Supplementation with Saccharomyces cerevisiae hydrolysate in a complex plant protein, low-fishmeal diet improves intestinal morphology, immune function and Vibrio harveyi disease resistance in Epinephelus coioides. Aquaculture 2020, 529, 735655. [Google Scholar] [CrossRef]
- Martínez-Lagos, R.; Tovar-Ramírez, D.; Gracia-López, V.; Lazo, J.P. Changes in digestive enzyme activities during larval development of leopard grouper (Mycteroperca rosacea). Fish Physiol. Biochem. 2014, 40, 773–785. [Google Scholar] [CrossRef]
- Gupta, S.K.; Fotedar, R.; Foysal, M.; Priyam, M.; Siddik, M.A.; Chaklader, M.; Dao, T.T.T.; Howieson, J. Impact of varied combinatorial mixture of non-fishmeal ingredients on growth, metabolism, immunity and gut microbiota of Lates calcarifer (Bloch, 1790) fry. Sci. Rep. 2020, 10, 17091. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Moon, J.S.; Seo, J.S.; Nam, T.J.; Lee, K.J.; Lim, S.G.; Kim, K.W.; Lee, B.J.; Hur, S.W.; Choi, Y.H. 2019. Effect of fish meal replacement on insulin-like growth factor-I expression in the liver and muscle and implications for the growth of olive flounder Paralichthys olivaceus. Korean J. Fish. Aquat. Sci. 2019, 52, 141–148. [Google Scholar] [CrossRef]
- Jo, S.-J.; Park, S.-J.; Lee, S.-B.; Tran, B.T.; Kim, J.S.; Song, J.-W.; Lee, B.-J.; Hur, S.-W.; Nam, T.-J.; Lee, K.-J.; et al. Effect of low-fishmeal diets on some digestive physiological responses of juvenile and growing olive flounder (Paralichthys olivaceus) fed at an industrial-scale fish farm. Aquac. Rep. 2021, 21, 100904. [Google Scholar] [CrossRef]
- Niu, K.M.; Lee, B.J.; Kothari, D.; Lee, W.D.; Hur, S.W.; Lim, S.G.; Kim, K.W.; Kim, K.D.; Kim, N.N.; Kim, S.K. Dietary effect of low fish meal aquafeed on gut microbiota in olive flounder (Paralichthys olivaceus) at different growth stages. MicrobiologyOpen 2020, 9, e992. [Google Scholar] [CrossRef]
- Park, S.J.; Seo, B.S.; Park, H.S.; Lee, B.J.; Hur, S.W.; Nam, T.J.; Lee, K.J.; Lee, S.; Choi, Y.H. Effect of fishmeal content in the diet on the growth and sexual maturation of olive flounder (Paralichthys olivaceus) at a typical fish farm. Animals 2021, 11, 2055. [Google Scholar] [CrossRef] [PubMed]
- Hack, N.L.; Strobel, J.S.; Journey, M.L.; Beckman, B.R.; Lema, S.C. Response of the insulin-like growth factor-1 (Igf1) system to nutritional status and growth rate variation in olive rockfish (Sebastes serranoides). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2018, 224, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Statist. 1984, 11, 265–270. Available online: https://www.jstor.org/stable/4615964 (accessed on 20 May 2022).
- Magurran, A.E. Measuring biological diversity. Curr. Biol. 2021, 31, R1174–R1177. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef]
- Kim, M.G.; Lim, H.; Lee, B.J.; Hur, S.W.; Lee, S.; Kim, K.W.; Lee, K.J. Replacing fish meal with a mixture of plant and animal protein sources in the diets of juvenile olive flounder Paralichthys olivaceus. Korean J. Fish. Aquat. Sci. 2020, 53, 577–582. [Google Scholar] [CrossRef]
- Kim, M.G.; Shin, J.; Lee, C.; Lee, B.J.; Hur, S.W.; Lim, S.G.; Lee, K.J. Evaluation of a mixture of plant protein source as a partial fish meal replacement in diets for juvenile olive flounder Paralichthys olivaceus. Korean J. Fish. Aquat. Sci. 2019, 52, 374–381. [Google Scholar] [CrossRef]
- Yuan, X.Y.; Jiang, G.Z.; Cheng, H.H.; Cao, X.F.; Wang, C.C.; Dai, Y.J.; Liu, W.B. Replacing fish meal with cottonseed meal protein hydrolysate affects growth, intestinal function, and growth hormone/insulin-like growth factor I axis of juvenile blunt snout bream (Megalobrama amblycephala). J. World Aquac. Soc. 2020, 51, 1235–1249. [Google Scholar] [CrossRef]
- Picha, M.E.; Turano, M.J.; Beckman, B.R.; Borski, R.J. Endocrine biomarkers of growth and applications to aquaculture: A minireview of growth hormone, insulin-like growth factor (IGF)-I, and IGF-binding proteins as potential growth indicators in fish. N. Am. J. Aquacult. 2008, 70, 196–211. [Google Scholar] [CrossRef]
- Gómez-Requeni, P.; Mingarro, M.; Calduch-Giner, J.A.; Médale, F.; Martin, S.A.M.; Houlihan, D.F.; Kaushik, S.; Pérez-Sánchez, J. Protein growth performance, amino acid utilisation and somatotropic axis responsiveness to fish meal replacement by plant protein sources in gilthead sea bream (Sparus aurata). Aquaculture 2004, 232, 493–510. [Google Scholar] [CrossRef]
- Dyer, A.R.; Barlow, C.G.; Bransden, M.P.; Carter, C.G.; Glencross, B.D.; Richardson, N.; Thomas, P.M.; Carragher, J.F. Correlation of plasma IGF-I concentrations and growth rate in aquacultured finfish: A tool for assessing the potential of new diets. Aquaculture 2004, 236, 583–592. [Google Scholar] [CrossRef]
- Pérez-Sánchez, J.; Martí-Palanca, H.; Kaushik, S.J. Ration size and protein intake affect circulating growth hormone concentration, hepatic growth hormone binding and plasma insulin-like growth factor-I immunoreactivity in a marine teleost, the gilthead sea bream (Sparus aurata). J. Nutr. 1995, 125, 546–552. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, H.; Zhang, J.; Lin, B.; Chen, L.; Wang, Q.; Li, G.; Deng, J. 2020. Assessment of rapeseed meal as fish meal alternative in diets for juvenile Asian red-tailed catfish (Hemibagrus wyckioides). Aquac. Rep. 2019, 18, 100497. [Google Scholar] [CrossRef]
- Kumar, V.; Khalil, W.K.B.; Weiler, U.; Becker, K. Influences of incorporating detoxified Jatropha curcas kernel meal in common carp (Cyprinus carpio L.) diet on the expression of growth hormone-and insulin-like growth factor-1-encoding genes. J. Anim. Physiol. Anim. Nutr. 2013, 97, 97–108. [Google Scholar] [CrossRef]
- Kim, S.; Kang, J.C.; Lee, J.S. 2016. Morphology and Histology of the Digestive Organ in the Sablefish, Anoplopoma fimbria (Teleostei: Anoplopomatidae). Korean J. Ichthyol. 2016, 28, 19–27. [Google Scholar]
- Rapatsa, M.M.; Moyo, N.A. Evaluation of Imbrasia belina meal as a fishmeal substitute in Oreochromis mossambicus diets: Growth performance, histological analysis and enzyme activity. Aquac. Rep. 2017, 5, 18–26. [Google Scholar] [CrossRef]
- Yang, P.; Li, X.; Song, B.; He, M.; Wu, C.; Leng, X. The potential of Clostridium autoethanogenum, a new single cell protein, in substituting fish meal in the diet of largemouth bass (Micropterus salmoides): Growth, feed utilization and intestinal histology. Aquac. Fish. 2021, 26, 1–9. [Google Scholar] [CrossRef]
- Zhou, Z.; Yao, W.; Ye, B.; Wu, X.; Li, X.; Dong, Y. Effects of replacing fishmeal protein with poultry by-product meal protein and soybean meal protein on growth, feed intake, feed utilization, gut and liver histology of hybrid grouper (Epinephelus fuscoguttatus♀ × Epinephelus lanceolatus♂) juveniles. Aquaculture 2020, 516, 734503. [Google Scholar] [CrossRef]
- Leduc, A.; Zatylny-Gaudin, C.; Robert, M.; Corre, E.; Le Corguille, G.; Castel, H.; Lefevre-Scelles, A.; Fournier, V.; Gisbert, E.; Andree, K.B.; et al. Dietary aquaculture by-product hydrolysates: Impact on the transcriptomic response of the intestinal mucosa of European seabass (Dicentrarchus labrax) fed low fish meal diets. BMC Genom. 2018, 19, 396. [Google Scholar] [CrossRef] [PubMed]
- Gunathilaka, B.E.; Khosravi, S.; Herault, M.; Fournier, V.; Lee, C.; Jeong, J.B.; Lee, K.J. Evaluation of shrimp or tilapia protein hydrolysate at graded dosages in low fish meal diet for olive flounder (Paralichthys olivaceus). Aquac. Nutr. 2020, 26, 1592–1603. [Google Scholar] [CrossRef]
- Moutinho, S.; Peres, H.; Serra, C.; Martínez-Llorens, S.; Tomás-Vidal, A.; Jover-Cerdá, M.; Oliva-Teles, A. Meat and bone meal as partial replacement of fishmeal in diets for gilthead sea bream (Sparus aurata) juveniles: Diets digestibility, digestive function, and microbiota modulation. Aquaculture 2017, 479, 721–731. [Google Scholar] [CrossRef]
- Zhou, L.; Budge, S.M.; Ghaly, A.E.; Brooks, M.S.; Dave, D. Extraction, purification and characterization of fish chymotrypsin: A review. Am. J. Biochem. Biotechnol. 2011, 7, 104–123. [Google Scholar] [CrossRef]
- Ni, J.; Yan, Q.; Yu, Y.; Zhang, T. Factors influencing the grass carp gut microbiome and its effect on metabolism. FEMS Microbiol. Ecol. 2014, 87, 704–714. [Google Scholar] [CrossRef]
- Jang, W.J.; Lee, S.J.; Jeon, M.H.; Kim, T.Y.; Lee, J.M.; Hasan, M.T.; Lee, H.T.; Park, J.H.; Lee, B.J.; Hur, S.W.; et al. Characterization of a Bacillus sp. KRF-7 isolated from the intestine of rockfish and effects of dietary supplementation with mannan oligosaccharide in rockfish aquaculture. Fish Shellfish Immunol. 2021, 119, 182–192. [Google Scholar] [CrossRef]
- Gao, Q.; Luo, J.; Liu, P.; Yi, S.; Tang, Q.; Li, J.; Xia, Z.; Yang, G. Characteristics of intestinal microbiota in male morphotypes of the giant freshwater prawn Macrobrachium rosenbergii. Aquaculture 2022, 555, 738200. [Google Scholar] [CrossRef]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Wastyk, H.C.; Fragiadakis, G.K.; Perelman, D.; Dahan, D.; Merrill, B.D.; Feiqiao, B.Y.; Topf, M.; Gonzalez, C.G.; Treuren, W.V.; Han, S.; et al. Gut-microbiota-targeted diets modulate human immune status. Cell 2021, 184, 4137–4153. [Google Scholar] [CrossRef] [PubMed]
- Boland, K.; Bedrani, L.; Turpin, W.; Kabakchiev, B.; Stempak, J.; Borowski, K.; Nguyen, G.; Steinhart, A.H.; Smith, M.I.; Croitoru, K.; et al. Persistent diarrhea in patients with Crohn’s disease after mucosal healing is associated with lower diversity of the intestinal microbiome and increased dysbiosis. Clin. Mol. Hepatol. 2021, 19, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.J.; Hasan, M.T.; Lee, B.J.; Hur, S.W.; Lee, S.; Kim, K.W.; Lee, E.W.; Kong, I.S. Effect of dietary differences on changes of intestinal microbiota and immune-related gene expression in juvenile olive flounder (Paralichthys olivaceus). Aquaculture 2020, 527, 735442. [Google Scholar] [CrossRef]
- Jang, W.J.; Hasan, M.T.; Lee, G.H.; Lee, B.J.; Hur, S.W.; Lee, S.; Kim, K.W.; Lee, E.W.; Kong, I.S. Comparison of spore or vegetative Bacillus sp. supplementation on physiological changes and gut microbiota of the olive flounder (Paralichthys olivaceus). Aquaculture 2021, 535, 736355. [Google Scholar] [CrossRef]
- Niu, K.M.; Khosravi, S.; Kothari, D.; Lee, W.D.; Lim, J.M.; Lee, B.J.; Kim, K.W.; Lim, S.G.; Lee, S.M.; Kim, S.K. Effects of dietary multi-strain probiotics supplementation in a low fishmeal diet on growth performance, nutrient utilization, proximate composition, immune parameters, and gut microbiota of juvenile olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2019, 93, 258–268. [Google Scholar] [CrossRef]
- Sohn, H.; Kim, J.; Jin, C.; Lee, J. Identification of Vibrio species isolated from cultured olive flounder (Paralichthys olivaceus) in Jeju Island, South Korea. Fish. Aquat. Sci. 2019, 22, 14. [Google Scholar] [CrossRef]
- Lock, E.J.; Arsiwalla, T.; Waagbø, R. Insect meal: A promising source of nutrients in the diet of Atlantic salmon (Salmo salar). In Proceedings of the Abstract Book Conference Insects to Feed The World, Ede, The Netherlands, 17 May 2014; Volume 14, pp. 14–17. [Google Scholar]
- Chaklader, M.R.; Siddik, M.A.; Fotedar, R.; Howieson, J. Insect larvae, Hermetia illucens in poultry by-product meal for barramundi, Lates calcarifer modulates histomorphology, immunity and resistance to Vibrio harveyi. Sci. Rep. 2019, 9, 16703. [Google Scholar] [CrossRef]
- Chaklader, M.R.; Howieson, J.; Siddik, M.A.; Foysal, M.J.; Fotedar, R. Supplementation of tuna hydrolysate and insect larvae improves fishmeal replacement efficacy of poultry by-product in Lates calcarifer (Bloch, 1790) juveniles. Sci. Rep. 2021, 11, 4997. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | Diet | |||
|---|---|---|---|---|
| FM70 (CON) | FM45 | FM35A | FM35B | |
| Sardine FM | 35.00 | 22.50 | 17.50 | 17.50 |
| Anchovy FM | 35.00 | 22.50 | 17.50 | 17.50 |
| Tankage meal | - | 8.00 | 11.50 | 10.50 |
| PBM 1 | - | 4.50 | 6.50 | 6.00 |
| TBM 2 | - | - | 1.00 | 1.60 |
| Wheat gluten | - | 5.50 | 4.70 | 4.70 |
| SPC 3 | - | 5.50 | 8.00 | 6.50 |
| Starch | 11.00 | 10.80 | 10.73 | 10.33 |
| Soybean meal | 12.00 | 12.00 | 12.00 | 12.50 |
| BSFL 4 | - | - | - | 3.50 |
| Fish oil | 3.30 | 4.30 | 4.20 | 2.50 |
| BO 5 | - | - | - | 0.50 |
| Lecithin | 0.50 | 0.50 | 0.70 | 0.70 |
| Betaine | - | 1.00 | 1.20 | 1.20 |
| Taurine | - | 0.50 | 0.80 | 0.80 |
| Met 99% | - | - | 0.07 | 0.07 |
| MCP 6 | 0.50 | 0.70 | 0.70 | 0.70 |
| Mineral mix | 1.00 | 1.00 | 1.00 | 1.00 |
| Vitamin mix | 1.00 | 1.00 | 1.00 | 1.00 |
| Vitamin C | 0.10 | 0.10 | 0.10 | 0.10 |
| Vitamin E | 0.10 | 0.10 | 0.10 | 0.10 |
| Choline | 0.50 | 0.50 | 0.70 | 0.70 |
| Proximate analysis (%, dry matter) | ||||
| Moisture | 6.22 | 8.35 | 6.64 | 6.10 |
| Crude protein | 56.90 | 56.80 | 57.00 | 56.30 |
| Crude lipid | 7.55 | 9.17 | 8.04 | 8.86 |
| Crude ash | 14.60 | 12.70 | 12.30 | 14.00 |
| Primer Name | Sequence (5′-3′) | Amplicon Size (bp) | Genbank No. | Ref. | |
|---|---|---|---|---|---|
| 18s rRNA | Forward | GGTCTGTGATGCCCTTAGATGTC | 107 | EF126037.1 | [25] |
| Reverse | AGTGGGGTTCAGCGGGTTAC | ||||
| IGF-1 | Forward | CGGCGCCTGGAGATGTACTG | 144 | AF016922.2 | [26] |
| Reverse | TGTCCTACGCTGTGCCT |
| Growth Performance | Diet | |||
|---|---|---|---|---|
| FM70 (CON) | FM45 | FM35A | FM35B | |
| FW (g) 1 | 1045.48 ± 200.77 | 1069.81 ± 156.08 | 1012.05 ± 210.21 | 980.90 ± 142.37 |
| WG (%) 2 | 192.81 ± 80.50 | 196.25 ± 77.84 | 181.10 ± 91.43 | 178.18 ± 83.92 |
| SGR (%) 3 | 0.67 ± 0.17 | 0.68 ± 0.14 | 0.64 ± 0.20 | 0.64 ± 0.17 |
| CF 4 | 2.31 ± 0.36 | 2.30 ± 0.25 | 2.30 ± 0.60 | 2.15 ± 0.23 |
| HSI (%) 5 | 1.90 ± 0.36 | 1.72 ± 0.46 | 1.66 ± 0.23 | 1.88 ± 0.37 |
| VSI (%) 6 | 4.18 ± 0.70 | 4.15 ± 0.49 | 4.02 ± 0.32 | 4.19 ± 0.56 |
| Survival (%) | 71.7 ± 0.04 | 70.7 ± 2.25 | 71.8 ± 1.79 | 73.7 ± 0.31 |
| Diet | ||||
|---|---|---|---|---|
| FM70 (CON) | FM45 | FM35A | FM35B | |
| Villus length (μm) | 1716.69 ± 156.66 | 1673.29 ± 127.24 | 1701.56 ± 149.23 | 1689.70 ± 102.67 |
| Muscular thickness (μm) | 224.06 ± 25.43 | 228.00 ± 27.37 | 222.88 ± 30.59 | 232.57 ± 19.88 |
| Goblet cell (cells/1000 µm2) | 358.60 ± 29.07 | 352.80 ± 28.44 | 365.00 ± 22.55 | 339.8 ± 15.35 |
| Enzyme (pg/mL) | Diet | |||
|---|---|---|---|---|
| FM70 (CON) | FM45 | FM35A | FM35B | |
| Trypsin | 92.74 ± 2.45 | 91.13 ± 3.73 | 89.59 ± 2.27 | 87.23 ± 1.94 |
| Chymotrypsin | 50.32 ± 1.49 a | 50.72 ± 0.88 a | 50.19 ± 1.44 a | 45.56 ± 2.17 b |
| Lipase | 29.66 ± 1.40 | 29.76 ± 0.89 | 30.93 ± 0.55 | 31.10 ± 0.64 |
| Diet | ||||
|---|---|---|---|---|
| FM70 (CON) | FM45 | FM35A | FM35B | |
| ACE | 207 ± 24 | 228 ± 43 | 270 ± 49 | 269 ± 92 |
| CHAO | 199 ± 23 | 222 ± 41 | 263 ± 46 | 262 ± 90 |
| Jackknife | 213 ± 25 | 240 ± 45 | 281 ± 51 | 282 ± 99 |
| Shannon | 1.82 ± 0.07 | 1.94 ± 0.41 | 2.24 ± 0.51 | 1.89 ± 0.31 |
| Simpson | 0.28 ± 0.02 | 0.26 ± 0.09 | 0.24 ± 0.13 | 0.27 ± 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, B.-S.; Park, S.-J.; Hwang, S.-Y.; Lee, Y.-I.; Lee, S.-H.; Hur, S.-W.; Lee, K.-J.; Nam, T.-J.; Song, J.-W.; Kim, J.-S.; et al. Effects of Decreasing Fishmeal as Main Source of Protein on Growth, Digestive Physiology, and Gut Microbiota of Olive Flounder (Paralichthys olivaceus). Animals 2022, 12, 2043. https://doi.org/10.3390/ani12162043
Seo B-S, Park S-J, Hwang S-Y, Lee Y-I, Lee S-H, Hur S-W, Lee K-J, Nam T-J, Song J-W, Kim J-S, et al. Effects of Decreasing Fishmeal as Main Source of Protein on Growth, Digestive Physiology, and Gut Microbiota of Olive Flounder (Paralichthys olivaceus). Animals. 2022; 12(16):2043. https://doi.org/10.3390/ani12162043
Chicago/Turabian StyleSeo, Bong-Seung, Su-Jin Park, So-Yeon Hwang, Ye-In Lee, Seung-Han Lee, Sang-Woo Hur, Kyeong-Jun Lee, Taek-Jeong Nam, Jin-Woo Song, Jae-Sig Kim, and et al. 2022. "Effects of Decreasing Fishmeal as Main Source of Protein on Growth, Digestive Physiology, and Gut Microbiota of Olive Flounder (Paralichthys olivaceus)" Animals 12, no. 16: 2043. https://doi.org/10.3390/ani12162043
APA StyleSeo, B.-S., Park, S.-J., Hwang, S.-Y., Lee, Y.-I., Lee, S.-H., Hur, S.-W., Lee, K.-J., Nam, T.-J., Song, J.-W., Kim, J.-S., Jang, W.-J., & Choi, Y.-H. (2022). Effects of Decreasing Fishmeal as Main Source of Protein on Growth, Digestive Physiology, and Gut Microbiota of Olive Flounder (Paralichthys olivaceus). Animals, 12(16), 2043. https://doi.org/10.3390/ani12162043