
Ad Libitum Feeding in Broiler Breeder Hens Alters the Transcriptome of Granulosa Cells of Pre-Hierarchal Follicles


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Sample Collection
2.3. RNA Extraction
2.4. RNA Sequencing and Quality Control
2.5. RNA Sequencing Analysis
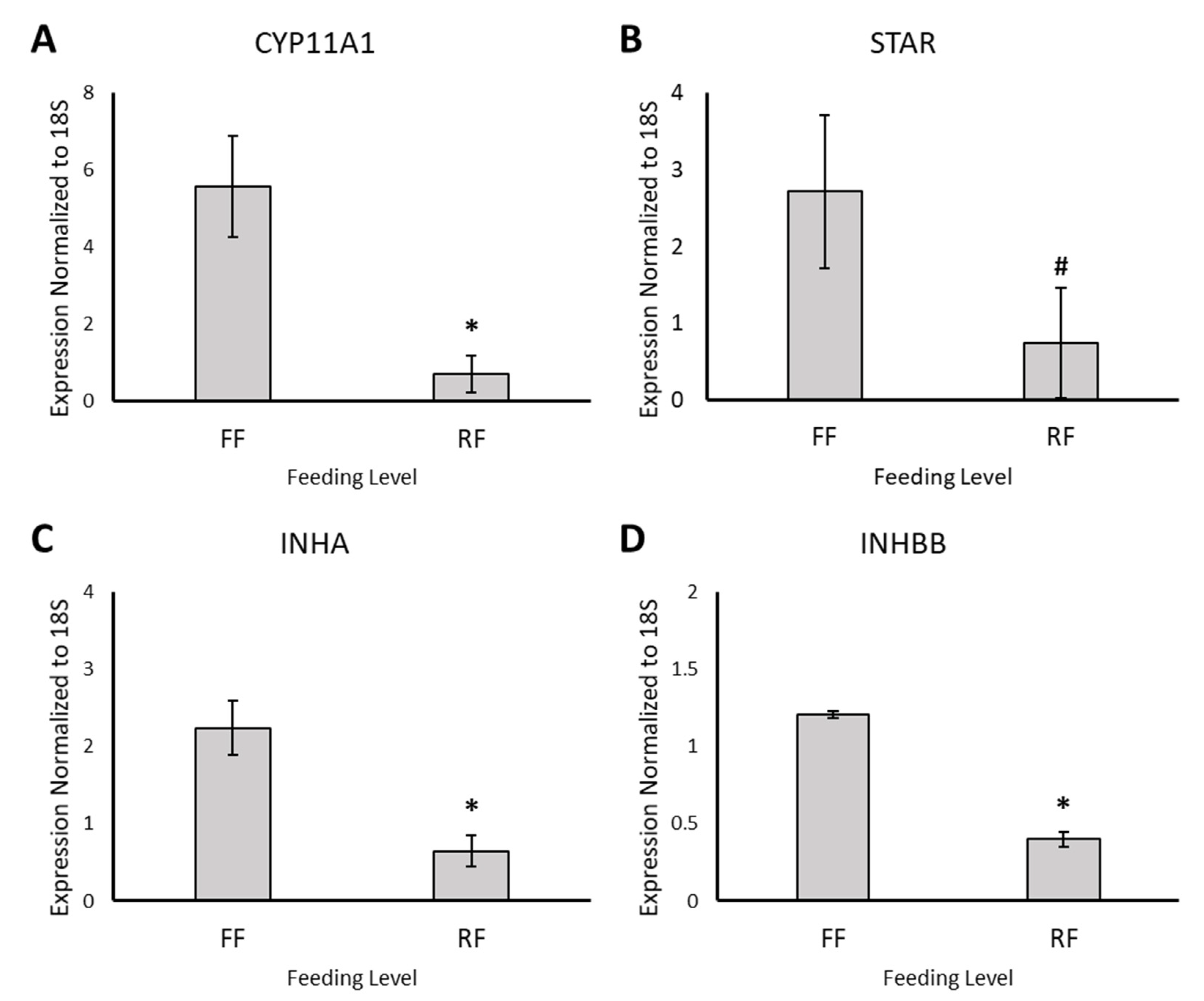
2.6. cDNA Synthesis and Real-Time qPCR
2.7. Statistical Analysis
3. Results
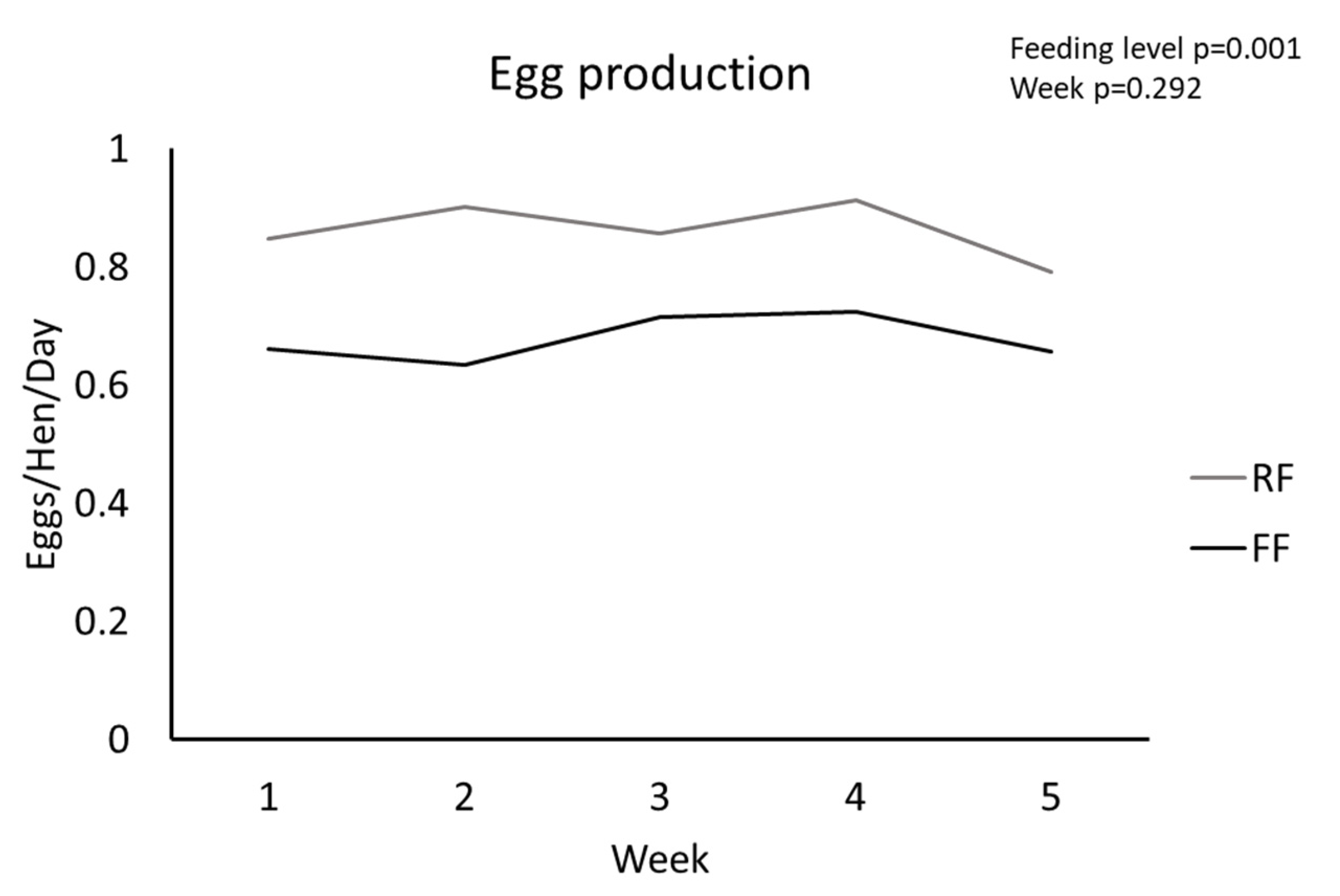
3.1. Body Parameters and Ovarian Morphology
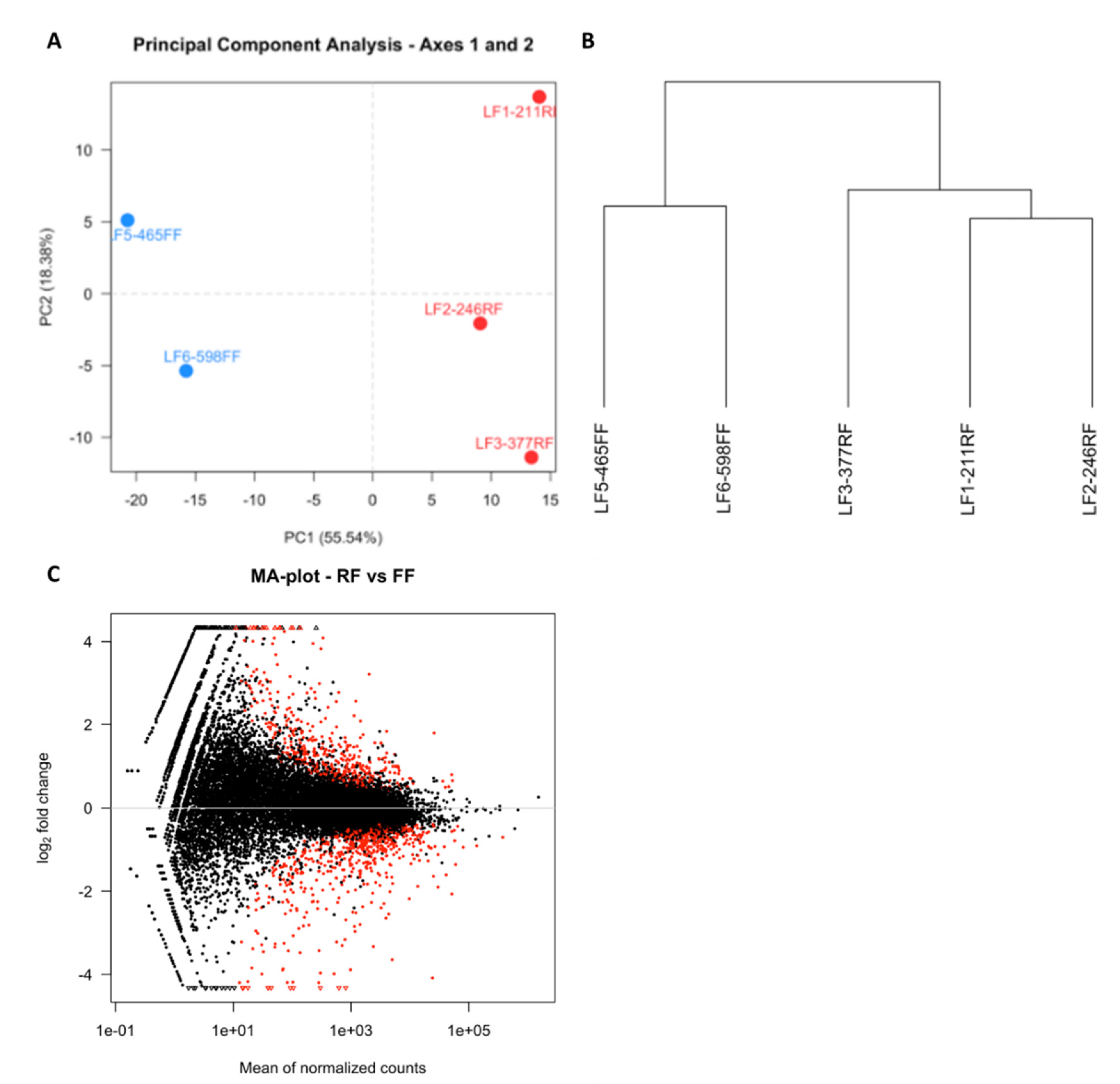
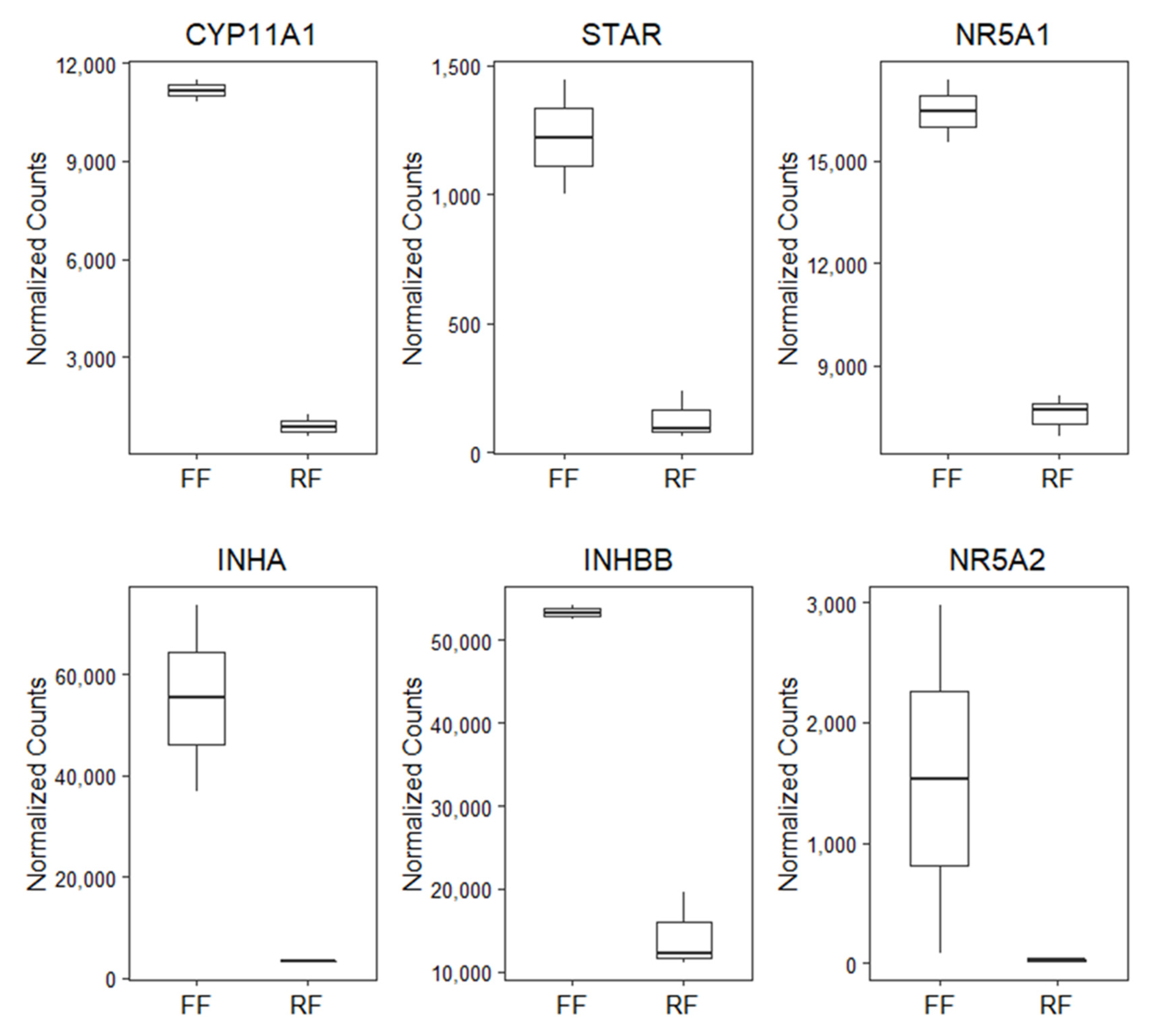
3.2. RNA Sequencing and Differential Gene Expression
3.3. Enrichment Analysis
3.4. Upstream Regulators
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu, M.W.; Robinson, F.E.; Charles, R.G.; Weingardt, R. Effect of feed allowance during rearing and breeding on female broiler breeders. 2. Ovarian morphology and production. Poult. Sci. 1992, 71, 1750–1761. [Google Scholar] [CrossRef] [PubMed]
- Renema, R.A.; Robinson, F.E.; Proudman, J.A.; Newcombe, M.; Mckay, R.I. Effects of Body Weight and Feed Allocation During Sexual Maturation in Broiler Breeder Hens. 2. Ovarian Morphology and Plasma Hormone Profiles. Poult. Sci. 1999, 78, 629–639. [Google Scholar] [CrossRef]
- Hocking, P.M.; Gilbert, A.B.; Walker, M.; Waddington, D. Ovarian follicular structure of White Leghorns fed ad libitum and dwarf and normal broiler breeders fed ad libitum or restricted until point of lay. Br. Poult. Sci. 1987, 28, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P. Follicle Selection in the Avian Ovary. Reprod. Domest. Anim. 2012, 47, 283–287. [Google Scholar] [CrossRef]
- Robinson, F.E.; Etches, R.J. Ovarian Steroidogenesis During Foillicular Maturation in the Domestic Fowl (Gallus Domesticus). Biol. Reprod. 1986, 35, 1096–1105. [Google Scholar] [CrossRef]
- Johnson, A.L.; Bridgham, J.T. Regulation of Steroidogenic Acute Regulatory Protein and Luteinizing Hormone Receptor Messenger Ribonucleic Acid in Hen Granulosa Cells. Endocrinology 2001, 142, 3116–3124. [Google Scholar] [CrossRef] [PubMed]
- Woods, D.C.; Johnson, A.L. Regulation of Follicle-Stimulating Hormone-Receptor Messenger RNA in Hen Granulosa Cells Relative to Follicle Selection. Biol. Reprod. 2005, 72, 643–650. [Google Scholar] [CrossRef]
- Johnson, A.; Woods, D.C. Ovarian dynamics and follicle development. In Reproductive Biology and Phylogeny of Birds; Jamieson, B.G.M., Ed.; Science Publishers, Inc.: Enfield, NH, USA, 2007; pp. 243–277. [Google Scholar]
- Li, Z.; Johnson, A.L. Regulation of P450 Cholesterol Side-Chain Cleavage Messenger Ribonucleic Acid Expression and Progesterone Production in Hen Granulosa Cells. Biol. Reprod. 1993, 49, 463–469. [Google Scholar] [CrossRef]
- Tilly, J.L.; Kowalski, K.I.; Johnson, A.L. Stage of Ovarian Follicular Development Associated with the Initiation of Steroidogenic Competence in Avian Granulosa Cells. Biol. Reprod. 1991, 44, 305–314. [Google Scholar] [CrossRef]
- Yu, M.W.; Robinson, F.E.; Etches, R.J. Quantification of ovarian steroidogenesis in the domestic fowl by incubation of intact large follicles. Poult. Sci. 1992, 71, 346–351. [Google Scholar] [CrossRef]
- Johnson, P.A.; Johnson, A.L.; van Tienhoven, A. Evidence for a positive feedback interaction between progesterone and luteinizing hormone in the induction of ovulation in the hen, Gallus domesticus. Gen. Comp. Endocrinol. 1985, 58, 478–485. [Google Scholar] [CrossRef]
- Cobb-Vantress, Cobb 700 Breeder Management Supplement. 2013.
- Stephens, C.S.; Johnson, P.A. Occludin expression and regulation in small follicles of the layer and broiler breeder hen. Gen. Comp. Endocrinol. 2017, 248, 106–113. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J.; Ying Wang, C.; Yan Kwok, A.H.; Leung, F.C. Epidermal Growth Factor (EGF) Receptor Ligands in the Chicken Ovary: I. Evidence for Heparin-Binding EGF-Like Growth Factor (HB-EGF) as a Potential Oocyte-Derived Signal to Control Granulosa Cell Proliferation and HB-EGF and Kit Ligand Expression. Endocrinology 2007, 148, 3426–3440. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef] [PubMed]
- Ingenuity Pathway Analysis. Available online: https://www.qiagenbioinformatics.com/products/ingenuitypathway-analysis (accessed on 19 January 2021).
- Krämer, A.; Green, J.; Pollard, J.; Tugendreich, S. Causal analysis approaches in ingenuity pathway analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.L.; Lee, J. Granulosa cell responsiveness to follicle stimulating hormone during early growth of hen ovarian follicles. Poult. Sci. 2016, 95, 108–114. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Stephens, C.S.; Hill-Ricciuti, A.; Francoeur, L.; Johnson, P.A. Feeding Level Is Associated With Altered Liver Transcriptome and Follicle Selection in the Hen. Biol. Reprod. 2021. in review. [Google Scholar]
- Ciccone, N.A.; Dunn, I.C.; Sharp, P.J. Increased food intake stimulates GnRH-I, glycoprotein hormone α-subunit and follistatin mRNAs, and ovarian follicular numbers in laying broiler breeder hens. Domest. Anim. Endocrinol. 2007, 33, 62–76. [Google Scholar] [CrossRef]
- Johnson, A.L.; Woods, D.C. Dynamics of avian ovarian follicle development: Cellular mechanisms of granulosa cell differentiation. Gen. Comp. Endocrinol. 2009, 163, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Fang, C.; Li, J.; Mo, C.; Wang, Y.; Li, J. Transcriptomic Diversification of Granulosa Cells during Follicular Development in Chicken. Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef]
- Saxena, D.; Escamilla-Hernandez, R.; Little-Ihrig, L.; Zeleznik, A.J. Liver Receptor Homolog-1 and Steroidogenic Factor-1 Have Similar Actions on Rat Granulosa Cell Steroidogenesis. Endocrinology 2007, 148, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Havelock, J.C.; Carr, B.R.; Attia, G.R. The Orphan Nuclear Receptor, Liver Receptor Homolog-1, Regulates Cholesterol Side-Chain Cleavage Cytochrome P450 Enzyme in Human Granulosa Cells. J. Clin. Endocrinol. Metab. 2005, 90, 1678–1685. [Google Scholar] [CrossRef]
- Saxena, D.; Safi, R.; Little-Ihrig, L.; Zeleznik, A.J. Liver receptor homolog-1 stimulates the progesterone biosynthetic pathway during follicle-stimulating hormone-induced granulosa cell differentiation. Endocrinology 2004, 145, 3821–3829. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.W.; Robinson, F.E.; Etches, R.J. Effect of feed allowance during rearing and breeding on female broiler breeders. 3. Ovarian steroidogenesis. Poult. Sci. 1992, 71, 1762–1767. [Google Scholar] [CrossRef] [PubMed]
- Hui, D.Y.; Howles, P.N. Carboxyl ester lipase: Structure-function relationship and physiological role in lipoprotein metabolism and atherosclerosis. J. Lipid Res. 2002, 43, 2017–2030. [Google Scholar] [CrossRef]
- Sato, K.; Seol, H.S.; Kamada, T. Tissue distribution of lipase genes related to triglyceride metabolism in laying hens (Gallus gallus). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2010, 155, 62–66. [Google Scholar] [CrossRef]
- Brodt-Eppley, J.; Hui, D.Y. Dietary regulation of cholesterol esterase mRNA level in rat pancreas. J. Lipid Res. 1994, 35, 27–35. [Google Scholar] [CrossRef]
- Favara, D.M.; Zois, C.E.; Haider, S.; Pires, E.; Sheldon, H.; McCullagh, J.; Banham, A.H.; Harris, A.L. ADGRL4/ELTD1 silencing in endothelial cells induces ACLY and SLC25A1 and alters the cellular metabolic profile. Metabolites 2019, 9, 287. [Google Scholar] [CrossRef]
- Bruggeman, V.; D′Hondt, E.; Berghman, L.; Onagbesan, O.; Vanmontfort, D.; Vandesande, F.; Decuypere, E. The effect of food intake from 2 to 24 weeks of age on LHRH-I content in the median eminence and gonadotrophin levels in pituitary and plasma in female broiler breeder chickens. Gen. Comp. Endocrinol. 1998, 112, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Hadinia, S.H.; Carneiro, P.R.O.; Fitzsimmons, C.J.; Bédécarrats, G.Y.; Zuidhof, M.J. Post-photostimulation energy intake accelerated pubertal development in broiler breeder pullets. Poult. Sci. 2020, 99, 2215–2229. [Google Scholar] [CrossRef]
- Onagbesan, O.M.; Metayer, S.; Tona, K.; Williams, J.; Decuypere, E.; Bruggeman, V. Effects of genotype and feed allowance on plasma luteinizing hormones, follicle-stimulating hormones, progesterone, estradiol levels, follicle differentiation, and egg production rates of broiler breeder hens. Poult. Sci. 2006, 85, 1245–1258. [Google Scholar] [CrossRef] [PubMed]
- Tosca, L.; Chabrolle, C.; Crochet, S.; Tesseraud, S.; Dupont, J. IGF-1 receptor signaling pathways and effects of AMPK activation on IGF-1-induced progesterone secretion in hen granulosa cells. Domest. Anim. Endocrinol. 2008, 34, 204–216. [Google Scholar] [CrossRef]
- Ahumada-Solórzano, S.M.; Martínez-Moreno, C.G.; Carranza, M.; Ávila-Mendoza, J.; Luna-Acosta, J.L.; Harvey, S.; Luna, M.; Arámburo, C. Autocrine/paracrine proliferative effect of ovarian GH and IGF-I in chicken granulosa cell cultures. Gen. Comp. Endocrinol. 2016, 234, 47–56. [Google Scholar] [CrossRef]
- Winters, T.A.; Hanten, J.A.; Veldhuis, J.D. In situ amplification of the cytochrome P-450 cholesterol side-chain cleavage enzyme mRNA in single porcine granulosa cells by IGF-1 and FSH acting alone or in concert. Endocrine 1998, 9, 57–63. [Google Scholar] [CrossRef]
- Adashi, E.Y.; Resnick, C.E.; Brodie, A.M.H.; Svoboda, M.E.; Van Wyk, J.J. Somatomedin-C-mediated potentiation of follicle-stimulating hormone-induced aromatase activity of cultured rat granulosa cells. Endocrinology 1985, 117, 2313–2320. [Google Scholar] [CrossRef] [PubMed]
- Eimerl, S.; Orly, J. Regulation of steroidogenic genes by insulin-like growth factor-1 and follicle-stimulating hormone: Differential responses of cytochrome P450 side-chain cleavage. Biol. Reprod. 2002, 67, 900–910. [Google Scholar] [CrossRef]
- Balasubramanian, K.; LaVoie, H.A.; Garmey, J.C.; Stocco, D.M.; Veldhuis, J.D. Regulation of Porcine Granulosa Cell Steroidogenic Acute Regulatory Protein (StAR) by Insulin-Like Growth Factor I: Synergism with Follicle-Stimulating Hormone or Protein Kinase A Agonist. Endocrinology 1997, 138, 433–439. [Google Scholar] [CrossRef][Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence |
|---|---|
| CYP11A1 | F 5′-ACTTCAAGGGACTGAGCTTTGGGT-3′ R 5′ AGTTCTCCAGGATGTGCATGAGGA 3′ |
| STAR | F 5′-TGCCTGAGCAGCAGGGATTTATCA- 3′ R 5′- TGGTTGATGATGGTCTTTGGCAGC-3′ |
| INHA | F 5′-TCTTCCCTTCCACAGACGTG- 3′ R 5′- CTGTAGAACCAGAGCTGGGC-3′ |
| INHBB | F 5′-TTCGCCGAGACAGACGAT- 3′ R 5′- TTACTTTTCGCCTGCTGCCT-3′ |
| Parameter | RF | FF | p-Value |
|---|---|---|---|
| Body weight (g) | 3389 ± 162 | 4483 ± 359 | <0.0001 |
| Fat pad weight/BW | 0.015 ± 0.007 | 0.030 ± 0.008 | <0.0001 |
| Liver weight/BW | 0.028 ± 0.005 | 0.048 ± 0.010 | <0.0001 |
| Ovary weight/BW | 0.018 ± 0.003 | 0.021 ± 0.005 | 0.101 |
| 3–5-mm follicles | 32.2 ± 11.0 | 36.3 ± 12.0 | 0.332 |
| 6–8-mm follicles | 11.6 ± 3.9 | 10.7 ± 3.7 | 0.519 |
| Follicles > 9-mm | 6.3 ± 0.9 | 8.2 ± 2.0 | 0.002 |
| Eggs/hen/day | 0.862 ± 0.048 | 0.678 ± 0.039 | 0.013 |
| GeneID | Gene Name | log2(FC) | FDR | FF Average | RF Average |
|---|---|---|---|---|---|
| ENSGALG00000034436 | CEL | −8.6 | 0.01 | 749 | 2 |
| ENSGALG00000002182 | NR5A2 | −6.4 | <0.01 | 1531 | 18 |
| ENSGALG00000038884 | SRL | −4.9 | <0.01 | 250 | 8 |
| ENSGALG00000045327 | - | −4.9 | <0.01 | 222 | 8 |
| ENSGALG00000026808 | TMEM72 | −4.8 | <0.01 | 1922 | 70 |
| ENSGALG00000006440 | - | −4.2 | <0.01 | 652 | 35 |
| ENSGALG00000050830 | SV2A | −4.2 | <0.01 | 237 | 13 |
| ENSGALG00000054770 | INHA | −4.1 | <0.01 | 55,417 | 3271 |
| ENSGALG00000010269 | KCNAB1 | −4.0 | <0.01 | 1174 | 75 |
| ENSGALG00000042836 | KCNH2 | −4.0 | <0.01 | 688 | 44 |
| ENSGALG00000047771 | PGF | −3.9 | <0.01 | 2220 | 150 |
| ENSGALG00000034982 | CYP11A1 | −3.6 | <0.01 | 11,165 | 893 |
| ENSGALG00000029968 | GADD45B | −3.5 | <0.01 | 300 | 27 |
| ENSGALG00000001207 | PLCH2 | −3.4 | 0.01 | 630 | 59 |
| ENSGALG00000006598 | SORL1 | −3.3 | <0.01 | 5205 | 517 |
| ENSGALG00000041932 | SLC5A5 | −3.3 | <0.01 | 1534 | 155 |
| ENSGALG00000003242 | STAR | −3.2 | <0.01 | 1222 | 132 |
| ENSGALG00000010364 | AADAC | −3.2 | 0.01 | 1694 | 189 |
| ENSGALG00000040355 | TCF24 | −3.1 | <0.01 | 220 | 26 |
| ENSGALG00000004974 | PPARG | −3.0 | <0.01 | 2454 | 314 |
| ENSGALG00000005884 | MAPKKK3L | −2.9 | <0.01 | 270 | 37 |
| ENSGALG00000011242 | OBSL1 | −2.8 | <0.01 | 2359 | 328 |
| ENSGALG00000042607 | RSPO3 | −2.8 | <0.01 | 321 | 45 |
| ENSGALG00000014938 | ABHD3 | −2.8 | <0.01 | 396 | 56 |
| ENSGALG00000038399 | PLEKHA6 | −2.8 | <0.01 | 2921 | 420 |
| ENSGALG00000050611 | - | −2.7 | <0.01 | 1650 | 257 |
| ENSGALG00000041143 | UMOD | −2.6 | <0.01 | 7916 | 1266 |
| ENSGALG00000011537 | PDE10A | −2.6 | <0.01 | 325 | 53 |
| ENSGALG00000008815 | LRRN4 | −2.5 | <0.01 | 8678 | 1484 |
| ENSGALG00000011608 | INF2 | −2.5 | <0.01 | 2394 | 418 |
| ENSGALG00000011803 | EMP1 | −2.5 | <0.01 | 347 | 61 |
| ENSGALG00000019077 | - | −2.5 | <0.01 | 2109 | 372 |
| ENSGALG00000043234 | HBA1 | −2.5 | <0.01 | 236 | 43 |
| ENSGALG00000042929 | - | −2.4 | <0.01 | 746 | 139 |
| ENSGALG00000010326 | FLVCR2 | −2.4 | <0.01 | 530 | 98 |
| ENSGALG00000006453 | TF | −2.4 | <0.01 | 232 | 43 |
| ENSGALG00000037603 | SESN2 | −2.4 | <0.01 | 6298 | 1195 |
| ENSGALG00000003750 | PLCG1 | −2.4 | <0.01 | 3056 | 590 |
| ENSGALG00000033683 | PHOSPHO1 | −2.3 | <0.01 | 6924 | 1408 |
| ENSGALG00000016415 | MAP7D2 | −2.3 | <0.01 | 4248 | 882 |
| ENSGALG00000044649 | P2RX2 | −2.3 | <0.01 | 1134 | 235 |
| ENSGALG00000016954 | RGCC | −2.3 | <0.01 | 416 | 87 |
| ENSGALG00000008537 | EPHB3 | −2.3 | <0.01 | 516 | 108 |
| ENSGALG00000036728 | PRDM16 | −2.2 | <0.01 | 436 | 92 |
| ENSGALG00000041708 | WNT4 | −2.2 | <0.01 | 18,324 | 4040 |
| ENSGALG00000014734 | - | −2.2 | <0.01 | 1166 | 260 |
| ENSGALG00000018803 | - | −2.1 | <0.01 | 415 | 98 |
| ENSGALG00000009512 | TFPI2 | −2.1 | <0.01 | 94,942 | 22,674 |
| ENSGALG00000049157 | DOK4 | −2.0 | <0.01 | 4274 | 1034 |
| ENSGALG00000038458 | LOXL1 | −2.0 | <0.01 | 1624 | 401 |
| GeneID | Gene Name | log2(FC) | FDR | FF Average | RF Average |
|---|---|---|---|---|---|
| ENSGALG00000051980 | - | 6.5 | 0.01 | 2 | 229 |
| ENSGALG00000036798 | COL4A1 | 4.1 | 0.02 | 32 | 535 |
| ENSGALG00000007819 | PDZRN3 | 4.0 | <0.01 | 24 | 361 |
| ENSGALG00000015908 | COL12A1 | 3.8 | <0.01 | 34 | 491 |
| ENSGALG00000043754 | GLUL | 3.2 | <0.01 | 340 | 3158 |
| ENSGALG00000010858 | LRP2 | 3.0 | 0.03 | 56 | 446 |
| ENSGALG00000040755 | ANGPT4 | 2.9 | <0.01 | 44 | 320 |
| ENSGALG00000048104 | - | 2.8 | <0.01 | 54 | 367 |
| ENSGALG00000003670 | MAFB | 2.8 | <0.01 | 137 | 938 |
| ENSGALG00000016843 | COL4A2 | 2.8 | 0.01 | 40 | 271 |
| ENSGALG00000033338 | GPT2 | 2.6 | <0.01 | 60 | 374 |
| ENSGALG00000030065 | TENM3 | 2.6 | <0.01 | 306 | 1805 |
| ENSGALG00000009405 | GRIA2 | 2.6 | <0.01 | 143 | 839 |
| ENSGALG00000034453 | SAMD11 | 2.4 | <0.01 | 113 | 604 |
| ENSGALG00000050840 | APCDD1 | 2.2 | <0.01 | 355 | 1641 |
| ENSGALG00000006172 | ABCC8 | 2.1 | 0.01 | 120 | 534 |
| ENSGALG00000038364 | NOV | 2.1 | <0.01 | 492 | 2083 |
| ENSGALG00000016820 | GAS6 | 2.1 | <0.01 | 49 | 207 |
| ENSGALG00000013697 | CNDP1 | 2.1 | <0.01 | 106 | 443 |
| ENSGALG00000011200 | THBS2 | 2.0 | <0.01 | 65 | 264 |
| ENSGALG00000014178 | - | 2.0 | <0.01 | 82 | 318 |
| ENSGALG00000011623 | ADAMTS3 | 1.9 | <0.01 | 208 | 803 |
| ENSGALG00000031916 | ZP1 | 1.8 | 0.01 | 78 | 283 |
| ENSGALG00000000667 | EDN2 | 1.8 | <0.01 | 1580 | 5639 |
| ENSGALG00000026055 | PALM | 1.8 | <0.01 | 60 | 210 |
| ENSGALG00000009687 | KCNK2 | 1.8 | <0.01 | 10,362 | 35,972 |
| ENSGALG00000004812 | FAM129A | 1.8 | 0.02 | 58 | 203 |
| ENSGALG00000007268 | - | 1.8 | <0.01 | 192 | 669 |
| ENSGALG00000002671 | - | 1.8 | <0.01 | 660 | 2230 |
| ENSGALG00000032836 | - | 1.7 | <0.01 | 137 | 445 |
| ENSGALG00000012595 | AGTPBP1 | 1.7 | <0.01 | 96 | 310 |
| ENSGALG00000001768 | TENM2 | 1.7 | 0.01 | 340 | 1095 |
| ENSGALG00000011994 | SYNPO2 | 1.7 | 0.02 | 66 | 215 |
| ENSGALG00000036883 | MET | 1.7 | <0.01 | 619 | 1988 |
| ENSGALG00000009612 | TGFB2 | 1.7 | <0.01 | 188 | 600 |
| ENSGALG00000045776 | CPN2 | 1.7 | 0.01 | 90 | 282 |
| ENSGALG00000002081 | MMP28 | 1.6 | <0.01 | 112 | 348 |
| ENSGALG00000015542 | PLPPR1 | 1.6 | <0.01 | 105 | 324 |
| ENSGALG00000017065 | - | 1.6 | 0.05 | 84 | 259 |
| ENSGALG00000011145 | TRIL | 1.6 | 0.01 | 99 | 301 |
| ENSGALG00000034085 | - | 1.6 | 0.01 | 207 | 627 |
| ENSGALG00000031534 | ARID5B | 1.6 | <0.01 | 360 | 1081 |
| ENSGALG00000026981 | NHSL1 | 1.6 | 0.01 | 370 | 1102 |
| ENSGALG00000027514 | - | 1.6 | <0.01 | 1168 | 3445 |
| ENSGALG00000016251 | - | 1.6 | <0.01 | 1485 | 4369 |
| ENSGALG00000012115 | WT1 | 1.5 | <0.01 | 1514 | 4420 |
| ENSGALG00000010902 | CERS6 | 1.5 | 0.05 | 153 | 448 |
| ENSGALG00000051001 | - | 1.5 | 0.01 | 294 | 844 |
| ENSGALG00000009495 | FGFR2 | 1.5 | <0.01 | 229 | 648 |
| ENSGALG00000012834 | AKR1D1 | 1.5 | <0.01 | 460 | 1297 |
| Functional Category | Genes | Total Genes | FDR |
|---|---|---|---|
| Lipid catabolic process | 10 | 186 | 2.17 × 10−4 |
| Heme export | 2 | 2 | 1.65 × 10−2 |
| Positive regulation of actin filament bundle assembly | 4 | 44 | 3.44 × 10−2 |
| Cellular lipid catabolic process | 6 | 126 | 3.44 × 10−2 |
| Proteoglycan biosynthetic process | 4 | 48 | 3.81 × 10−2 |
| Inositol trisphosphate biosynthetic process | 3 | 21 | 3.81 × 10−2 |
| Positive regulation of cellular component biogenesis | 9 | 345 | 3.81 × 10−2 |
| Heme transport | 2 | 6 | 4.43 × 10−2 |
| Inositol trisphosphate metabolic process | 3 | 24 | 4.43 × 10−2 |
| Transmembrane transport | 17 | 1122 | 4.43 × 10−2 |
| Functional Category | Genes | Total Genes | FDR |
|---|---|---|---|
| Ion transport | 12 | 1117 | 3.26 × 10−2 |
| Cation transport | 10 | 757 | 3.26 × 10−2 |
| Muscle contraction | 5 | 171 | 3.26 × 10−2 |
| Nitric oxide mediated signal transduction | 2 | 11 | 3.26 × 10−2 |
| Regulation of heart contraction | 4 | 112 | 3.26 × 10−2 |
| Regulation of nitric oxide mediated signal transduction | 2 | 6 | 3.26 × 10−2 |
| Metal ion transport | 8 | 545 | 3.26 × 10−2 |
| Ion transmembrane transport | 10 | 790 | 3.26 × 10−2 |
| Regulation of system process | 6 | 291 | 3.26 × 10−2 |
| Positive regulation of cell size | 2 | 6 | 3.26 × 10−2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Francoeur, L.; Stephens, C.S.; Johnson, P.A. Ad Libitum Feeding in Broiler Breeder Hens Alters the Transcriptome of Granulosa Cells of Pre-Hierarchal Follicles. Animals 2021, 11, 2706. https://doi.org/10.3390/ani11092706
Francoeur L, Stephens CS, Johnson PA. Ad Libitum Feeding in Broiler Breeder Hens Alters the Transcriptome of Granulosa Cells of Pre-Hierarchal Follicles. Animals. 2021; 11(9):2706. https://doi.org/10.3390/ani11092706
Chicago/Turabian StyleFrancoeur, Laurie, Claire S. Stephens, and Patricia A. Johnson. 2021. "Ad Libitum Feeding in Broiler Breeder Hens Alters the Transcriptome of Granulosa Cells of Pre-Hierarchal Follicles" Animals 11, no. 9: 2706. https://doi.org/10.3390/ani11092706
APA StyleFrancoeur, L., Stephens, C. S., & Johnson, P. A. (2021). Ad Libitum Feeding in Broiler Breeder Hens Alters the Transcriptome of Granulosa Cells of Pre-Hierarchal Follicles. Animals, 11(9), 2706. https://doi.org/10.3390/ani11092706