
Modulatory Effects of Arctostaphylos uva-urs Extract In Ovo Injected into Broiler Embryos Contaminated by Aflatoxin B1





Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Arctostaphylos uva-ursi In Vitro Analysis
2.1.1. Preparation and Extraction of Methanol Extract
2.1.2. Determination of Total Phenolic Content
2.1.3. Determination of Total Flavonoid Content
2.1.4. Estimation of In Vitro Antioxidant Activity
2.1.5. GC-MS Analysis
2.2. Arctostaphylos uva-ursi In Vivo Analysis
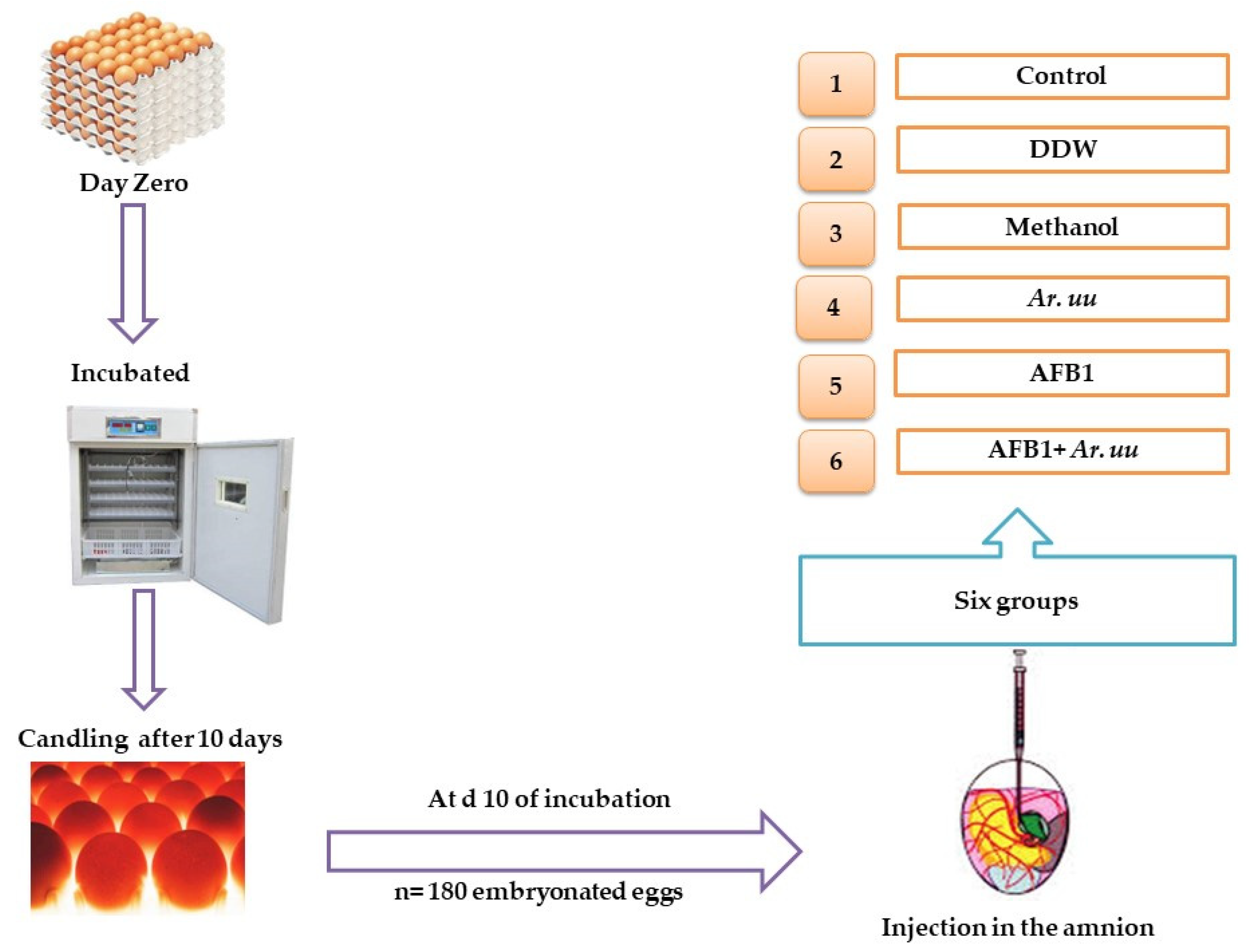
2.2.1. Eggs Incubation Protocol
2.2.2. Experimental Design
2.2.3. Embryonic Development Indices
Effect of Arctostaphylos uva-ursi Fruit Methanolic Extract and/or AFB1 In Ovo Injection on the Growth Rate of Broiler Chicks
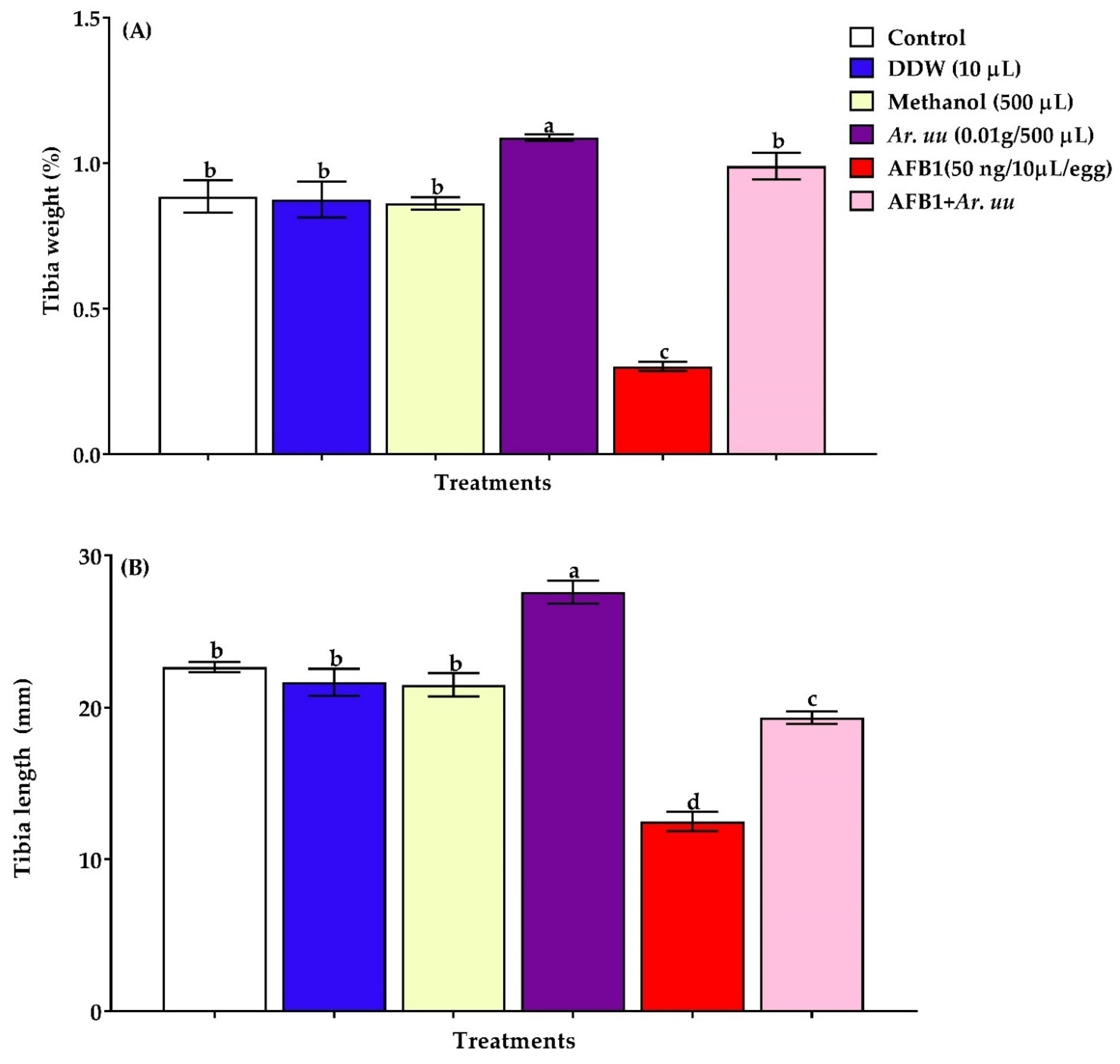
Effect of Arctostaphylos uva-ursi Fruit Methanolic Extract and/or AFB1 In Ovo Injection on the Development of Tibia of Broiler Chicks
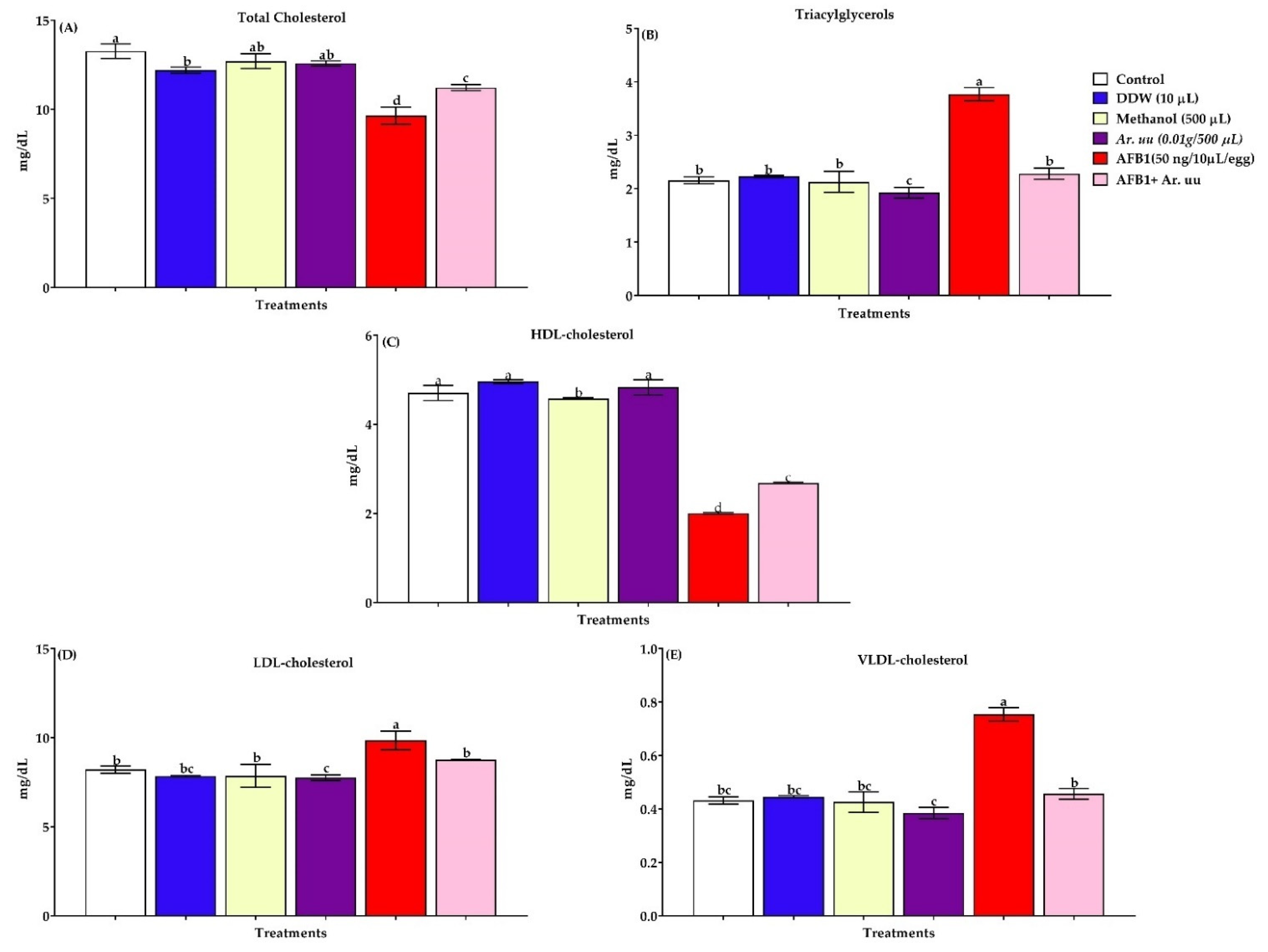
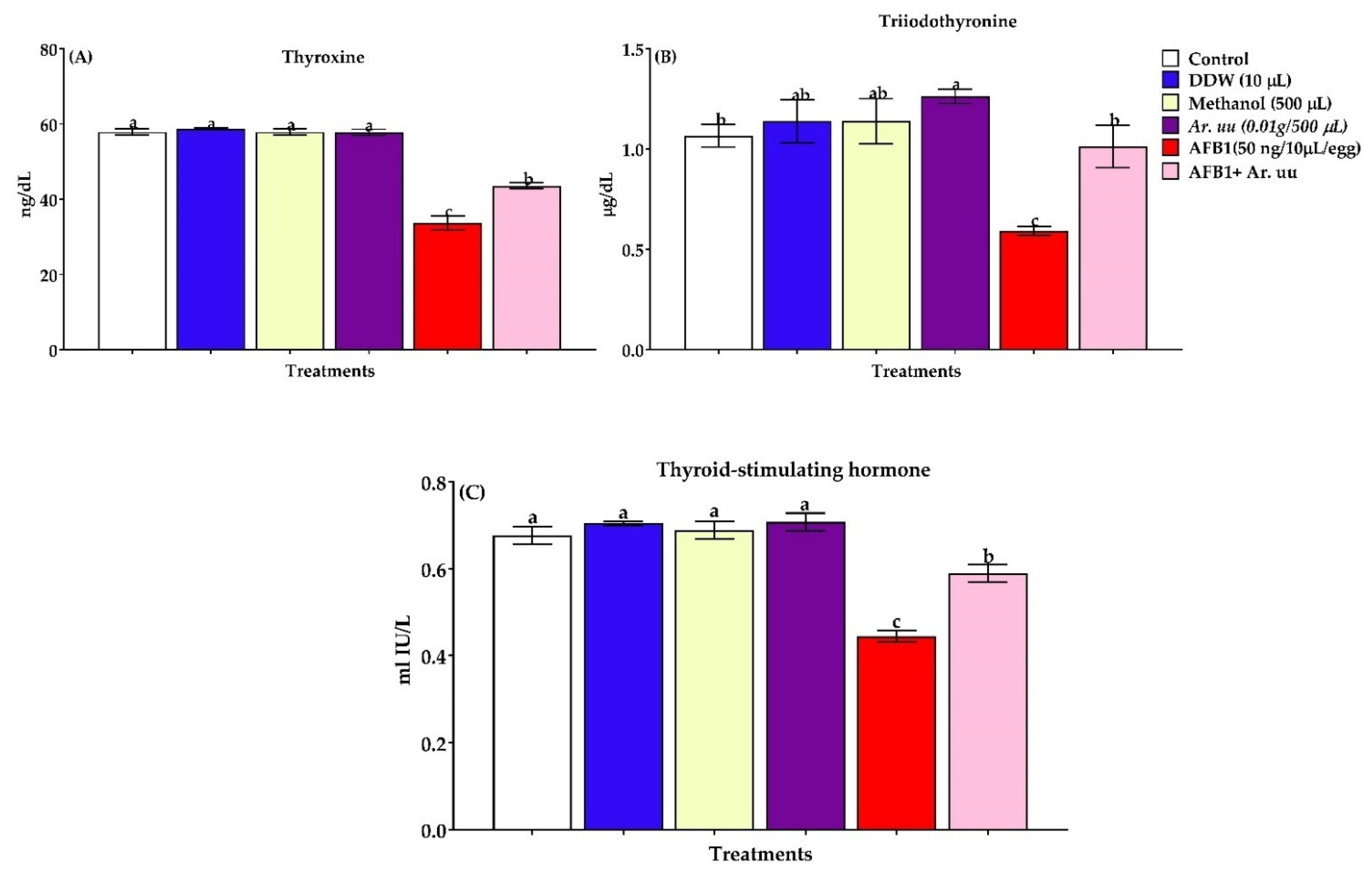
Effect of Arctostaphylos uva-ursi Fruit Methanolic Extract and/or AFB1 In Ovo Injection on Serum Biochemical Indices and Antioxidant Biomarkers in the Serum and Tissues
Effect of Arctostaphylos uva-ursi Fruit Methanolic Extract and/or AFB1 In Ovo Injection on the Spleen Histological, Immunohistochemistry and Apoptosis Examination
2.3. Statistical Analysis
3. Results
3.1. Arctostaphylos uva-ursi In Vitro Studies
3.1.1. Levels of Total Phenolic, Total Flavonoids, and Total Antioxidant Capacity
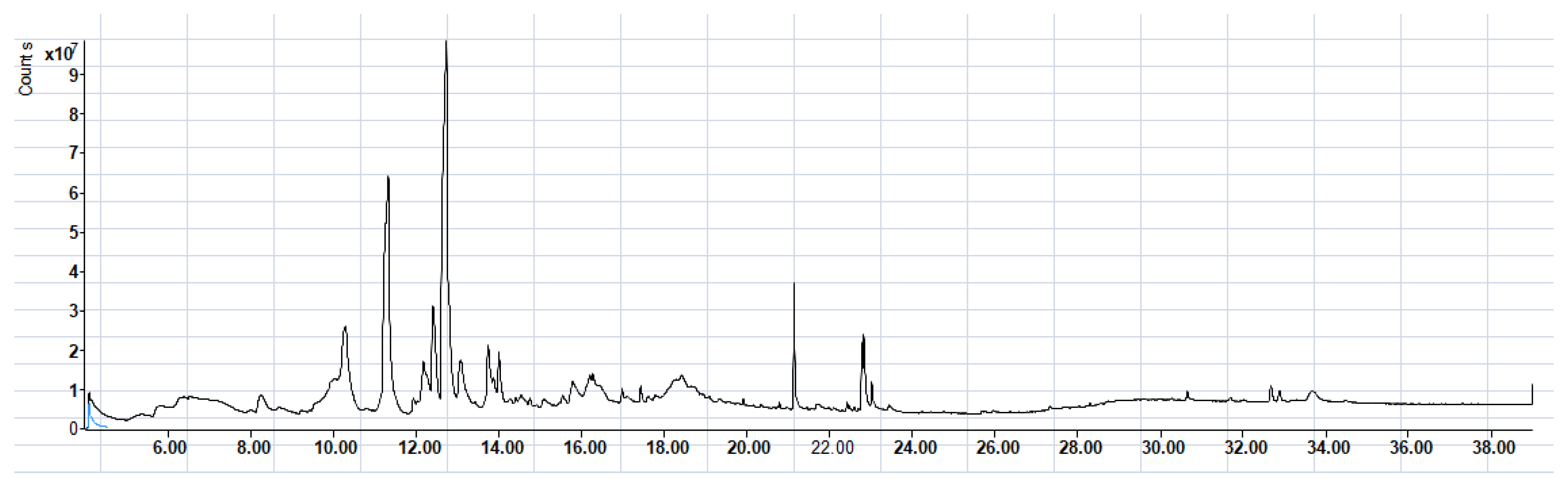
3.1.2. GC-MS Analysis
3.2. In Vivo Studies of Arctostaphylos uva-ursi Fruit Methanolic Extract
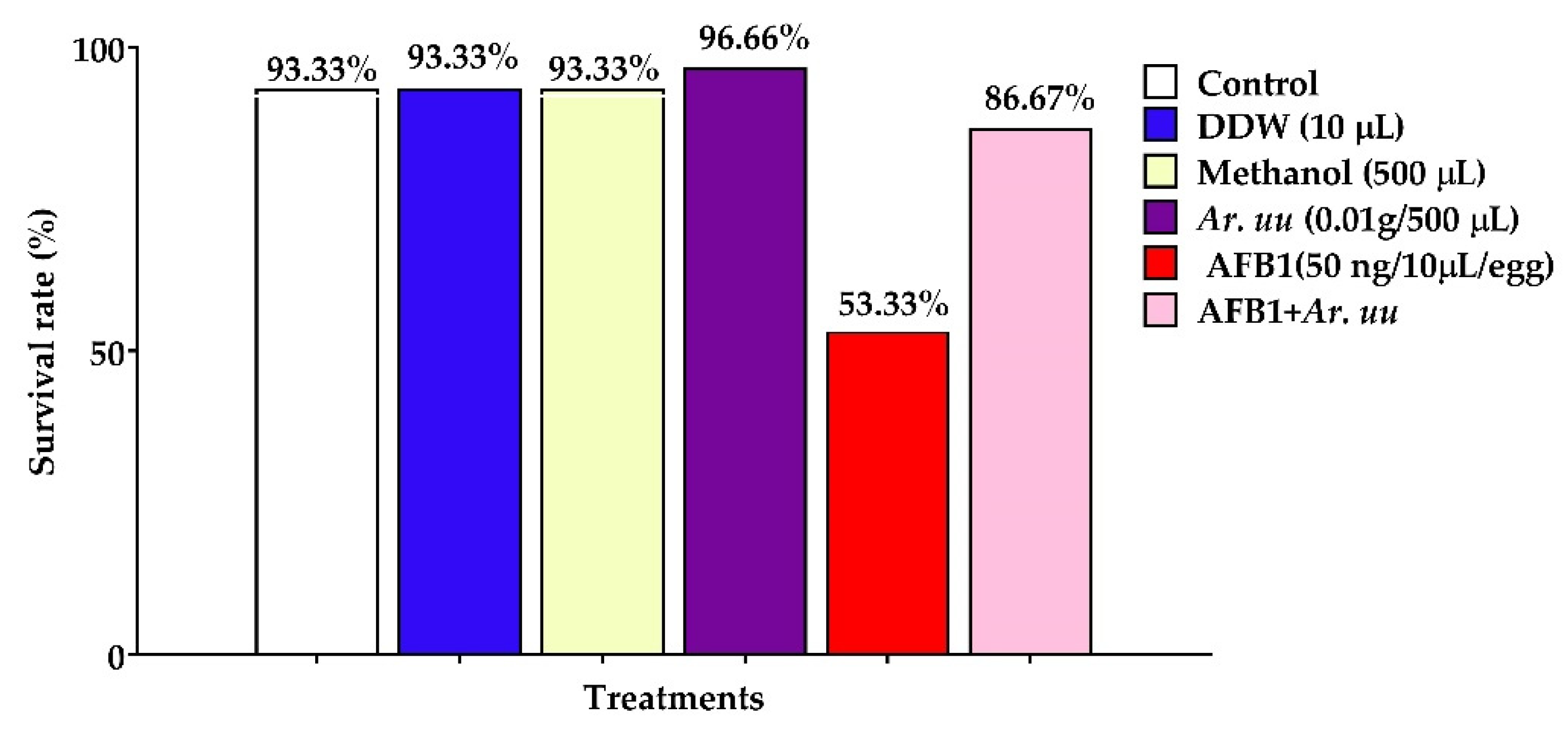
3.2.1. The Survival Rate of Broiler Embryos
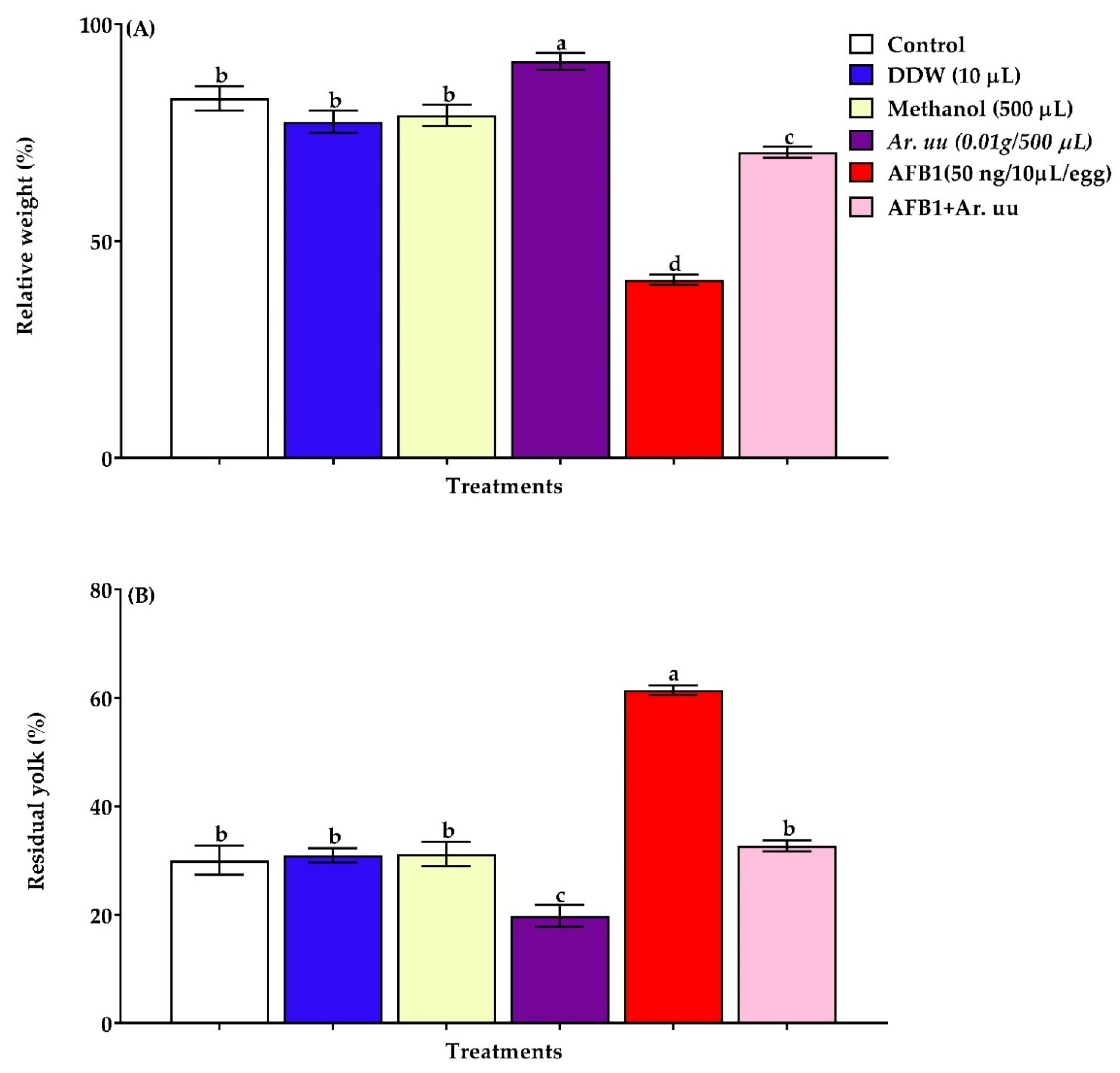
3.2.2. The Relative Residual Yolk Sac Weight and Relative Embryo Weight of Broiler Chicks
3.2.3. Effect of Arctostaphylos uva-ursi Fruit Methanolic Extract or/and AFB1 In Ovo Injection on the Tibia Bone Development of Broiler Embryos
3.2.4. Liver and Kidneys Functions of Broiler Chicks
3.2.5. Serum Lipid Profiles
3.2.6. Serum Thyroid Activity
3.2.7. Serum Total Immunoglobulins and Immunoglobulins Fractions
3.2.8. Antioxidant Indices, Activities of Dehydrogenase Enzymes in Serum, Liver, Kidneys, and Heart Tissues of Newly Hatched Broilers Chicks
3.2.9. Effect of Arctostaphylos uva-ursi Fruit Methanolic Extract and/or AFB1 In Ovo Injection on the Spleen Histology and Immunohistochemistry and Apoptosis Examination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tessari, E.N.C.; Oliveira, C.A.F.; Cardoso, A.L.S.P.; Ledoux, D.R.; Rottinghaus, G.E. Effects of aflatoxin B1 and fumonisin B1 on body weight, antibody titres and histology of broiler chicks. Br. Poult. Sci. 2006, 47, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Oguz, H.; Nizamlıoglu, F.; Dinc, I.; Uney, K.; Aydın, H. Determination of aflatoxin existence in mixed feed, wheat flour and bulgur samples. Eurasian J. Vet. Sci. 2011, 27, 171–175. [Google Scholar]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawal, S.; Kim, J.E.; Coulombe, R., Jr. Aflatoxin B1 in poultry: Toxicology, metabolism and prevention. Res. Vet. Sci. 2010, 89, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J. Regulatory aspects of mycotoxins in the European Community and USA. Fungi Mycotoxins Stored Prod. 1991, 36, 194–197. [Google Scholar]
- Murugesan, G.R.; Ledoux, D.R.; Naehrer, K.; Berthiller, F.; Applegate, T.J.; Grenier, B.; Phillips, T.D.; Schatzmayr, G. Prevalence and effects of mycotoxins on poultry health and performance, and recent development in mycotoxin counteracting strategies. Poult. Sci. 2015, 94, 1298–1315. [Google Scholar] [CrossRef]
- Haque, M.A.; Wang, Y.; Shen, Z.; Li, X.; Saleemi, M.K.; He, C. Mycotoxin contamination and control strategy in human, domestic animal and poultry: A review. Microb. Pathog. 2020, 142, 104095. [Google Scholar] [CrossRef]
- Peng, X.; Chen, K.; Chen, J.; Fang, J.; Cui, H.; Zuo, Z.; Deng, J.; Chen, Z.; Geng, Y.; Lai, W. Aflatoxin B1 affects apoptosis and expression of Bax, Bcl-2, and Caspase-3 in thymus and bursa of fabricius in broiler chickens. Environ. Toxicol. 2016, 31, 1113–1120. [Google Scholar] [CrossRef]
- Oswald, I.P.; Marin, D.E.; Bouhet, S.; Pinton, P.; Taranu, I.; Accensi, F.J.F.A. Immunotoxicological risk of mycotoxins for domestic animals. Food Addit. Contam. 2005, 22, 354–360. [Google Scholar] [CrossRef]
- Gündüz, N.; Oznurlu, Y. Adverse effects of aflatoxin B1 on skeletal muscle development in broiler chickens. Br. Poult. Sci. 2014, 55, 684–692. [Google Scholar] [CrossRef]
- Fouad, A.M.; Ruan, D.; El-Senousey, H.K.; Chen, W.; Jiang, S.; Zheng, C. Harmful Effects and Control Strategies of Aflatoxin B₁ Produced by Aspergillus flavus and Aspergillus parasiticus Strains on Poultry: Review. Toxins 2019, 11, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, J.W.; Bloom, S.E. Correlation between induction of xenobiotic metabolism and DNA damage from chemical carcinogens in the chick embryo in vivo. Carcinogenesis 1986, 7, 1101–1106. [Google Scholar] [CrossRef] [PubMed]
- Joshi, M.S.; Joshi, M.V. Effect of aflatoxin B1 on early embryonic stages of the chick Gallus domesticus cultured in vitro. Indian J. Exp. Biol. 1981, 19, 528–531. [Google Scholar]
- Geissler, F.; Faustman, E.M. Developmental toxicity of aflatoxin B1 in the rodent embryo in vitro: Contribution of exogenous biotransformation systems to toxicity. Teratology 1988, 37, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Celik, I.; Oguz, H.; Demet, O.; Boydak, M.; Donmez, H.H.M.; Sur, E.F.; Nizamlioglu, F. Embryotoxicity assay of aflatoxin produced by Aspergillus parasiticus NRRL 2999. Br. Poult. Sci. 2000, 41, 401–409. [Google Scholar] [CrossRef]
- Sur, E.; Celik, I. Effects of aflatoxin B1 on the development of the bursa of Fabricius and blood lymphocyte acid phosphatase of the chicken. Br. Poult. Sci. 2003, 44, 558–566. [Google Scholar] [CrossRef]
- Oznurlu, Y.; Celik, I.; Sur, E.; Ozaydın, T.; Oguz, H.; Altunba, K. Determination of the effects of aflatoxin B1 given In Ovo on the proximal tibial growth plate of broiler chickens: Histological, histometric and immunohistochemical findings. Avian Pathol. 2012, 41, 469–477. [Google Scholar] [CrossRef] [Green Version]
- Elwan, H.; Xie, C.; Miao, L.P.; Dong, X.; Zou, X.T.; Mohany, M.; Elnesr, S.S. Methionine alleviates aflatoxinb1-induced broiler chicks embryotoxicity through inhibition of caspase-dependent apoptosis and enhancement of cellular antioxidant status. Poult. Sci. 2021, 100, 101103. [Google Scholar] [CrossRef]
- Dwyer, M.R.; Kubena, L.F.; Harvey, R.B.; Mayura, K.; Sarr, A.B.; Buckley, S.; Bailey, R.H.; Phillips, T.D. Effects of inorganic adsorbents and cyclopiazonic acid in broiler chickens. Poult. Sci. 1997, 76, 1141–1149. [Google Scholar] [CrossRef]
- Phillips, T.D. Dietary clay in the chemoprevention of aflatoxin-induced disease. Toxicol. Sci. 1999, 52, 118–126. [Google Scholar] [CrossRef]
- Rosa, C.A.R.; Miazzo, R.; Magnoli, C.; Salvano, M.; Chiacchiera, S.M.; Ferrero, S.; Saenz, M.; Dalcero, A. Evaluation of the efficacy of bentonite from the south of Argentina to ameliorate the toxic effects of aflatoxin in broilers. Poult. Sci. 2001, 80, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Marchese, S.; Polo, A.; Ariano, A.; Velotto, S.; Costantini, S.; Severino, L. Aflatoxin B1 and M1: Biological properties and their involvement in cancer development. Toxins 2018, 10, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Nekeety, A.A.; Mohamed, S.R.; Hathout, A.S.; Hassan, N.S.; Aly, S.E.; Abdel-Wahhab, M.A. Antioxidant properties of Thymus vulgaris oil against aflatoxin-induce oxidative stress in male rats. Toxicon 2011, 57, 984–991. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Yang, H.; Jiao, Y.; Pang, Q.; Wang, Y.; Wang, M.; Shan, A.; Feng, X. Dietary Curcumin Alleviated Acute Ileum Damage of Ducks (Anas platyrhynchos) Induced by AFB1 through Regulating Nrf2-ARE and NF-κB Signaling Pathways. Foods 2021, 10, 1370. [Google Scholar] [CrossRef]
- Sarker, M.T.; Wan, X.; Yang, H.; Wang, Z. Dietary Lycopene Supplementation Could Alleviate Aflatoxin B1 Induced Intestinal Damage through Improving Immune Function and Anti-Oxidant Capacity in Broilers. Animals 2021, 11, 3165. [Google Scholar] [CrossRef]
- Isabelle, M.; Lee, B.L.; Lim, M.T.; Koh, W.P.; Huang, D.; Ong, C.N. Antioxidant activity and profiles of common vegetables in Singapore. Food Chem. 2010, 120, 993–1003. [Google Scholar] [CrossRef]
- Zomorodian, K.; Moein, M.; Lori, Z.G.; Ghasemi, Y.; Rahimi, M.J.; Bandegani, A.; Abbasi, N. Chemical composition and antimicrobial activities of the essential oil from myrtus communis leaves. J. Essent. Oil-Bear. Plants 2013, 16, 76–84. [Google Scholar] [CrossRef]
- Helander, I.M.; Alakomi, H.L.; Latva-Kala, K.; Mattila-Sandholm, T.; Pol, I.; Smid, E.J.; Gorris, L.G.M.; von Wright, A. Characterization of the action of selected essential oil components on Gram-negative bacteria. J. Agric. Food Chem. 1998, 46, 3590–3595. [Google Scholar] [CrossRef]
- Farag, R.S.; Daw, Z.Y.; Abo-Raya, S.H. Influence of some spice essential oils on Aspergillus parasiticus growth and production of aflatoxins in a synthetic medium. J. Food Sci. 1989, 54, 74–76. [Google Scholar] [CrossRef]
- Bowles, B.L.; Miller, A.J. Antibotulinal properties of selected aromatic and aliphatic aldehydes. J. Food Prod. 1993, 56, 788–794. [Google Scholar] [CrossRef]
- Roby, M.H.H.; Sarhan, M.A.; Selim, K.A.H.; Khalel, K.I. Evaluation of antioxidant activity, total phenols and phenolic compounds in thyme (Thymus vulgaris L.), sage (Salvia officinalis L.), and marjoram (Origanum majorana L.) extracts. Ind. Crops Prod. 2013, 43, 827–831. [Google Scholar] [CrossRef]
- Naczk, M.; Pegg, R.B.; Amarowicz, R. Protein-precipitating capacity of bearberry-leaf (Arctostaphylos uva-ursi L. Sprengel) polyphenolics. Food Chem. 2011, 124, 1507–1513. [Google Scholar] [CrossRef]
- Pegg, R.B.; Amarowicz, R.; Barl, B. Application of plant polyphenolics in model and meat systems. In Proceedings of the 47th International Congress of Meat Science and Technology, Krakow, Poland, 26–31 August 2001; pp. 234–235. [Google Scholar]
- Amarowicz, R.; Pegg, R.B.; Rahimi-Moghaddam, P.; Barl, B.; Weil, J.A. Free radical scavenging capacity and antioxidant activity of selected plant species from the Canadian prairies. Food Chem. 2004, 84, 551–562. [Google Scholar] [CrossRef]
- Dykes, G.A.; Amarowicz, R.; Pegg, R.B. Enhancement of nisin antibacterial activity by a bearberry (Arctostaphylos uva-ursi) leaf extract. Food Microbiol. 2003, 20, 211–216. [Google Scholar] [CrossRef]
- Pegg, R.B.; Amarowicz, R.; Naczk, M.; Shahidi, F. PHOTOCHEM® for determination of antioxidant capacity of plant extract. ACS Symp. Ser. 2009, 956, 140–158. [Google Scholar]
- Barl, B.; Loewen, D.; Svendsen, E. Saskatchewan Herb Database; Department of Horticulture Science, University of Saskatchewan: Saskatoon, SK, Canada, 1996. [Google Scholar]
- Youssef, H.A.; Ali, S.M.; Sanad, M.I.; Dawood, D.H. Chemical Investigation of Flavonoid, Phenolic Acids Composition and Antioxidant Activity of Mexican Marigold (Tagetes erecta L.) Flowers. Egypt. J. Chem. 2020, 63, 2605–2615. [Google Scholar]
- Limmongkon, A.; Janhom, P.; Amthong, A.; Kawpanuk, M.; Nopprang, P.; Poohadsuan, J.; Somboon, T.; Saijeen, S.; Surangkul, D.; Srikummool, M.; et al. Antioxidant activity, total phenolic, and resveratrol content in five cultivars of peanut sprouts. Asian Pac. J. Trop Biomed. 2017, 7, 332–338. [Google Scholar] [CrossRef]
- Munhoz, V.M.; Longhini, R.; Souza, J.R.P.; Zequi, J.A.C.; Mello, E.V.S.L.; Lopes, G.C.; Mello, J.C.P. Extraction of flavonoids from Tagetes patula: Process optimization and screening for biological activity. Rev. Bras. Farmacog. 2014, 24, 576–583. [Google Scholar] [CrossRef] [Green Version]
- Dawood, H.D.; Mohamed, S.E.; Amr, N.; Mohamed, A.T. Extraction and chemical characterization of novel water-soluble polysaccharides from two palm species and their antioxidant and antitumor activities. Egypt. J. Basic Appl. Sci. 2020, 7, 141–158. [Google Scholar] [CrossRef]
- Elwan, H.A.M.; Dawood, H.D.; El-Shafei, S.M.A.; Abd El-Rahman, A.A.; Abdel-Latif, S.A.; Mohany, M.; Alqahtani, F.; Alqahtani, S.; Al-Rejaie, S.S. The potential role of citrus limon powder as a natural feed supplement to boost the productive performance, antioxidant status, and blood biochemistry of growing rabbits. Animals 2019, 9, 426. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.Z.; Hoque, M.M.; Asif-Ul-Alam, S.M.; Monalisa, K. Chemical composition, antioxidant capacities and storage stability of Citrus macroptera and Garciniape dunculata fruits. Emir J. Food Agric. 2015, 27, 275–282. [Google Scholar] [CrossRef] [Green Version]
- Campêlo, M.C.S.; Medeiros, J.M.S.; Silva, J.B.A. Natural products in food preservation. Int. Food Res. J. 2019, 26, 41–46. [Google Scholar]
- Ghafar, M.F.D.; Nagendra, P.K.; Weng, K.K.; Ismail, A. Flavonoid, hesperidine, total phenolic contents and antioxidant activities from Citrus species. Afr. J. Biotechnol. 2010, 9, 326–330. [Google Scholar]
- Azman, N.A.M.; Gallego, M.G.; Segovia, F.; Abdullah, S.; Shaarani, S.M.; Almajano Pablos, M.P. Study of the properties of Bearberry leaf extract as a natural antioxidant in model foods. Antioxidants 2016, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Dragana, M.V.; Miroslav, R.P.; Branka, B.R.G.; Sava, M.V.; Ljiljana, R. In vitro efficacy of extracts of Arctostaphylos uva-ursi L. on clinical isolated Escherichia coli and Enterococcus faecalis strains. Kragujev. J. Sci. 2013, 35, 107–113. [Google Scholar]
- Deng, Y.; Yang, G.; Yue, J.; Qian, B.; Liu, Z.; Wang, D.; Zhong, Y.; Zhao, Y. Influences of ripening stage and extracting solvents on the polyphenolic compounds, antimicrobial and antioxidant activities of blue berry leaf extracts. Food Control 2014, 38, 184–191. [Google Scholar] [CrossRef]
- Barapatre, A.; Meena, A.S.; Mekala, S.; Das, A.; Jha, H. In vitro evaluation of antioxidant and cytotoxic activities of ligninfractions extracted from Acacia nilotica. Int. J. Biol. Micromol. 2016, 86, 443–453. [Google Scholar] [CrossRef]
- Ling, Z.; Jianping, C.; Jianyu, S.; Lin, L.; Songqing, H.; Bing, L.; Xia, Z.; Zhenbo, X.; Tianfeng, C. In Vitro Antioxidant and Antiproliferative Activities of 5-Hydroxymethylfurfural. J. Agric. Food Chem. 2013, 61, 10604–10611. [Google Scholar]
- Jelinek, R.; Peterka, M.; Rychter, Z. Chick embryotoxicity screening test—130 substances tested. Indian J. Exp. Biol. 1985, 23, 588–595. [Google Scholar]
- Yin, H.; Jiang, M.; Peng, X.; Cui, H.; Zhou, Y.; He, M.; Zuo, Z.; Ouyang, P.; Fan, J.; Fang, J. The molecular mechanism of G2/M cell cycle arrest induced by AFB1 in the jejunum. Oncotarget 2016, 7, 35592–35606. [Google Scholar] [CrossRef] [Green Version]
- Del Vesco, A.P.; Gasparino, E.; Grieser, O.D.; Zancanela, V.; Soares, M.A.; Neto, A.R. Effects of methionine supplementation on the expression of oxidative stress-related genes in acute heat stress exposed broilers. Br. J. Nutr. 2015, 113, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Khlangwiset, P.; Shephard, G.S.; Wu, F. Aflatoxins and growth impairment: A review. Crit. Rev. Toxicol. 2011, 41, 740–755. [Google Scholar] [CrossRef] [PubMed]
- R’ehault-Godbert, S.; Mann, K.; Bourin, M.; Brionne, A.; Nys, Y. Effect of embryonic development on the chicken egg yolk plasma proteome after 12 d of incubation. J. Agric. Food Chem. 2014, 62, 2531–2540. [Google Scholar] [CrossRef] [PubMed]
- Yadgary, L.; Wong, E.A.; Uni, Z. Temporal transcriptome analysis of the chicken embryo yolk sac. BMC Genom. 2014, 15, 690–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yalcin, S.; Bağdatlioğlu, N.; Bruggeman, V.; Babacanoğlu, E.; Uysal, İ.; Buyse, J.; Decuypere, E.; Siegel, P.B. Acclimation to heat during incubation. 2. Embryo composition and residual egg yolk sac fatty acid profiles in chicks. Poult. Sci. 2008, 87, 1229–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huff, W.E.; Doerr, J.A.; Hamilton, P.B.; Hamann, D.D.; Peterson, R.E.; Ciegler, A. Evaluation of bone strength during aflatoxicosis and ochratoxicosis. Appl. Environ. Microbiol. 1980, 40, 102–107. [Google Scholar] [CrossRef] [Green Version]
- Chaudhry, Z.I. Effects of Aflatoxicosis on Chick Muscle, Fat and Bone Growth and Possible Reversal of Aflatoxicosis by Nandrolonedecanoate. Ph.D. Thesis, Punjab University, Lahore, Pakistan, 1996. [Google Scholar]
- Khan, W.A.; Khan, M.Z.; Khan, A.; Hassan, Z.U.; Rafique, S.; Saleemi, M.K.; Ahad, A. Dietary vitamin E in White Leghorn layer breeder hens: A strategy to combat aflatoxin B1-induced damage. Avian Pathol. 2014, 43, 389–395. [Google Scholar] [CrossRef]
- Aydin, M.F.; Celik, I.; Sur, E.; Ozparlak, H.; Telatar, T. Effects of in ovo given aflatoxin Bı on the chick hatching weight. Eurasian J. Vet. Sci. 2005, 21, 85–89. [Google Scholar]
- Seebaluck-Sandoram, R.; Lall, N.; Fibrich, B.; Van Staden, A.B.; Mahomoodally, F. Antibiotic-potention, antioxidant, cytotoxic, antiinflammatory and anti-acetylcholinesterase potential of Antidesma madagascariense Lam. (Euphorbiaceae). S. Afr. J. Bot. 2017, 111, 194–201. [Google Scholar] [CrossRef]
- Supasuteekul, C.; Nonthitipong, W.; Tadtong, S.; Likhitwitayawuid, K.; Tengamnuay, P.; Sritularak, B. Antioxidant, DNA damage protective, neuroprotective, and α-glucosidase inhibitory activities of a flavonoid glycoside from leaves of Garcinia gracilis. Rev. Bras. Farmacog. 2016, 26, 312–320. [Google Scholar] [CrossRef] [Green Version]
- Lourenço, S.C.; Moldão-Martins, M.; Alves, V.D. Antioxidants of natural plant origins: From sources to food industry applications. Molecules 2019, 24, 4132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanescu, B.E.; Szabo, K.; Mocan, A.; Crisan, G. Phenolic compounds from five Ericaceae species leaves and their related bioavailability and health benefits. Molecules 2019, 24, 2046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.P.; Calomme, M.; Lasure, A.; De Bruyne, T.; Pieters, L.; Vlietinck, A.; Vanden Berghe, D.A. Structure–activity relationship of flavonoids with superoxide scavenging activity. Biol. Trace Elem. Res. 1995, 47, 327–331. [Google Scholar] [CrossRef]
- Nijveldt, R.J.; van Nood, E.; van Hoorn, D.E.; Boelens, P.G.; van Norren, K.; van Leewen, P.A. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar] [CrossRef]
- Masella, R.; Di Benedetto, R.; Varì, R.; Filesi, C.; Giovannini, C. Novel mechanisms of natural antioxidant compounds in biological systems: Involvement of glutathione and glutathione-related enzymes. J. Nutr. Biochem. 2005, 16, 577–586. [Google Scholar] [CrossRef]
- Spencer, J.P.; Rice-Evans, C.; Williams, R.J. Modulation of pro-survival Akt/PKB and ERK1/2 signalling cascades by quercetin and its in vivo metabolites underlie their action on neuronal viability. J. Biol. Chem. 2003, 278, 34783–34793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, S.O.; Simon, S.; Chae, K.; Metzler, M.; Korach, K.S. Phytoestrogens and their human metabolites show distinct agonistic and antagonistic properties on estrogen receptor alpha (ERalpha) and ERbeta in human cells. Toxicol. Sci. 2004, 80, 14–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, P.M.; Lane, D.P. Inhibitors of cyclin-dependent kinases as anticancer therapeutics. Curr. Med. Chem. 2000, 7, 1213–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallerath, T.; Poleo, D.; Li, H.; Fostermann, U. Red wine increases the expression of human endothelial nitric oxide synthase: A mechanism that may contribute to its beneficial cardiovascular effects. J. Am. Coll. Cardiol. 2003, 41, 471–478. [Google Scholar] [CrossRef] [Green Version]
- Murphy, K.J.; Chronopoulos, A.K.; Singh, I.; Francis, M.A.; Moriarty, H.; Pike, M.J.; Sinclair, A.J. Dietary flavanols and procyanidin oligomers from cocoa (Teobroma cacao) inhibit platelet function. Am. J. Clin. Nutr. 2003, 77, 1466–1473. [Google Scholar] [CrossRef] [Green Version]
- Birt, D.F.; Hendrich, S.; Wang, W. Dietary agents in cancer prevention: Flavonoids and isoflavonoids. Pharmacol. Ther. 2001, 90, 157–177. [Google Scholar] [CrossRef]
- Way, T.D.; Kao, M.C.; Lin, J.K. Degradation of HER2/neu by apigenin induces apoptosis through cytochrome c release and caspase-3 activation in HER2/neu-overexpressing breast cancer cells. FEBS Lett. 2005, 579, 145–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelloff, G.J.; Crowell, J.A.; Steele, V.E.; Lubet, R.A.; Malone, W.A.; Boone, C.W.; Sigman, C.C. Progress in cancer chemoprevention: Development of diet-derived chemopreventive agents. J. Nutr. 2000, 130, 467S–471S. [Google Scholar] [CrossRef]
- Van Zanden, J.J.; Geraets, L.; Worteboer, H.M.; Van Bladeren, P.J.; Rietjens, N.H.P.; Cnubben, N.H.P. Structural requirements for the flavonoidmediated modulation of glutathione S-transferase and GS-X pump activity in MCF7 breast cancer cells. Biochem. Pharmacol. 2004, 67, 1607–1617. [Google Scholar] [CrossRef] [PubMed]
- Depeint, F.; Gee, J.M.; Williamson, G.; Johnson, I.T. Evidence for consistent patterns between flavonoid structures and cellular activities. Proc. Nutr. Soc. 2002, 61, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Fukao, T.; Hosono, T.; Misawa, S.; Seki, T.; Ariga, T. The effects of allyl sulfides on the induction of phase II detoxification enzymes and liver injury by carbon tetrachloride. Food. Chem. Toxicol. 2004, 42, 743–749. [Google Scholar] [CrossRef]
- Kumar, G.; Xu, B.J. A critical review on polyphenols and health benefits of black soybeans. Nutrients 2017, 9, 455. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, T.; Yokozawa, T. Direct scavenging of nitric oxide and superoxide by green tea. Food Chem. Toxicol. 2002, 40, 1745–1750. [Google Scholar] [CrossRef]
- Yiannakopoulou, E.C. Targeting oxidative stress response by green tea polyphenols: Clinical implications. Free Radic. Res. 2013, 47, 667–671. [Google Scholar] [CrossRef]
- Tsao, R.; Li, H. Antioxidant properties in vitro and in vivo: Realistic assessments of efficacy of plant extracts. Plant Sci. Rev. 2012, 7, 11–13. [Google Scholar]
- Lambert, J.D.; Elias, R.J. The antioxidant and pro-oxidant activities of green tea polyphenols: A role in cancer prevention. Arch. Biochem. Biophys. 2010, 501, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubena, L.F.; Harvey, R.B.; Huff, W.E.; Corrier, D.E.; Phillips, T.D.; Rottinghaus, G.E. Efficacy of a hydrated sodium calcium aluminosilicate to reduce the toxicity of aflatoxin and T-2 toxin. Poult. Sci. 1990, 69, 1078–1086. [Google Scholar] [CrossRef] [PubMed]
- Ellis, W.O.; Smith, J.P.; Simpson, B.K. Aflatoxin in food: Occurrence, biosynthesis, effects on organisms, detection and methods of control. Crit. Rev. Food Sci. Nutr. 1991, 30, 403–439. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, E.; Yalcinkaya, I.; Kanbur, M.U.R.A.T.; Cinar, M.; Oruc, E. Effects of yeast glucomannan on performance, some biochemical parameters and pathological changes in experimental aflatoxicosis in broiler chickens. Rev. Med. Vet. 2011, 162, 413–420. [Google Scholar]
- Büchau, A.S.; Gallo, R.L. Innate immunity and antimicrobial defense systems in psoriasis. Clin. Dermatol. 2007, 25, 616–624. [Google Scholar] [CrossRef] [Green Version]
- Donmez, N.; Keskin, E. The effect of aflatoxin and glucomannan on some antioxidants and biochemical parameters in rabbits. Acta Vet. 2008, 58, 307–313. [Google Scholar] [CrossRef]
- Haschek, W.M.; Voss, K.A.; Beasley, V.R. Selected Mycotoxins Affecting Animal and Human Health, 2nd ed.; Academic Press: New York, NY, USA, 2002; Volume 1, pp. 645–699. [Google Scholar]
- Agag, B.I. Mycotoxins in foods and feeds: 1-aflatoxins. Assiut Univ. Bull. Environ. Res. 2004, 7, 173–205. [Google Scholar]
- Wayne, L.B. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed Sci. Technol. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Elnesr, S.S.; Abdel-Razik, A.R.H.; Elwan, H.A. Impact of humate substances and Bacillus subtilis PB6 on thyroid activity and histomorphometry, iron profile and blood haematology of quail. J. Anim. Physiol. Anim. Nutr. 2022, 106, 110–117. [Google Scholar] [CrossRef]
- Verma, R.J. Aflatoxin cause DNA damage. Int. J. Hum. Genet. 2004, 4, 231–236. [Google Scholar] [CrossRef]
- Surai, P.F. Natural antioxidants and mycotoxins. In Natural Antioxidants in Avian Nutrition and Reproduction; Nottingham University Press: Nottingham, UK, 2002; pp. 455–489. [Google Scholar]
- Bozakova, N.; Popova-Ralcheva, S. Thyroid hormones level and relative liver weight in male turkeys in relationship with their welfare. Biotechnol. Anim. Husb. 2007, 23, 1511–1518. [Google Scholar] [CrossRef] [Green Version]
- Berry, M.J.; Larsen, P.R. The role of selenium in thyroid hormone action. Endocr. Rev. 1992, 13, 207–219. [Google Scholar] [PubMed]
- Rose, S.R. Disorders of thyrotropin synthesis, secretion, and function. Curr. Opin. Pediatr. 2000, 12, 375–381. [Google Scholar] [CrossRef]
- Noyanm, A. Hormonlar. In Yasamda ve Hekimlikte Fizyoloji; Plume: Ankara, Turkey, 1993; pp. 977–1033. [Google Scholar]
- Markou, K.; Georgopoulos, N.; Kyriazopoulou, V.; Vagenakis, A.G. Iodineinduced hypothyroidism. Thyroid 2011, 11, 501–510. [Google Scholar] [CrossRef]
- Eraslan, G.; Akdoğan, M.; Liman, B.C.; Kanbur, M.; Delibaş, N. Effects of dietary aflatoxin and hydrate sodium calcium aluminosilicate on triiodothyronine, thyroxine, thyrotrophin and testosterone levels in quails. Turk. J. Vet. Anim. Sci. 2006, 30, 41–45. [Google Scholar]
- Lee, K.I.; Rhee, S.H.; Park, K.Y. Anticancer activity of phytol and eicosatrienoic acid identified from Perilla leaves. J. Korean Soc. Food Sci. Nutr. 1999, 28, 1107–1112. [Google Scholar]
- Rastogi, R.; Srivastava, A.K.; Rastogi, A.K. Long term effect of aflatoxin B1 on lipid peroxidation in rat liver and kidney: Effect of picroliv and silymarin. Phytother. Res. 2001, 15, 307–310. [Google Scholar] [CrossRef]
- Carew, L.B.; Evarts, K.G.; Alster, F.A. Growth, feed intake, and plasma thyroid hormone levels in chicks fed dietary excesses of essential amino acids. Poult. Sci. 1998, 77, 295–298. [Google Scholar] [CrossRef]
- Johri, T.S.; Sadagopan, V.R.; Shrivastava, M.A.; Jumdar, S. Effect of dietary aflatoxin on the performance of purebred broiler chicks. Indian J. Anim. Sci. 1990, 60, 1246–1248. [Google Scholar]
- Graczyk, S.; Kotoński, B.; Malicki, A.; Orda, J.; Zawadzki, W. The thyroxine levels and blood serum profiles in ducklings after chronic aflatoxin B1 administration. Med. Vet. 2002, 1, 21–29. [Google Scholar]
- Zhao, J.; Shirley, R.B.; Dibner, J.D.; Uraizee, F.; Officer, M.; Kitchell, M.; Vazquez-Anon, M.; Knight, C.D. Comparison of hydrated sodium calcium aluminosilicate and yeast cell wall on counteracting aflatoxicosis in broiler chicks. Poult. Sci. 2010, 89, 2147–2156. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.H.; Mohamed, M.H. Haemato-biochemical and pathological studies on aflatoxicosis and treatment of broiler chicks in Egypt. Vet. Ital. 2009, 45, 323–337. [Google Scholar]
- Sakhare, P.S.; Harne, S.D.; Kalorey, D.R.; Warke, S.R.; Bhandarkar, A.G.; Kurkure, N.V. Effect of Toxiroak® polyherbal feed supplement during induced aflatoxicosis, ochratoxicosis and combined mycotoxicoses in broilers. Vet. Arh. 2007, 77, 129–146. [Google Scholar]
- Ibrahim, I.K.; Shareef, A.M.; Al-Joubory, K.M.T. Ameliorative effects of sodium bentonite on phagocytosis and Newcastle disease antibody formation in broiler chickens during aflatoxicosis. Res. Vet. Sci. 2000, 69, 119–122. [Google Scholar] [CrossRef]
- Ledoux, D.R.; Rottinghaus, G.E.; Bermudez, A.J.; Alonso-Debolt, M. Efficacy of hydrated sodium calcium aluminosilicate to ameliorate the toxic effects of aflatoxin in broiler chicks. Poult. Sci. 1999, 78, 204–210. [Google Scholar] [CrossRef]
- Assar, M.H.; Attia, K.M.; Eid, Y. Evaluation of the ability of a feed additive to ameliorate the adverse effects of aflatoxins in broiler chickens. Alex. J. Vet. Sci. 2018, 56, 1–18. [Google Scholar] [CrossRef]
- Shen, H.M.; Ong, C.N.; Shi, C.Y. Involvement of reactive oxygen species in aflatoxin B1 induced cell injury in cultured rat hepatocytes. Toxicology 1995, 99, 115–123. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Methanol Extract | TPC mg GAE/g DW | TFC mg QE/g DW | Antioxidant Activity (DPPH %) |
|---|---|---|---|
| Arctostaphylos uva-ursi | 212.88 | 91.28 | 60.25 |
| RT * (Min) | Compound Name | Structure | Compound Nature | Peak Area | Peak Area (%) |
|---|---|---|---|---|---|
| 10.295 | Thymine |  | Alkaloids | 109,350,462.3 | 17.40 |
| 11.323 | 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl- |  | Phenols | 153,643,563.1 | 24.45 |
| 12.171 | Phloroglucinol, trimethylsilyl ether | 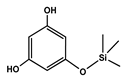 | Phenols | 3,447,892.805 | 0.55 |
| 12.407 | Benzofuran, 2,3-dihydro- |  | Coumarins | 71,308,776.28 | 11.34 |
| 12.727 | 5-Hydroxymethylfurfural |  | Glycosides | 275,409,104.1 | 43.83 |
| 13.093 | 1,2-Benzenediol, 3-methoxy- | 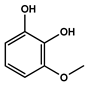 | Phenols | 3,877,902.97 | 0.62 |
| 13.748 | 2-Methoxy-4-vinylphenol | 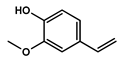 | phenols | 4,725,079.814 | 0.75 |
| 14.222 | trans-3-Trifluoromethylcinnamic acid, 4-nitrophenyl ester |  | Cinnamic derivatives | 134,564.4497 | 0.021 |
| 15.784 | Succinic acid, 3-nitrobenzyl pentyl ester |  | Organic acids | 191,184.1509 | 0.030 |
| 16.270 | Phenol, 2,6-bis(1,1-dimethylethyl)- | 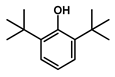 | Phenols | 777,540.8755 | 0.12 |
| 21.138 | n-Hexadecanoic acid | 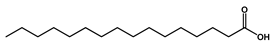 | Long-chain fatty acid | 2,705,961.767 | 0.43 |
| 22.821 | 9-Octadecenoic acid, (E)- | 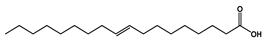 | Unsaturated fatty acid | 2,720,133.219 | 0.42 |
| Items | Treatments | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| Control | DDW | Methanol | Ar. uu. | AFB1 | AFB1 + Ar. uu | |||
| Total Protein (g/L) | 6.838 a | 6.806 a | 6.166 a | 6.527 a | 5.687 b | 6.398 a | 0.196 | 0.04 |
| Albumin (g/L) | 2.515 b | 2.631 b | 2.357 b | 2.210 b | 3.950 a | 2.833 b | 0.313 | 0.01 |
| Globulin (g/L) | 4.323 a | 4.175 a | 3.809 a | 4.317 a | 1.737 b | 3.565 a | 0.270 | 0.01 |
| Alb/Glo ratio | 0.582 b | 0.630 b | 0.619 b | 0.512 b | 2.274 a | 0.795 b | 0.109 | 0.01 |
| GGT (IU) | 0.650 c | 0.670 c | 0.688 c | 0.676 c | 1.340 a | 0.836 b | 0.002 | 0.01 |
| AKP (IU) | 549.33 c | 524.33 c | 577.73 c | 538.67 c | 1576.00 a | 886.67 b | 27.72 | 0.01 |
| GPT (IU) | 22.33 c | 24.24 c | 25.18 c | 23.40 c | 35.93 a | 28.67 b | 1.118 | 0.01 |
| GOT (IU) | 2.513 c | 2.716 c | 2.889 c | 12.963 c | 6.856 a | 3.622 b | 0.019 | 0.01 |
| Urea Nitrogen | 1.22 c | 1.19 c | 1.22 c | 1.24 c | 2.15 a | 1.53 b | 0.004 | 0.01 |
| Uric acid (µmol/L) | 0.831 c | 0.809 c | 0.882 c | 0.848 c | 1.99 a | 0.457 b | 0.110 | 0.01 |
| C K * (mmol/L) | 1.100 c | 1.06 c | 1.09 c | 1.01 c | 2.03 a | 1.40 b | 0.004 | 0.01 |
| Items | Treatment Groups | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| Control | DDW | Methanol | AR. UU. | AFB1 | AFB1 + Ar. uu. | |||
| Serum | ||||||||
| TAOC (U/mg prot.) | 6.04 a | 6.24 a | 6.30 a | 5.66 b | 2.66 d | 4.67 c | 0.016 | 0.01 |
| SOD (U/mg prot.) | 102.33 b | 100.66 b | 102.14 b | 113.66 a | 76.00 d | 88.00 c | 5.503 | 0.01 |
| GSH (µmol/g prot.) | 13.21 b | 9.03 c | 6.73 c | 12.24 b | 8.75 c | 17.59 a | 1.187 | 0.01 |
| GSH-Px activity (U) | 958.1 bc | 917.6 bc | 937.0 bc | 2579.3 a | 621.6 c | 1306.1 b | 61.371 | 0.01 |
| Catalase (U/mg prot.) | 47.02 b | 44.78 b | 43.61 b | 54.79 a | 19.01 d | 34.76 c | 2.741 | 0.01 |
| MDA (nmol/mg prot.) | 7.93 c | 5.67 e | 3.96 e | 6.77 cd | 18.97 a | 10.29 b | 0.545 | 0.01 |
| LDH (U/mL) | 831.41 b | 947.54 b | 1026.94 b | 851.80 b | 1428.91 a | 1038.04 b | 25.365 | 0.01 |
| SDH (U/mL) | 21.66 e | 28.00 cd | 33.19 c | 24.66 ed | 81.00 a | 50.00 b | 2.019 | 0.01 |
| GluDH (U/mL) | 34.00 a | 31.66 a | 30.64 a | 33.00 a | 22.33 b | 33.00 a | 3.483 | 0.01 |
| Liver | ||||||||
| TAOC (U/mg prot.) | 256.06 a | 264.57 a | 260.80 a | 240.18 b | 112.99 d | 198.10 c | 3.138 | 0.01 |
| SOD (U/mg prot.) | 433.74 b | 426.67 b | 430.34 b | 481.78 a | 322.12 d | 372.99 c | 5.739 | 0.01 |
| GSH (µmol/g prot.) | 5.60 b | 4.83 c | 4.64 bc | 5.18 b | 3.71 c | 4.75 a | 0.248 | 0.01 |
| GSH-Px activity (U) | 406.11 bc | 388.94 bc | 403.43 bc | 458.93 b | 263.44 c | 330.58 b | 46.540 | 0.01 |
| Catalase (U/mg prot.) | 49.83 b | 47.45 b | 48.28 b | 58.05 a | 20.15 d | 36.84 c | 1.007 | 0.01 |
| MDA (nmol/mg prot.) | 4.20 c | 3.00 d | 3.45 cd | 3.58 cd | 10.05 a | 5.45 b | 0.214 | 0.01 |
| LDH (U/mL) | 1276.76 c | 1304.43 c | 1300.63 c | 1206.56 c | 2597.06 a | 1633.50 b | 27.900 | 0.01 |
| SDH (U/mL) | 11.47 d | 14.83 c | 13.54 cd | 13.06 cd | 42.91 a | 26.49 b | 0.650 | 0.01 |
| GluDH (U/mL) | 180.13 a | 177.77 a | 172.52 a | 174.83 a | 138.65 c | 154.83 b | 5.684 | 0.01 |
| Kidney | ||||||||
| TAOC (U/mg prot.) | 107.93 a | 111.51 a | 109.92 a | 101.23 b | 47.62 d | 83.50 c | 1.322 | 0.01 |
| SOD (U/mg prot.) | 182.82 b | 179.84 b | 181.39 b | 203.07 a | 135.77 d | 157.21 c | 2.419 | 0.01 |
| GSH (µmol/g prot.) | 2.36 a | 1.61 c | 1.96 bc | 2.18 b | 1.56 c | 2.14 b | 0.104 | 0.01 |
| GSH-Px activity (U) | 171.17 b | 163.94 bc | 170.04 b | 196.80 a | 111.04 d | 146.52 c | 19.616 | 0.01 |
| Catalase (U/mg prot.) | 21.00 b | 20.00 b | 20.35 b | 24.47 a | 8.49 d | 15.52 c | 0.424 | 0.01 |
| MDA (nmol/mg prot.) | 1.77 c | 1.26 d | 1.45 cd | 1.51 cd | 4.23 a | 2.29 b | 0.090 | 0.01 |
| LDH (U/mL) | 1251.22 c | 1278.34 c | 1274.62 c | 1245.12 c | 2682.43 a | 1661.25 b | 27.341 | 0.01 |
| SDH (U/mL) | 4.83 d | 6.25 c | 5.71 cd | 5.29 cd | 18.08 a | 11.16 b | 0.274 | 0.01 |
| GluDH (U/mL) | 75.92 a | 70.71 a | 72.71 a | 73.69 a | 49.87 b | 73.96 a | 2.396 | 0.01 |
| Heart | ||||||||
| TAOC (U/mg prot.) | 133.39 a | 137.82 a | 135.85 a | 125.11 b | 58.86 d | 103.19 c | 1.635 | 0.01 |
| SOD (U/mg prot.) | 225.94 b | 222.26 b | 224.18 b | 250.97 a | 167.80 d | 194.30 c | 2.989 | 0.01 |
| GSH (µmol/g prot.) | 2.91 b | 1.99 c | 2.42 bc | 2.70 b | 1.93 c | 3.88 a | 0.129 | 0.01 |
| GSH-Px activity (U) | 211.55 bc | 202.61 bc | 210.16 bc | 227.23 a | 137.23 c | 188.37 b | 24.24 | 0.01 |
| Catalase (U/mg prot.) | 25.95 b | 24.71 b | 25.15 b | 28.51 a | 10.34 d | 19.19 c | 0.524 | 0.01 |
| MDA (nmol/mg prot.) | 2.18 c | 1.56 d | 1.80 cd | 1.86 cd | 5.23 a | 2.84 b | 0.111 | 0.01 |
| LDH (U/mL) | 1280.63 c | 1308.39 c | 1304.57 c | 1210.21 c | 2604.93 a | 1638.44 b | 27.98 | 0.01 |
| SDH (U/mL) | 5.98 d | 7.72 c | 7.05 cd | 6.80 cd | 22.35 a | 13.80 b | 0.339 | 0.01 |
| GluDH (U/mL) | 93.83 a | 87.39 a | 89.87 a | 91.07 a | 61.63 b | 91.07 a | 2.96 | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elwan, H.; Mohamed, A.S.A.; Dawood, D.H.; Elnesr, S.S. Modulatory Effects of Arctostaphylos uva-urs Extract In Ovo Injected into Broiler Embryos Contaminated by Aflatoxin B1. Animals 2022, 12, 2042. https://doi.org/10.3390/ani12162042
Elwan H, Mohamed ASA, Dawood DH, Elnesr SS. Modulatory Effects of Arctostaphylos uva-urs Extract In Ovo Injected into Broiler Embryos Contaminated by Aflatoxin B1. Animals. 2022; 12(16):2042. https://doi.org/10.3390/ani12162042
Chicago/Turabian StyleElwan, Hamada, Abdelhameed S. A. Mohamed, Dawood Hosni Dawood, and Shaaban S. Elnesr. 2022. "Modulatory Effects of Arctostaphylos uva-urs Extract In Ovo Injected into Broiler Embryos Contaminated by Aflatoxin B1" Animals 12, no. 16: 2042. https://doi.org/10.3390/ani12162042
APA StyleElwan, H., Mohamed, A. S. A., Dawood, D. H., & Elnesr, S. S. (2022). Modulatory Effects of Arctostaphylos uva-urs Extract In Ovo Injected into Broiler Embryos Contaminated by Aflatoxin B1. Animals, 12(16), 2042. https://doi.org/10.3390/ani12162042