
Effect of High Sulfur Diet on Rumen Fermentation, Microflora, and Epithelial Barrier Function in Steers


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
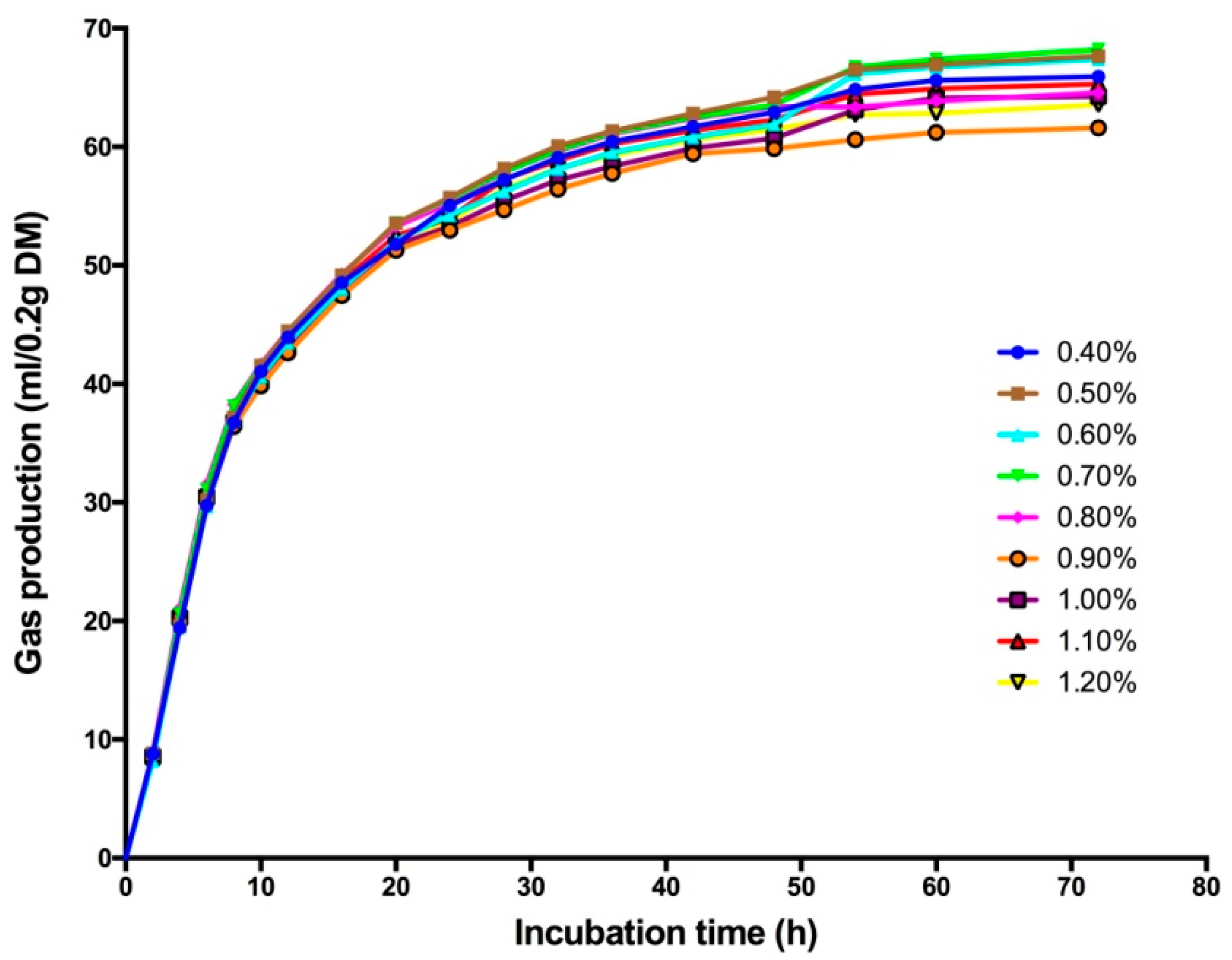
2.1. In Vitro Incubation Procedure
2.2. Animals, Diets, and Experimental Design
2.3. Sample Collection and Measurement
2.3.1. Collection of Rumen Fluid
2.3.2. Collection of Rumen Epithelial Tissue Samples
2.4. DNA Extraction, High-Throughput Sequencing, and Data Processing
2.5. Pretreatment of the Rumen Epithelial Tissue Section for Staining and Electron Microscopy
2.6. Measurement of Relative Expression of RNA
2.6.1. RNA Isolation and cDNA Synthesis
2.6.2. Primer Design and qRT-PCR
2.7. Statistical Analysis
3. Results
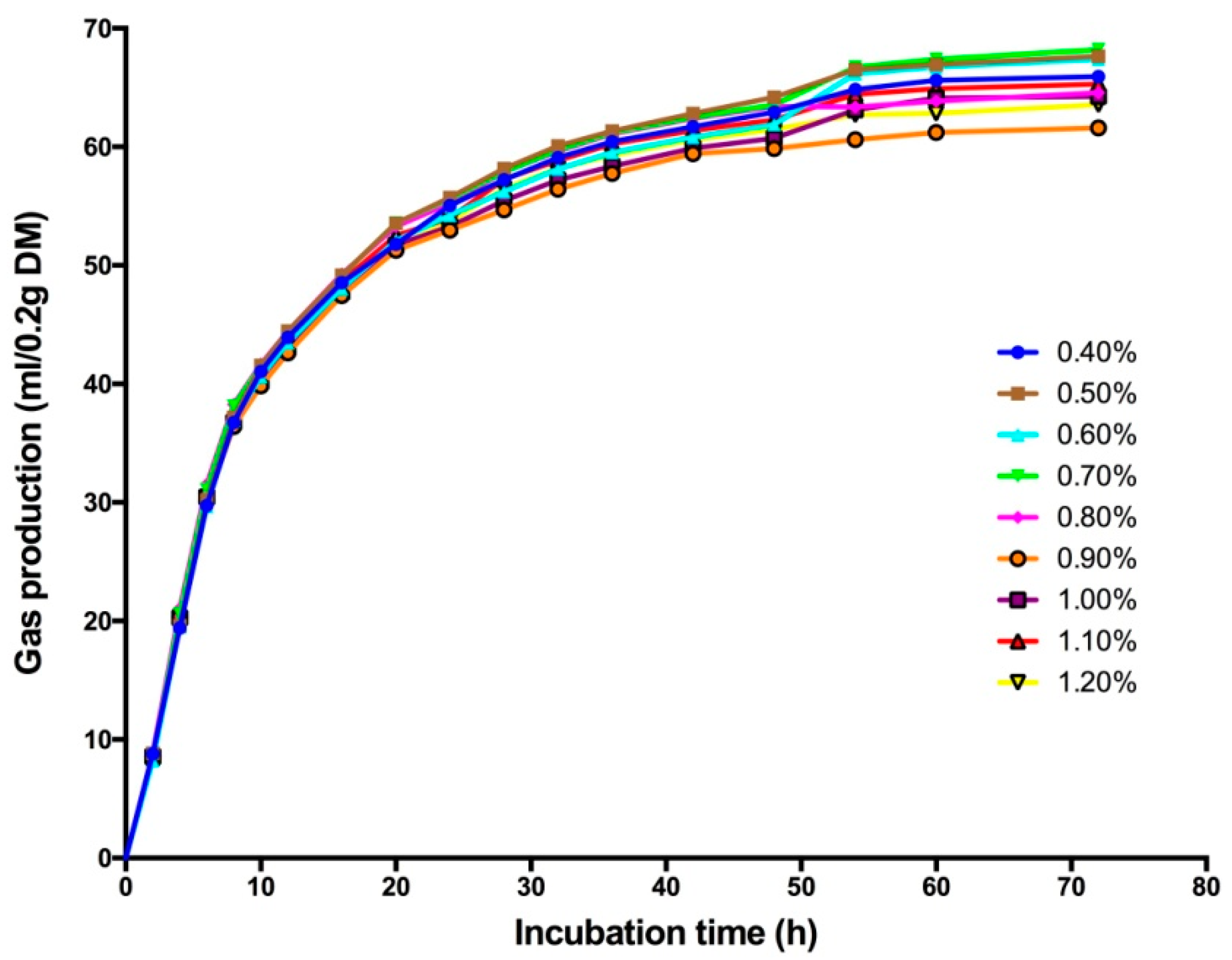
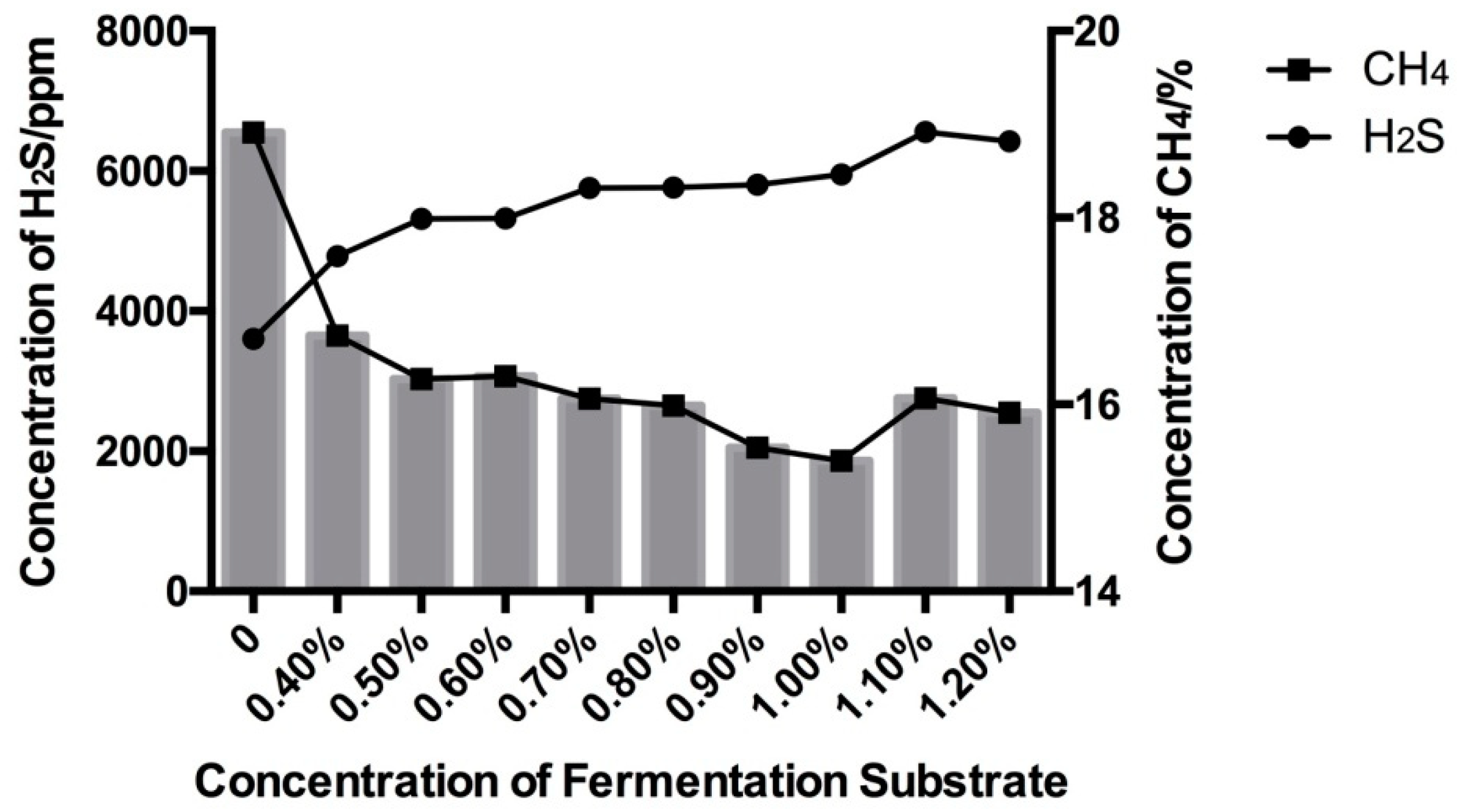
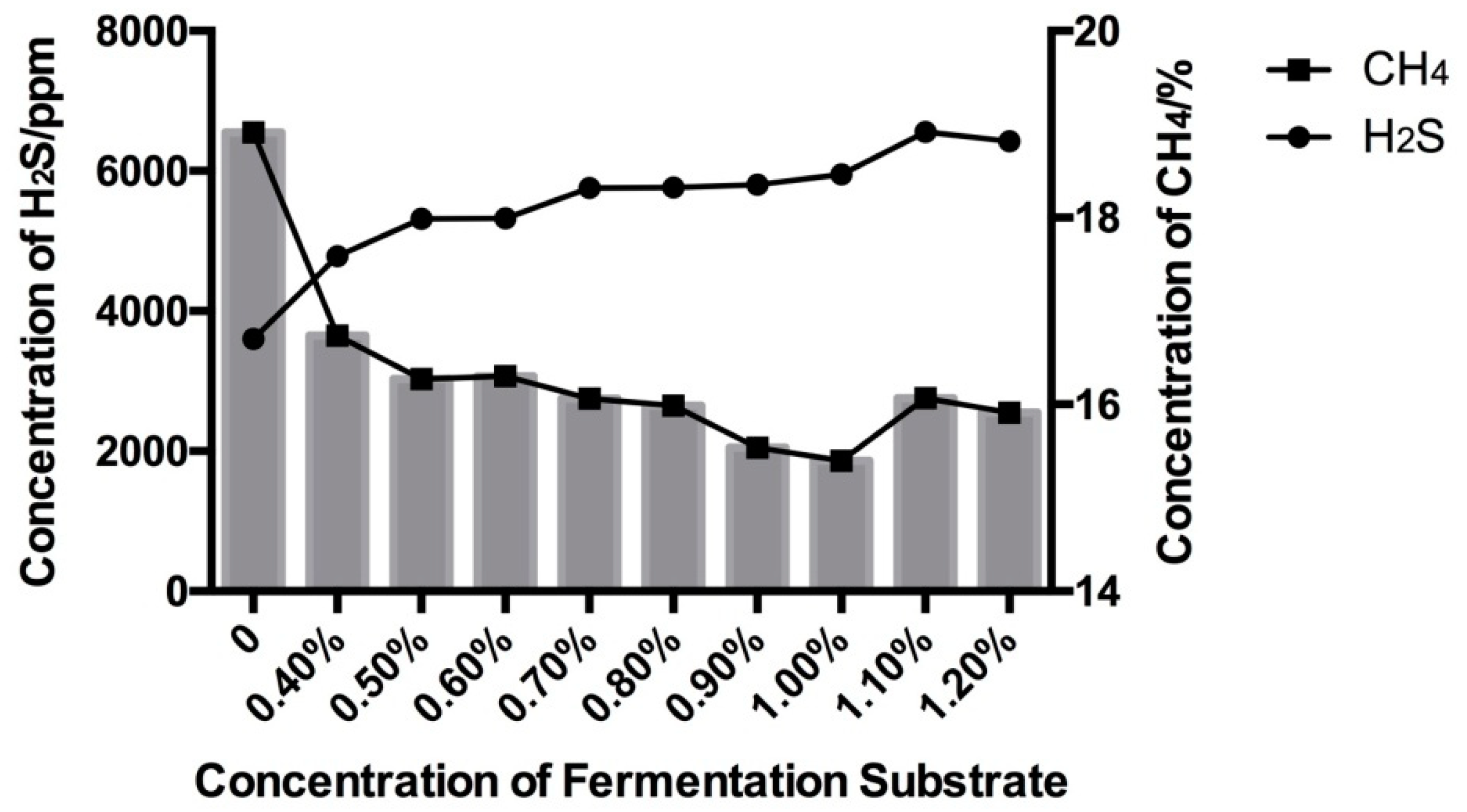
3.1. Effect of High Sulfur Diet on Rumen Fermentation
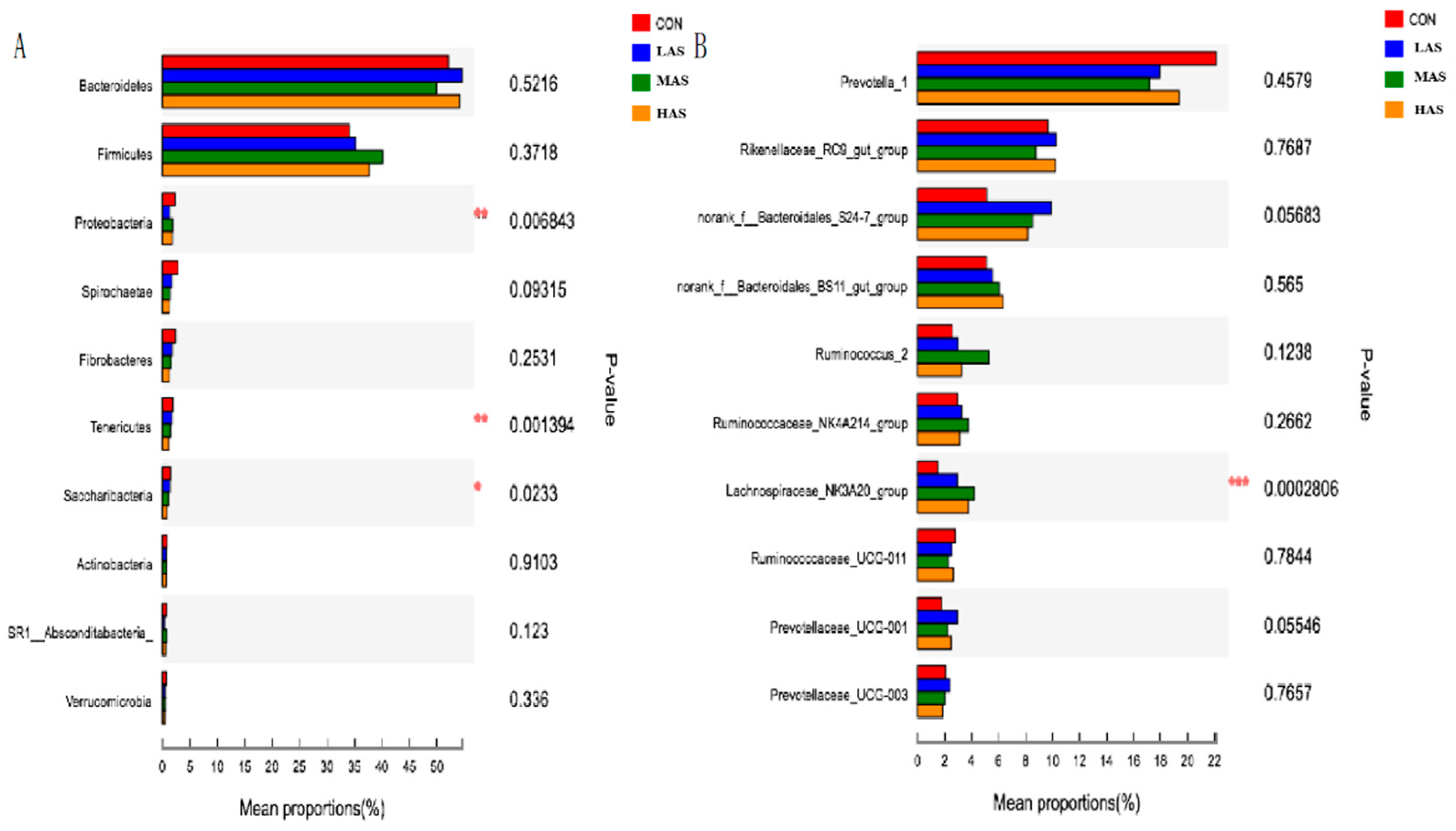
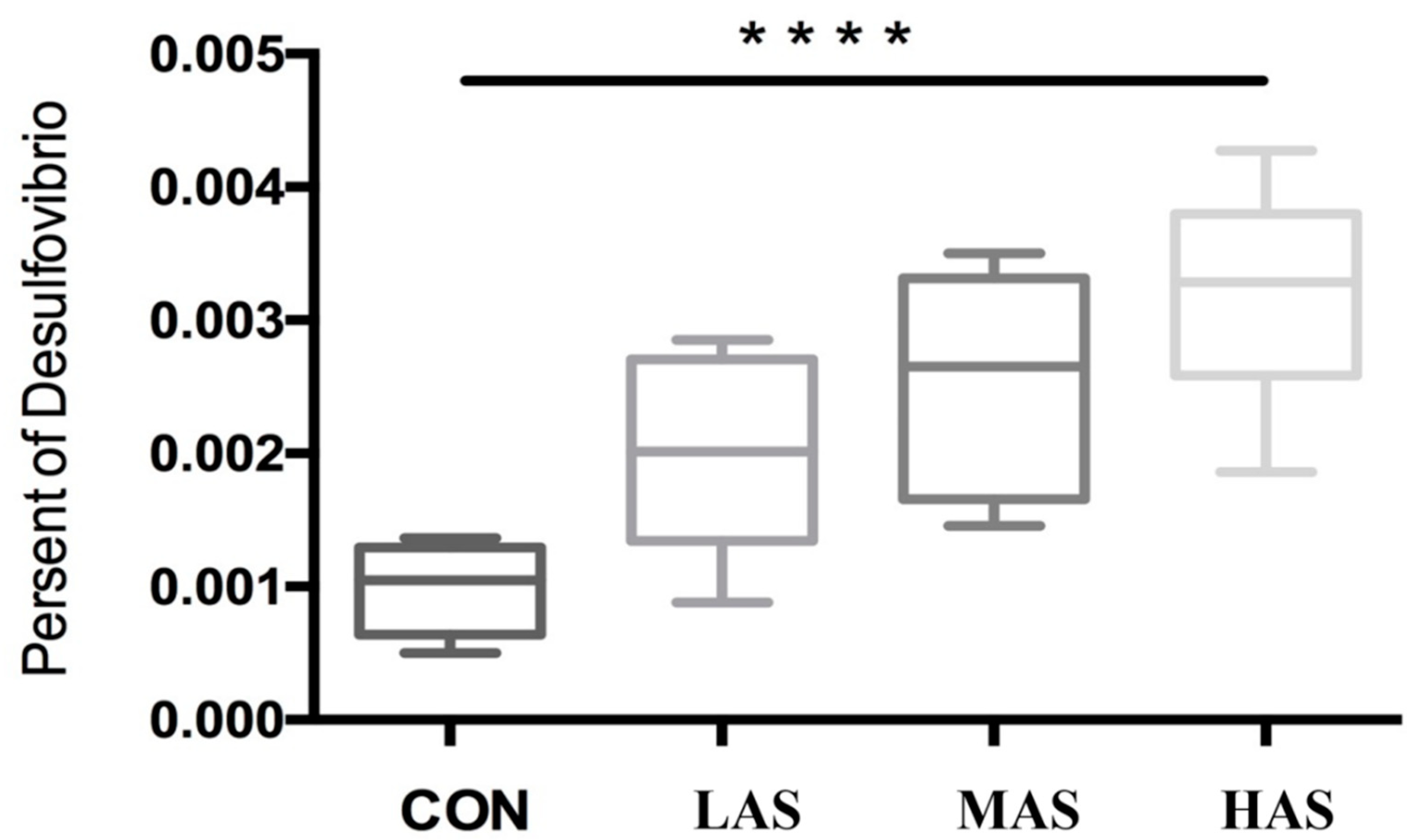
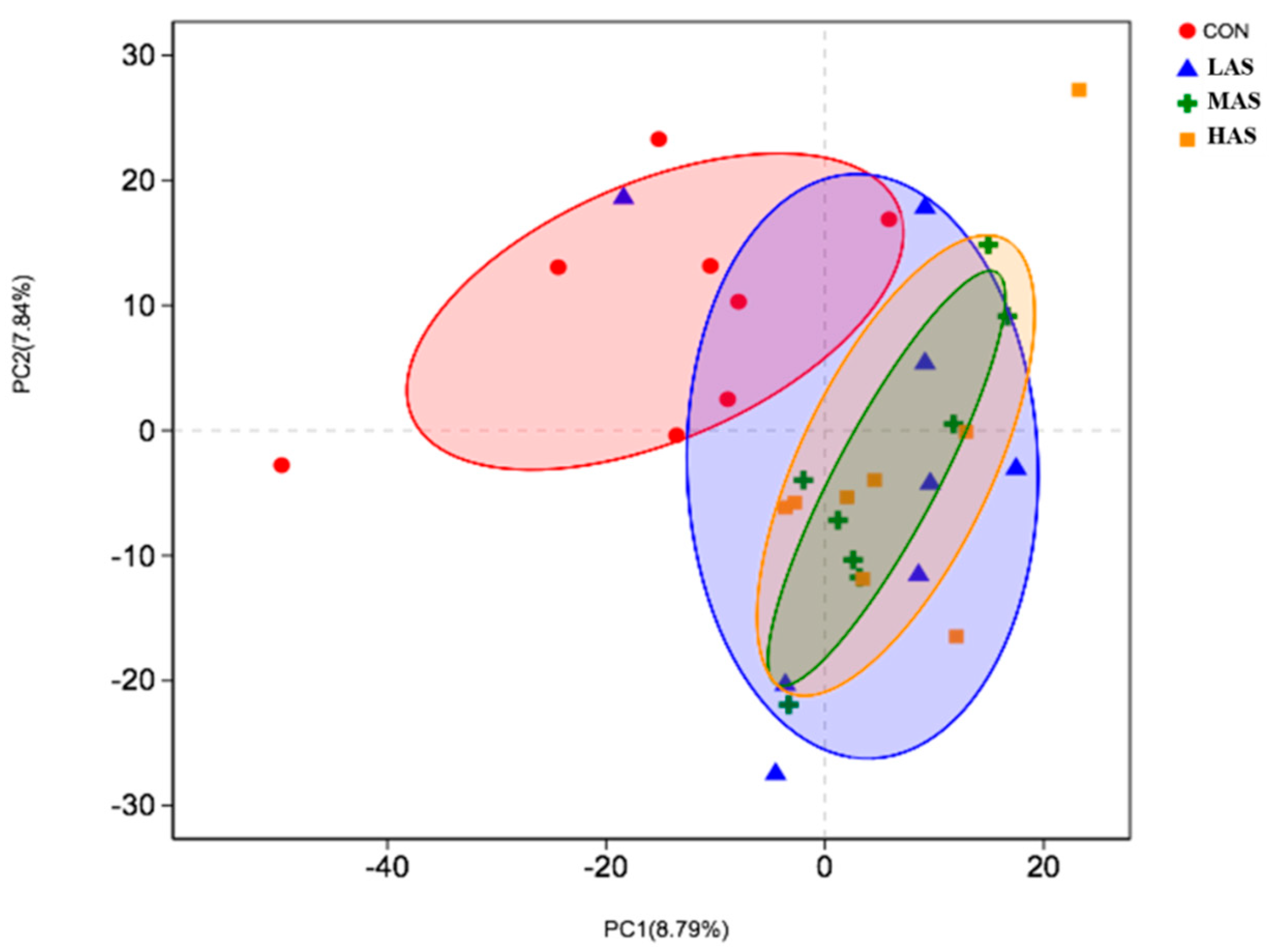
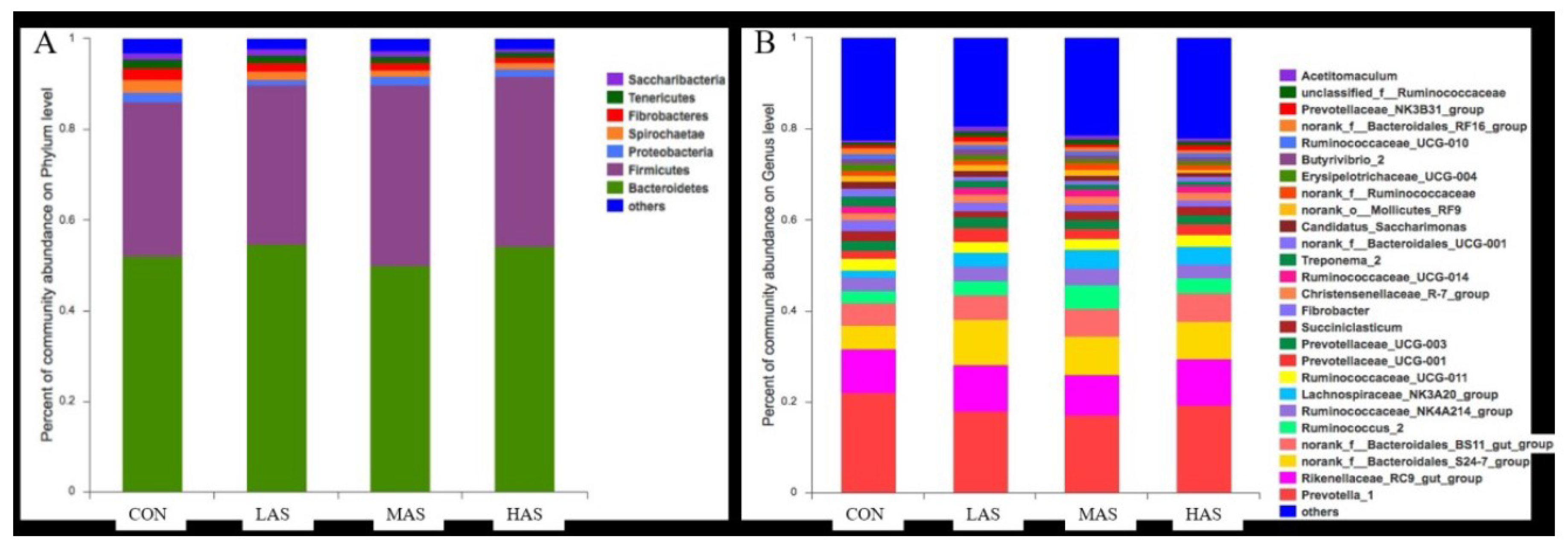
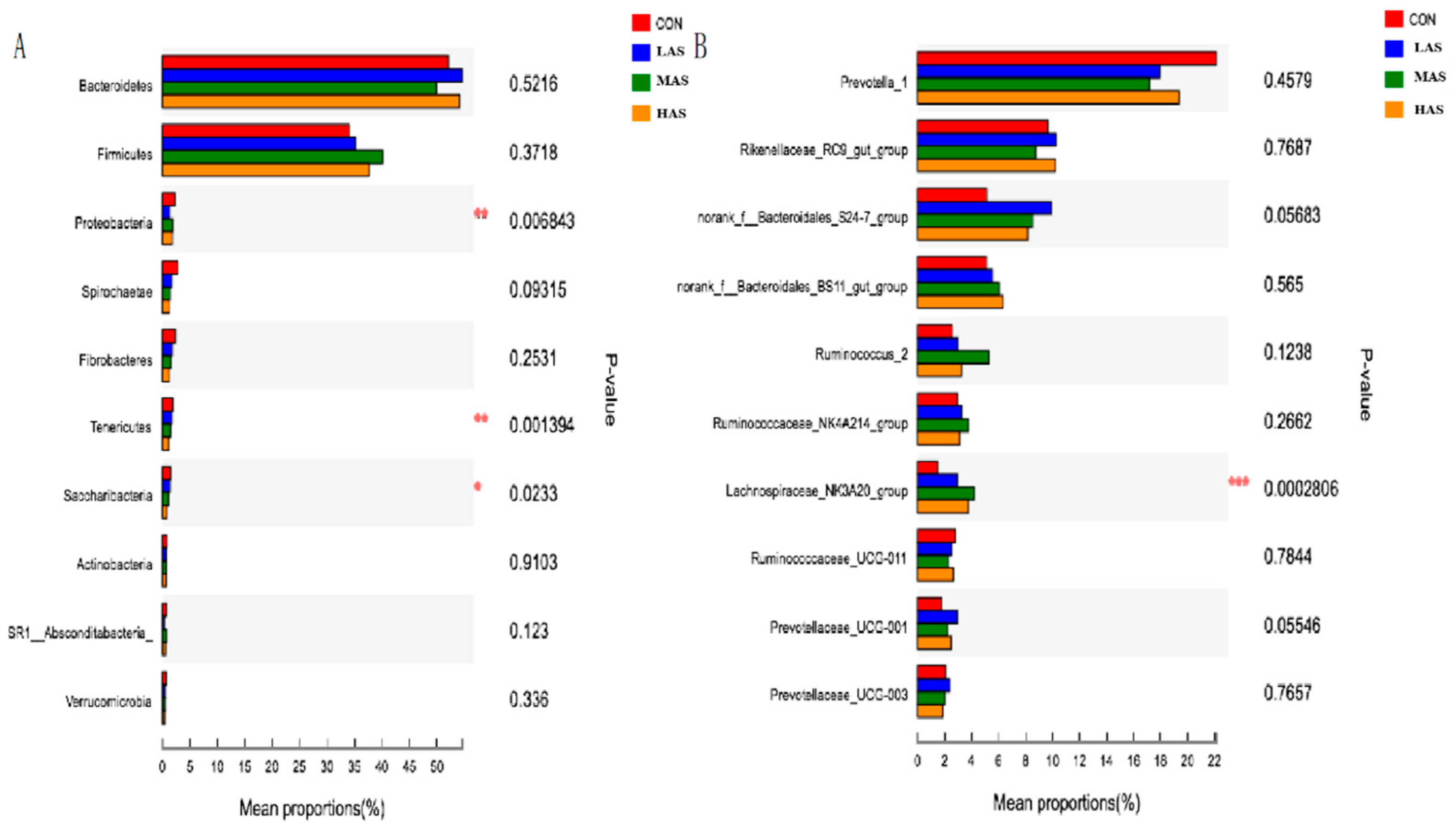
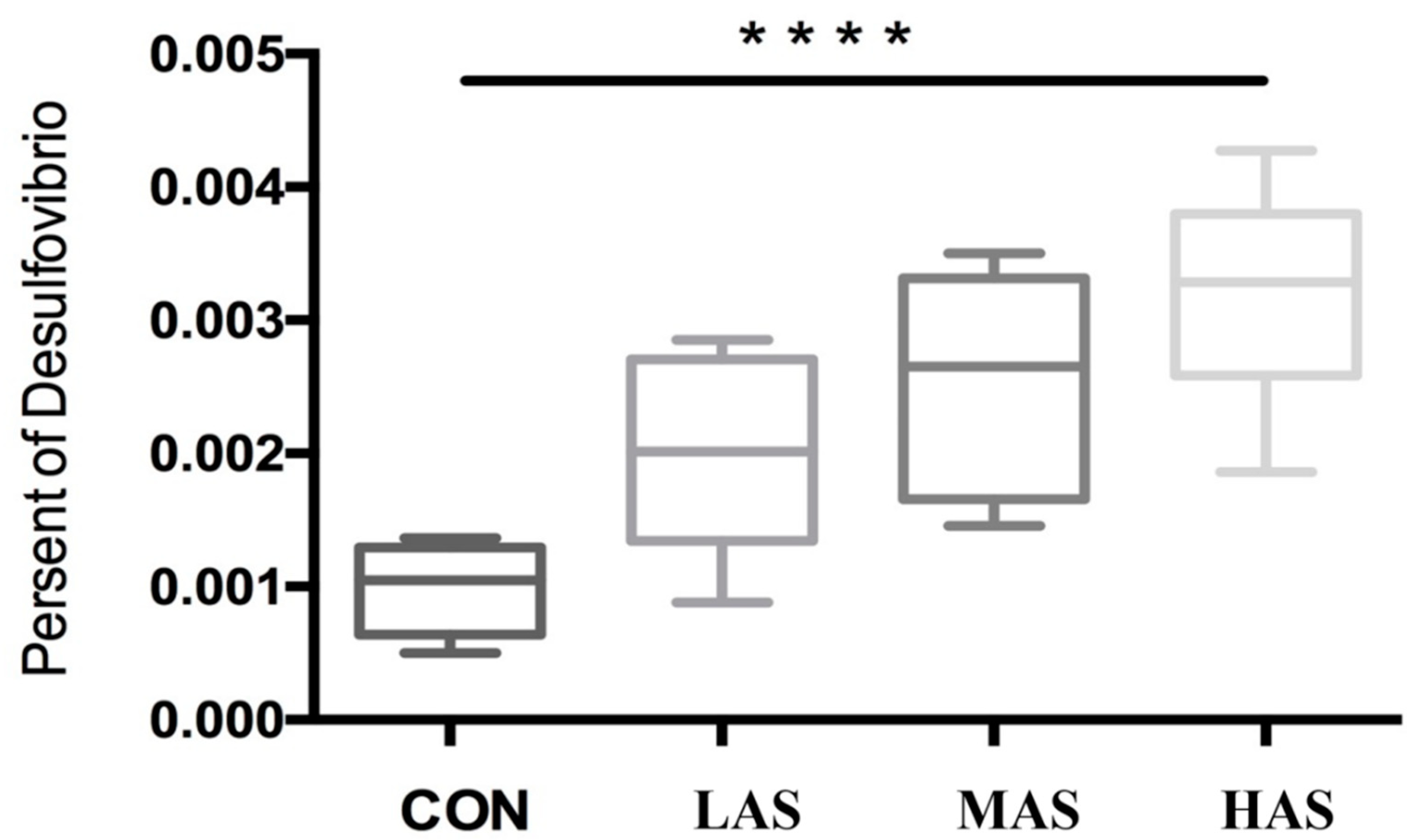
3.2. Effect of High Sulfur Diet on Rumen Microflora
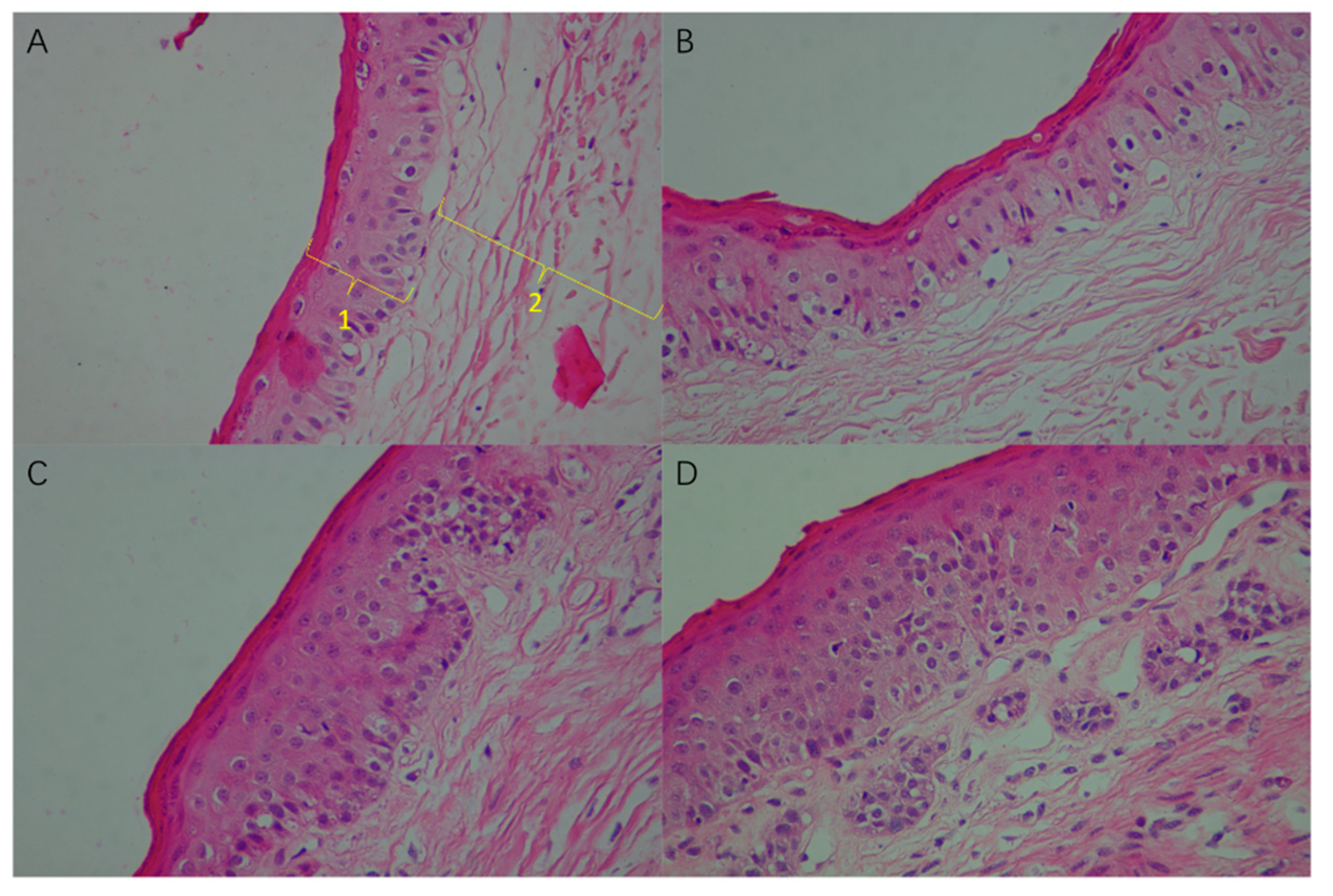
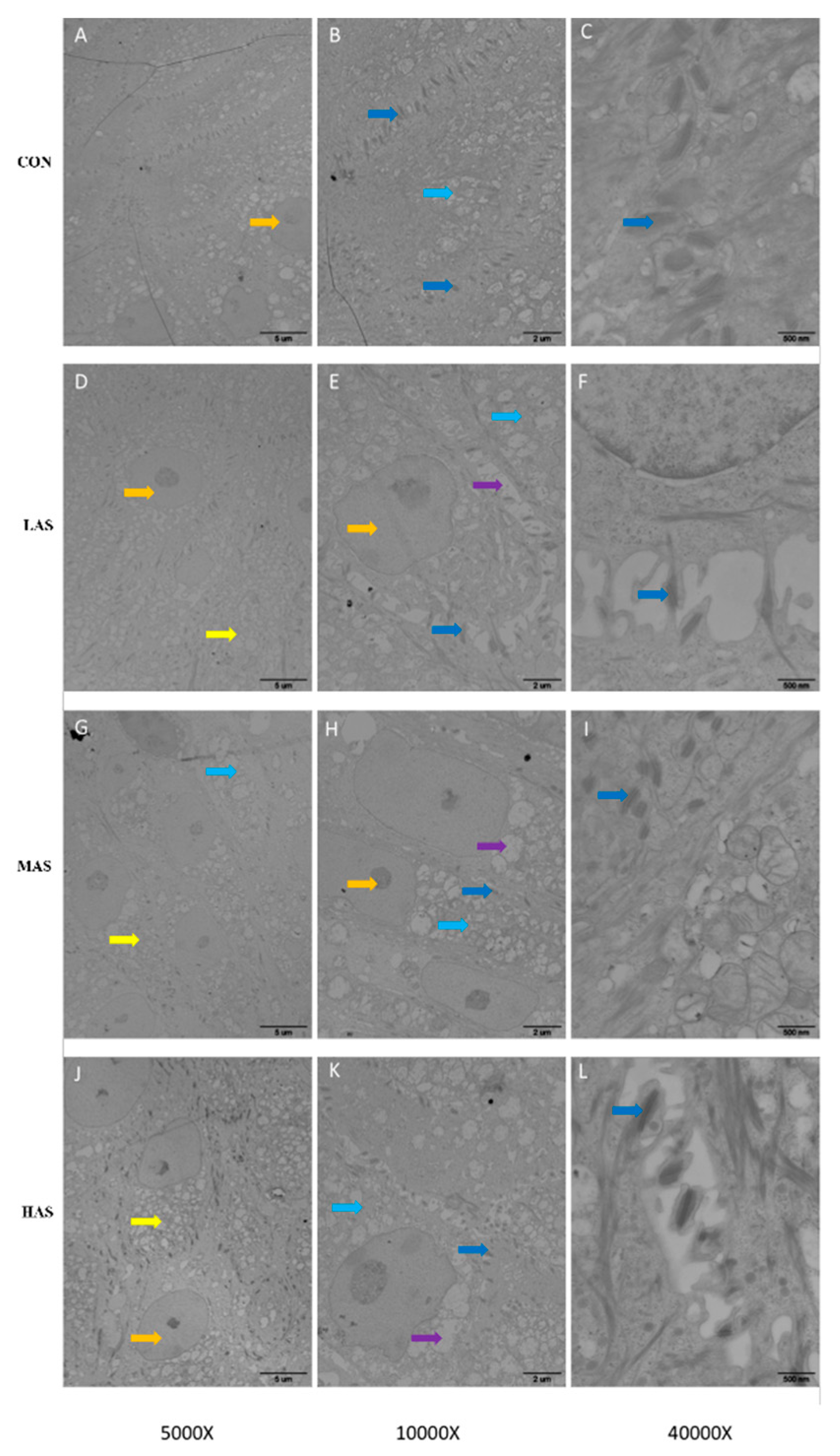
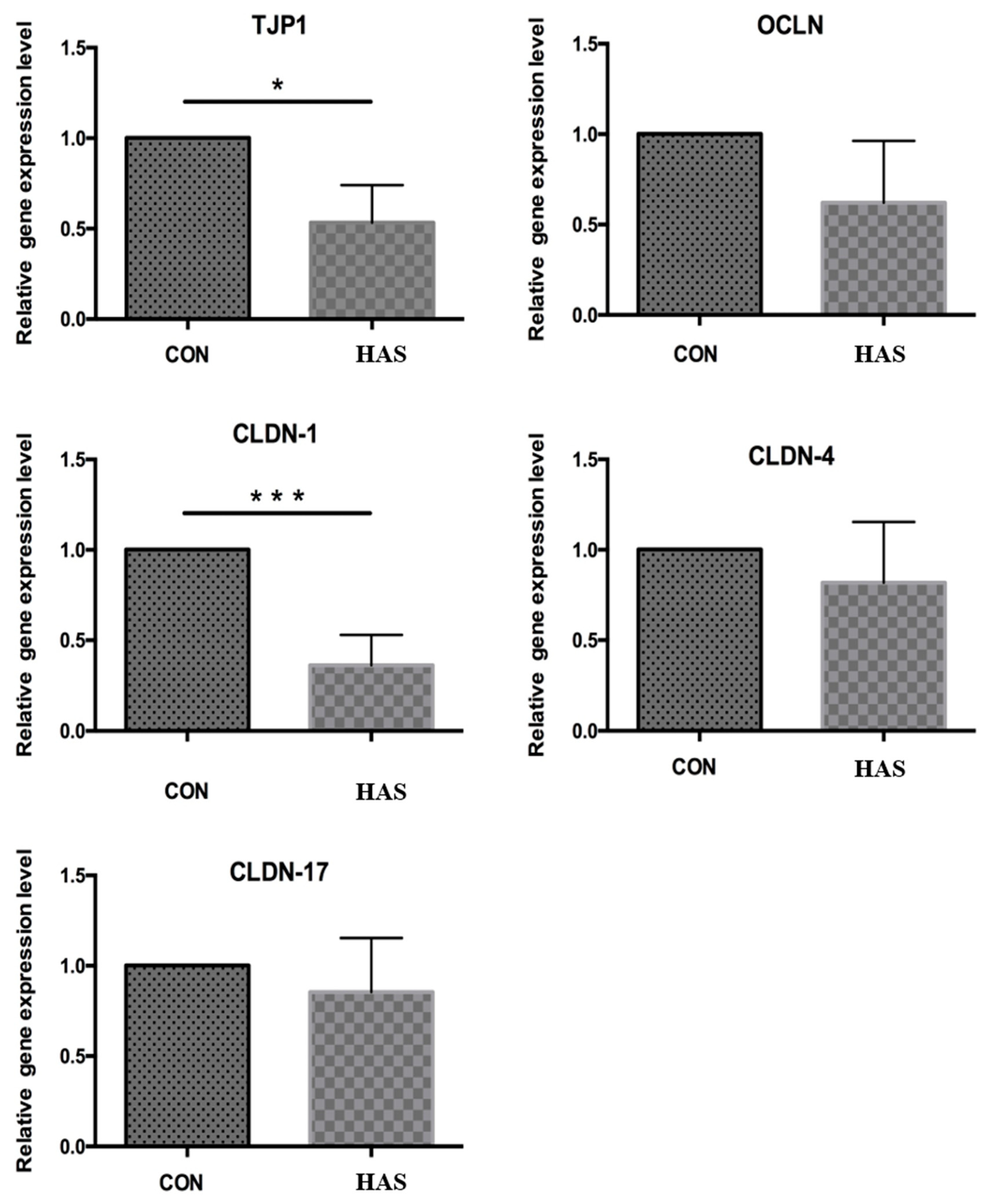
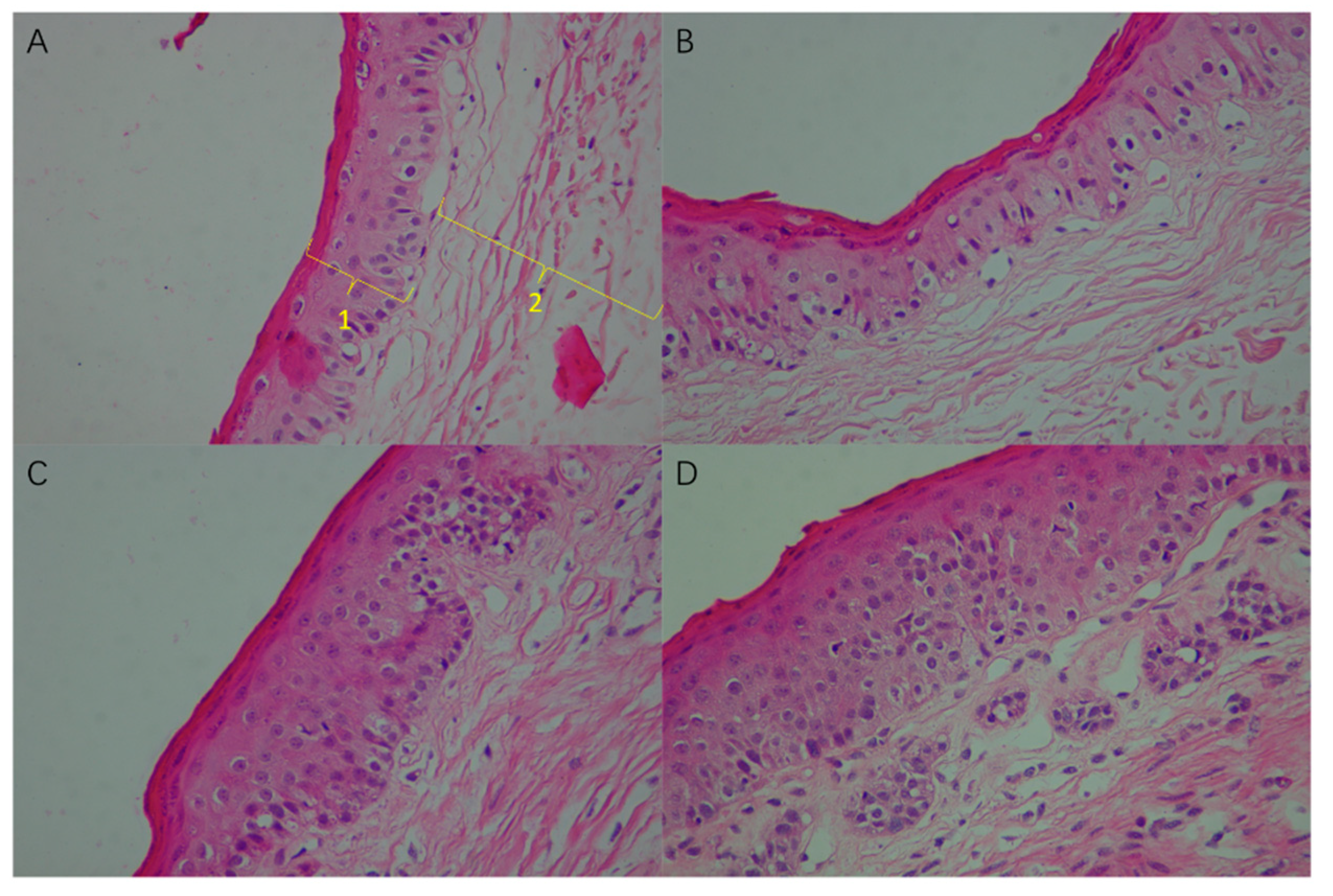
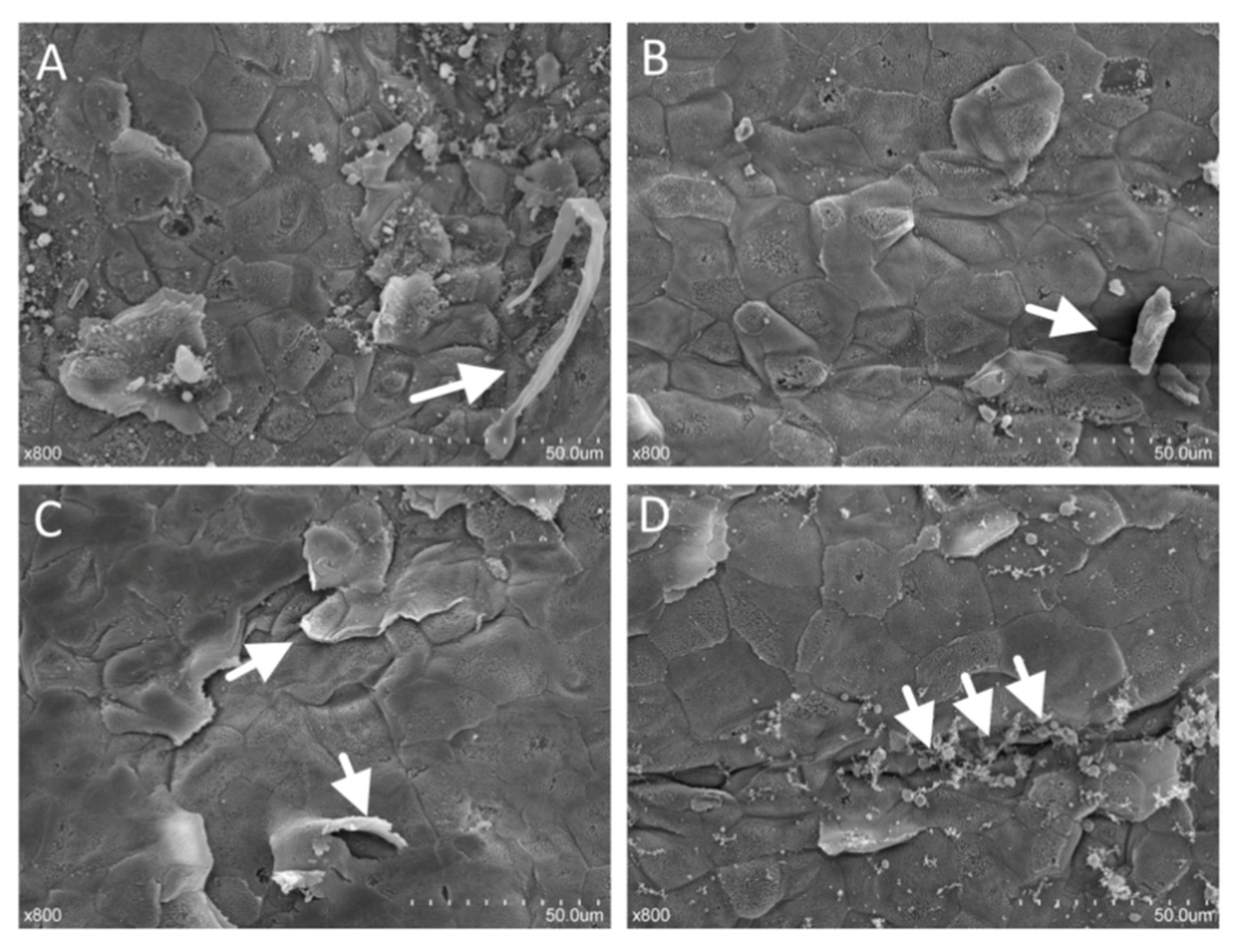
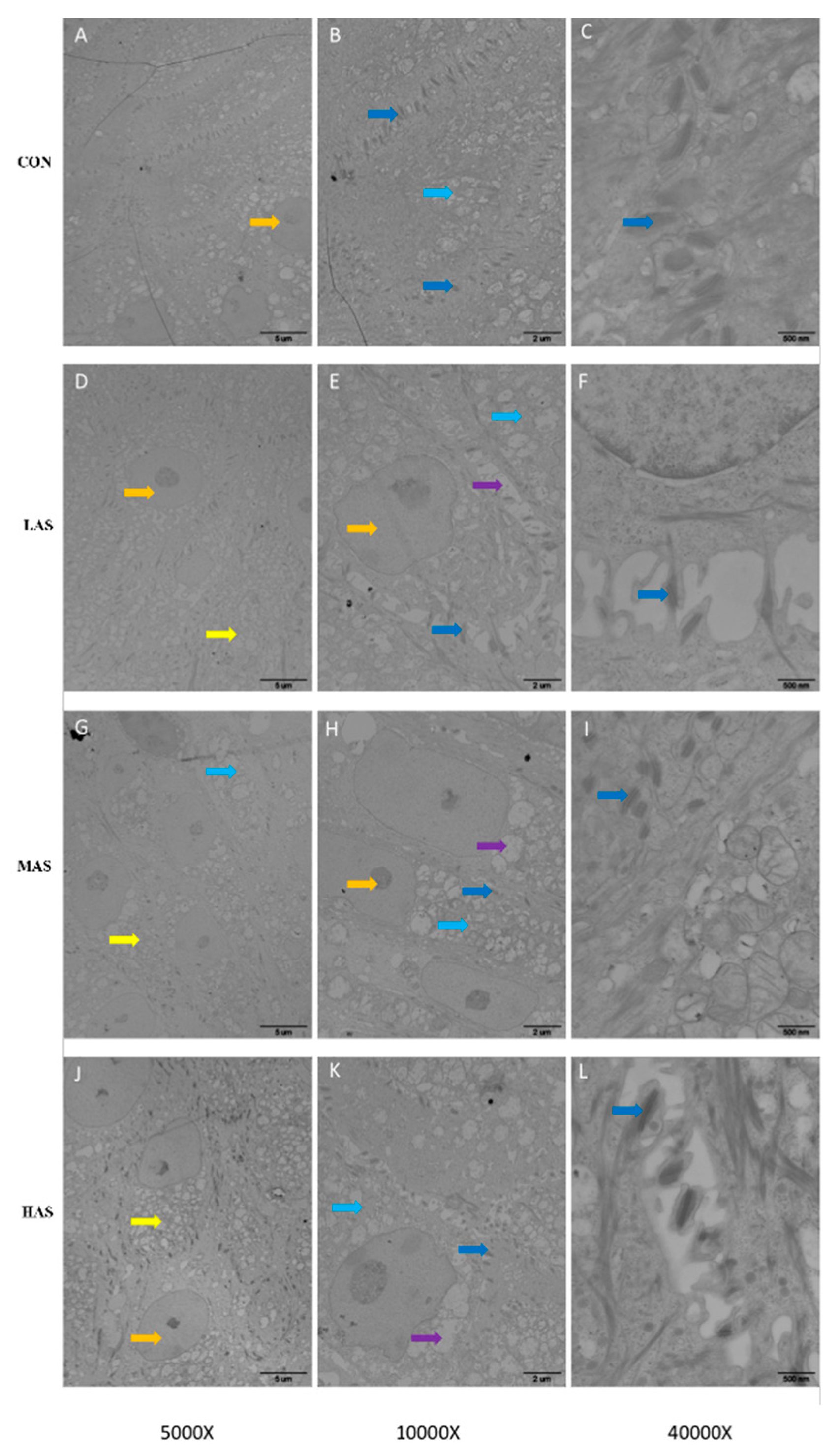
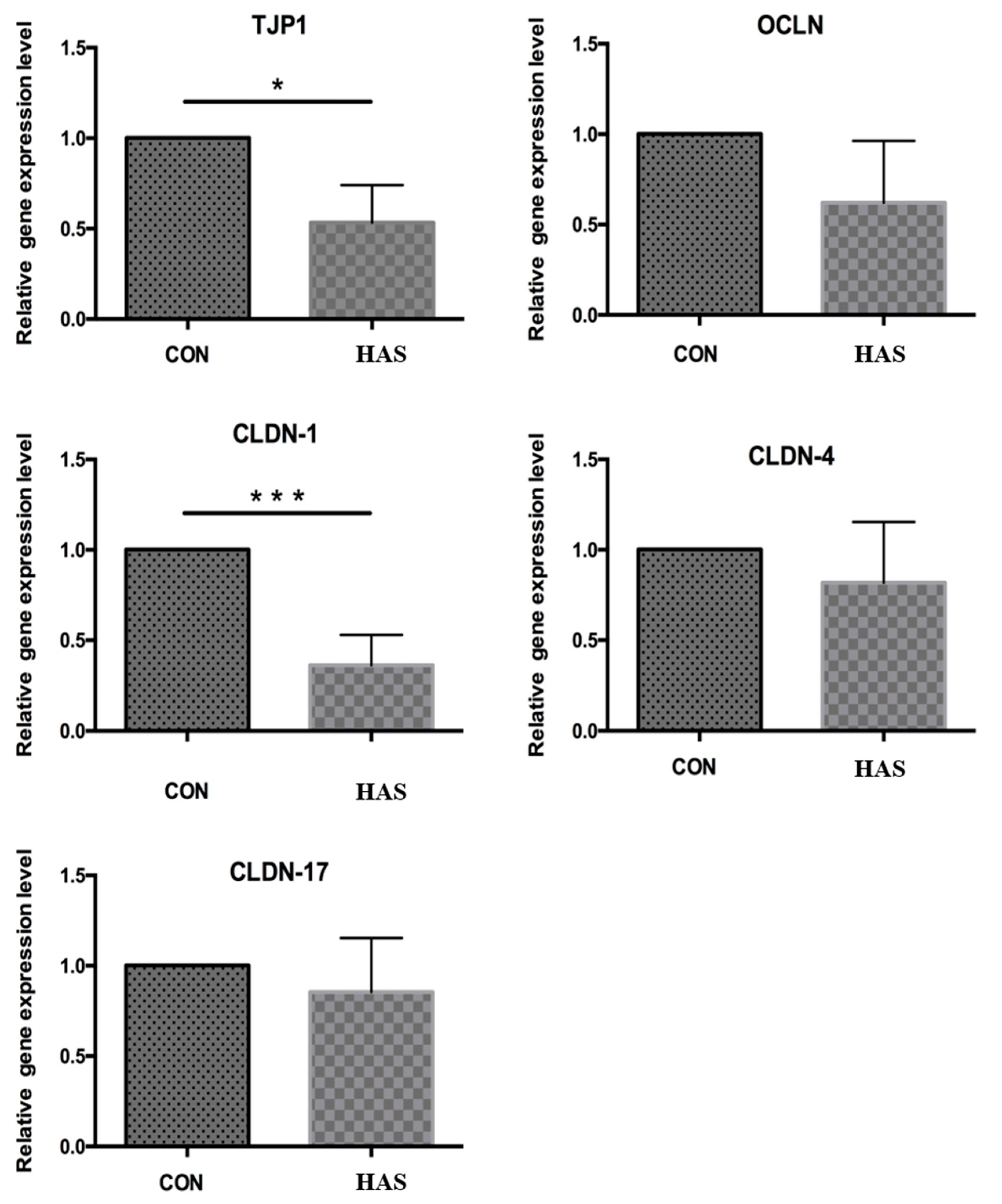
3.3. Effect of High Sulfur Diet on Rumen Epithelial Barrier Function
4. Discussion
4.1. Effect of High Sulfur Diet on Rumen Fermentation
4.2. Effect of High Sulfur Diet on Rumen Microflora
4.3. Effect of High Sulfur Diet on Rumen Epithelial Barrier Function
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Felix, T.L.; Radumz, A.E.; Loerch, S.C. Effects of limit feeding corn or dried distillers’ grains with soluble at 2 intakes during the growing phase on the performance or feedlot cattle. J. Anim. Sci. 2011, 89, 2273–2279. [Google Scholar] [CrossRef]
- Wu, H.; Meng, Q.; Yu, Z. Effect of pH buffering capacity and sources of dietary sulfur on rumen fermentation, sulfide production, methane production, sulfate reducing bacteria, and total Archaea in in vitro rumen cultures. Bioresour. Technol. 2015, 186, 25–33. [Google Scholar] [CrossRef]
- Coleman, G.S. A sulfate-reducing bacterium from the sheep rumen. J. Gen. Microbiol. 1960, 22, 423–436. [Google Scholar] [CrossRef] [Green Version]
- Menke, K.H. The gas production method. J. Agric. Sci. Camb. 1979, 93, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Morrison, M. Improved extraction of PCR-quality community DNA from digesta and fecal samples. Biotechniques 2004, 36, 808–812. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Fang, L.; Meng, Q.; Li, S.; Chai, S.; Liu, S.; Schonewille, J.T. Assessment of Ruminal Bacterial and Archaeal Community Structure in Yak (Bos grunniens). Front. Microbiol. 2017, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Odongo, N.E.; AlZahal, O.; Lindinger, M.I.; Duffield, T.F.; Valdes, E.V.; Terrell, S.P.; McBride, B.W. Effects of mild heat stress and grain challenge on acid-base balance and rumen tissue histology in lambs. J. Anim. Sci. 2006, 84, 447–455. [Google Scholar] [CrossRef]
- Steele, M.A.; Croom, J.; Kahler, M.; AlZahal, O.; Hook, S.E.; Plaizier, K.; McBride, B.W. Bovine rumen epithelium undergoes rapid structural adaptations during grain-induced subacute ruminal acidosis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, 1515–1523. [Google Scholar] [CrossRef] [Green Version]
- Graham, C.; Simmons, N.L. Functional organization of the bovine rumen epithelium. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Xu, T.; Liu, Y.; Zhu, W.; Mao, S. A high-grain diet causes massive disruption of ruminal epithelial tight junctions in goats. Am. J. Physiol. Regul. Intergr. Comp. Physiol. 2013, 305, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Gibson, G.R.; Cummings, J.H.; Macfarlane, G.T. Competition for hydrogen between sulphate-reducing bacteria and methanogenic bacteria from the human large intestine. J. Appl. Microbiol. 2010, 65, 241–247. [Google Scholar] [CrossRef]
- Hunerberg, M.; McGinn, S.M.; Beauchemin, K.A.; Okine, E.K.; Harstad, O.M.; McAllister, T.A. Effects of dried distillers grains plus soluble on enteric methane emissions and nitrogen excretion from growing beef cattle. J. Anim. Sci. 2013, 91, 2846–2857. [Google Scholar] [CrossRef] [Green Version]
- Morrison, M.; Murray, R.M.; Boniface, A.N. Nutrient metabolism and rumen micro-organisms in sheep fed a poor-quality tropical grass hay supplemented with sulphate. J. Agric. Sci. 1990, 115, 269–275. [Google Scholar] [CrossRef]
- Ballard, F.J. Supply and utilization of acetate in mammals. Am. J. Clin. Nutr. 1972, 25, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Zijderveld, S.M.V.; Gerrits, W.J.J.; Apajalahti, J.A.; Newbold, J.R.; Dijkstra, J.; Leng, R.A.; Perdok, H.B. Nitrate and sulfate: Effective alternative hydrogen sinks for mitigation of ruminal methane production in sheep. J. Dairy Sci. 2010, 93, 5856–5866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klevenhusen, F.; Hollmann, M.; Podstatzky-Lichtenstein, L.; Krametter-Frötscher, R.; Aschenbach, J.R.; Zebeli, Q. Feeding barley grain-rich diets altered electrophysiological properties and permeability of the ruminal wall in a goat model. J. Dairy Sci. 2013, 96, 2293–2302. [Google Scholar] [CrossRef] [Green Version]
- Evans, N.J.; Brown, J.M.; Murray, R.D.; Getty, B.; Birtles, R.J.; Hart, C.A.; Carter, S.D. Characterization of novel bovine gastrointestinal tract Treponema isolates and comparison with bovine digital dermatitis treponemes. Appl. Environ. Microbiol. 2011, 77, 138–147. [Google Scholar] [CrossRef] [Green Version]
- Spence, C.; Wells, W.G.; Smith, C.J. Characterization of the primary starch utilization operon in the obligate anaerobe Bacteroides fragilis: Regulation by carbon source and oxygen. J. Bacterial. 2006, 188, 4663–4672. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Lozupone, C.A.; Hamady, M.; Knight, R.; Gordon, J.I. Worlds within worlds: Evolution of the vertebrate gut microbiota. Nat. Rev. Microbiol. 2008, 6, 776. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Ye, H.; Liu, J.; Mao, S. High-grain diets altered rumen fermentation and epithelial bacterial community and resulted in rumen epithelial injuries of goats. Appl. Microbiol. Biotechnol. 2017, 101, 6981–6992. [Google Scholar] [CrossRef]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Saresella, M.; Mendozzi, L.; Rossi, V.; Mazzali, F.; Piancone, F.; Larosa, F.; Marventano, I.; Caputo, D.; Felis, G.E.; Clerici, M. Immunological and clinical effect of diet modulation of the gut microbiome in multiple sclerosis patients: A pilot study. Front. Immunol. 2017, 8, 1–11. [Google Scholar] [CrossRef]
- Flemer, B.; Warren, R.D.; Barrett, M.P.; Cisek, K.; Das, A.; Jeffery, I.B.; Hurley, E.; Micheal, O.R.; Shanahan, F.; Paul, W.T. The oral microbiota in colorectal cancer is distinctive and predictive. Gut 2018, 67, 1454–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, F.; Zheng, R.D.; Sun, X.Q.; Ding, W.J.; Wang, X.Y.; Fan, J.G. Gut microbiota dysbiosis in patients with non-alcoholic fatty liver disease. Hepatobiliary Pancreat. Dis. Int. 2017, 16, 375–381. [Google Scholar] [CrossRef]
- Dirksen, G.; Liebich, H.; Mayer, E. Adaptive changes of the ruminal mucosa and their functional and clinical significance. Bovine Pract. 1985, 20, 116–120. [Google Scholar]
- Bull, L.S.; Bush, L.J.; Friend, J.D.; Harris, B.; Jones, E.W. Incidence of ruminal parakeratosis in calves fed different rations and its relation to volatile fatty acid absorption. J. Dairy Sci. 1965, 48, 1459–1466. [Google Scholar] [CrossRef]
- Kleen, J.L.; Hooijer, G.A.; Rehage, J.; Noordhuizen, J.P. Subacute ruminal acidosis (SARA): A review. J. Vet. Med. A 2003, 50, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Khafipour, E.; Plaizier, J.; Aikman, P.C.; Krause, D. Population structure of rumen Escherichia coli associated with subacute ruminal acidosis (SARA) in dairy cattle. J. Dairy Sci. 2011, 94, 351–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steele, M.A.; Greenwood, S.L.; Croom, J.; Mcbride, B.W. An increase in dietary non-structural carbohydrates alters the structure and metabolism of the rumen epithelium in lambs. Can. J. Plant Sci. 2012, 92, 123–130. [Google Scholar] [CrossRef]
- Yang, S. Study on Effect of Subacute Ruminal Acidosis on Rumen Epithelial Barrier Function in Dairy Goats. Master’s Thesis, Inner Mongolia Agricultural University, Huhhot, China, 2014. [Google Scholar]
- Chen, M. Effect of Subacute Ruminal Acidosis on Rumen Epithelium Permeability and Intercellular Junction Protein Expression in Dairy Goats. Master’s Thesis, Inner Mongolia Agricultural University, Huhhot, China, 2016. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Treatment 1 | |||
|---|---|---|---|---|
| CON | LAS | MAS | HAS | |
| Ingredient (%DM) | ||||
| Steam-flake corn | 40.00 | 40.00 | 40.00 | 40.00 |
| Whole corn silage | 40.00 | 40.00 | 40.00 | 40.00 |
| Cottonseed meal | 4.50 | 4.50 | 4.50 | 4.50 |
| Brewer’s grain | 13.40 | 13.40 | 13.40 | 13.40 |
| Sodium sulfate | - | 0.89 | 1.78 | 2.66 |
| Salt | 0.50 | 0.50 | 0.50 | 0.50 |
| Premix 2 | 0.50 | 0.50 | 0.50 | 0.50 |
| Calcium hydrogen phosphate | 0.60 | 0.60 | 0.60 | 0.60 |
| Limestone | 0.50 | 0.50 | 0.50 | 0.50 |
| Nutritional composition 3 | ||||
| ME (MJ/kg) | 7.34 | 7.34 | 7.34 | 7.34 |
| CP (%DM) | 13.61 | 13.61 | 13.61 | 13.61 |
| NDF (%DM) | 43.20 | 43.20 | 43.20 | 43.20 |
| ADF (%DM) | 22.89 | 22.89 | 22.89 | 22.89 |
| Ca (%DM) | 0.47 | 0.47 | 0.47 | 0.47 |
| P (%DM) | 0.22 | 0.22 | 0.22 | 0.22 |
| S (%DM) | 0.40 | 0.60 | 0.80 | 1.00 |
| Target | Accession Number | Primer Sequences 1 | PCR Product Size (bp) |
|---|---|---|---|
| Claudin-1 | NM_001001854.2 | F: 5′CCGTGCCTTGATGGTGATTGG3′ | 107 |
| R: 5′TCTTCTGTGCCTCGTCGTCTTC3′ | |||
| Claudin-4 | NM_001014391.2 | F: 5′ATCGGCAGCAACATCGTCAC3′ | 107 |
| R: 5′AGCAGCGAGTCGTACACCTT3′ | |||
| Claudin-17 | XM_010800919.3 | F: 5′GCGTCCGACAAGCCAAGG3′ | 133 |
| R: 5′CCAACAAGCAGAGCAATCACAGA 3′ | |||
| Occludin | NM_001082433.2 | F: 5′GCTACGGCTATGGCTACGGTTA3′ | 194 |
| R: 5′CAGGACGGCGGTCACTATTATCA 3′ | |||
| TJP1 | - | F: 5′ CGGATGGTGCTACAAGTGATGAC3′ | 194 |
| R: 5′CGCCTTCTGTGTCTGTGTCTTC3′ | |||
| β-actin | NM_173979.3 | F: 5′CATCGGCAATGAGCGGTTCC3′ | 145 |
| R: 5′CGTGTTGGCGTAGAGGTCCTT3′ |
| Item 1 | Treatment | SEM 2 | p-Value | Contrast 3 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.40% | 0.50% | 0.60% | 0.70% | 0.80% | 0.90% | 1.00% | 1.10% | 1.20% | PL | PQ | |||
| Gas production dynamic (mL/0.2 g DM) | |||||||||||||
| GP24 | 55.04 ab | 55.72 a | 54.22 abc | 55.59 a | 55.25 ab | 52.96c | 53.29 c | 53.97 abc | 53.70 bc | 0.21 | <0.01 | <0.01 | 0.94 |
| GP48 | 62.93 abc | 64.18 a | 61.88 abcd | 63.53 ab | 63.42 ab | 59.85 d | 60.73 cd | 62.27 abcd | 61.47 bcd | 0.31 | <0.01 | <0.01 | 0.61 |
| B | 63.88 abc | 65.36 a | 64.97 ab | 65.47 a | 62.05 bcd | 59.48 d | 61.90 cd | 63.14 abc | 60.98 cd | 0.48 | <0.01 | <0.01 | 0.75 |
| c (h−1) | 0.094 c | 0.096 bc | 0.096 bc | 0.099 abc | 0.101 ab | 0.101 ab | 0.101 ab | 0.104 a | 0.106 a | 0.001 | 0.01 | <0.01 | 0.79 |
| 24 h fermentation parameter | |||||||||||||
| NH3-N (mg/100 mL) | 17.41 | 25.69 | 18.44 | 18.87 | 22.39 | 19.02 | 18.86 | 15.69 | 19.81 | 0.83 | 0.19 | 0.29 | 0.63 |
| TVFA (mmol/L) | 47.37 a | 53.62 a | 46.02 ab | 47.77 a | 38.8 b | 51.89 a | 53.57 a | 49.99 a | 49.66 a | 1.12 | 0.02 | 0.40 | 0.22 |
| Acetate (%) | 64.01 | 64.06 | 63.18 | 64.51 | 62.04 | 63.45 | 64.10 | 63.70 | 62.98 | 0.24 | 0.13 | 0.31 | 0.59 |
| Propionate (%) | 19.06 | 18.70 | 19.01 | 18.52 | 19.16 | 19.10 | 18.54 | 18.50 | 19.10 | 0.17 | 0.37 | 0.70 | 0.67 |
| Isobutyrate (%) | 1.36 | 1.57 | 1.55 | 1.61 | 1.68 | 1.45 | 1.69 | 1.77 | 1.60 | 0.11 | 0.46 | 0.09 | 0.45 |
| Butyrate (%) | 12.11 | 12.21 | 12.67 | 12.02 | 13.37 | 12.53 | 12.23 | 12.49 | 12.76 | 0.20 | 0.18 | 0.24 | 0.41 |
| Isovalerate (%) | 2.67 | 2.59 | 2.75 | 2.50 | 2.90 | 2.61 | 2.49 | 2.63 | 2.65 | 0.09 | 0.39 | 0.68 | 0.82 |
| Valerate (%) | 0.79 | 0.87 | 0.83 | 0.85 | 0.85 | 0.86 | 0.95 | 0.91 | 0.90 | 0.02 | 0.81 | 0.12 | 0.82 |
| A/P | 3.36 | 3.43 | 3.32 | 3.49 | 3.24 | 3.32 | 3.46 | 3.45 | 3.30 | 0.05 | 0.19 | 0.91 | 0.94 |
| 48 h fermentation parameter | |||||||||||||
| NH3-N (mg/100 mL) | 26.62 | 27.99 | 27.8 | 29.67 | 27.16 | 25.53 | 28.49 | 31.56 | 28.18 | 0.79 | 0.86 | 0.48 | 0.89 |
| TVFA (mmol/L) | 54.67 | 46.69 | 50.39 | 49.15 | 59.54 | 49.17 | 57.50 | 62.16 | 62.26 | 1.82 | 0.29 | 0.04 | 0.27 |
| Acetate (%) | 63.11 | 63.12 | 62.40 | 62.53 | 62.81 | 63.28 | 62.55 | 62.93 | 62.69 | 0.18 | 0.97 | 0.79 | 0.75 |
| Propionate (%) | 18.50 | 18.44 | 18.45 | 18.43 | 18.41 | 18.49 | 18.31 | 18.06 | 18.18 | 0.06 | 0.81 | 0.11 | 0.51 |
| Isobutyrate (%) | 2.11 | 2.22 | 2.12 | 2.12 | 2.33 | 2.08 | 2.40 | 2.32 | 2.51 | 0.07 | 0.86 | 0.17 | 0.57 |
| Butyrate (%) | 11.88 | 11.85 | 12.32 | 12.25 | 12.03 | 11.98 | 12.19 | 11.98 | 12.13 | 0.08 | 0.92 | 0.67 | 0.51 |
| Isovalerate (%) | 3.48 | 3.50 | 3.65 | 3.63 | 3.33 | 3.27 | 3.43 | 3.45 | 3.40 | 0.05 | 0.77 | 0.33 | 0.90 |
| Valerate (%) | 0.92 | 0.87 | 1.07 | 1.03 | 1.09 | 0.91 | 1.12 | 1.25 | 1.10 | 0.04 | 0.26 | 0.03 | 0.83 |
| A/P | 3.41 | 3.42 | 3.38 | 3.39 | 3.41 | 3.42 | 3.42 | 3.48 | 3.45 | 0.02 | 0.93 | 0.27 | 0.48 |
| Item | Treatment | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| CON | LAS | MAS | HAS | |||
| NH3-N (mg/100 mL) | 18.63 a | 5.98 c | 6.88 bc | 14.91 ab | 1.66 | <0.01 |
| TVFA (mmol/L) | 41.72 b | 38.51 b | 39.15 b | 52.87 a | 1.35 | <0.01 |
| Acetate (%) | 67.66 ab | 67.81 a | 66.42 b | 66.41 b | 0.23 | 0.03 |
| Propionate (%) | 20.18 a | 16.09 c | 20.03 a | 18.14 b | 0.38 | <0.01 |
| Isobutyrate (%) | 0.63 b | 1.03 a | 0.71 b | 0.57 b | 0.05 | <0.01 |
| Butyrate (%) | 9.19 c | 13.56 a | 11.13 b | 13.24 a | 0.35 | <0.01 |
| Isovalerate (%) | 1.70 a | 1.30 b | 1.33 b | 1.18 b | 0.06 | 0.01 |
| Valerate (%) | 0.63 a | 0.20 c | 0.37 b | 0.46 b | 0.03 | <0.01 |
| A/P | 3.37 bc | 4.24 a | 3.33 c | 3.69 b | 0.08 | <0.01 |
| Item 1 | Observed Species | Shannon | Simpson | Ace | Chao | Good’s Coverage |
|---|---|---|---|---|---|---|
| CON | 1580.50 | 6.08 | 0.0074 | 1857.33 | 1882.86 | 0.9826 |
| LAS | 1467.25 | 5.91 | 0.0089 | 1757.12 | 1770.60 | 0.9830 |
| MAS | 1522.00 | 5.95 | 0.0095 | 1813.51 | 1836.60 | 0.9824 |
| HAS | 1518.13 | 6.01 | 0.0083 | 1797.46 | 1821.01 | 0.9830 |
| SEM | 16.36 | 0.03 | 0.0006 | 17.54 | 18.68 | 0.0002 |
| p-Value | 0.10 | 0.33 | 0.67 | 0.25 | 0.21 | 0.65 |
| Item 1 | Treatment 2 | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| CON | LAS | MAS | HAS | |||
| Epithelial papilla length/mm | 5.24 c | 7.33 bc | 8.67 ab | 10.94 a | 0.58 | <0.01 |
| spinous layer + basal layer/µm | 66.16 b | 67.23 b | 73.42 ab | 80.10 a | 1.86 | 0.02 |
| cuticle + granular layer/µm | 16.88 | 15.01 | 18.25 | 18.57 | 0.62 | 0.17 |
| Total epithelium/µm | 83.04 b | 82.24 b | 91.67 ab | 98.67 a | 2.07 | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Li, Y.; Meng, Q.; Zhou, Z. Effect of High Sulfur Diet on Rumen Fermentation, Microflora, and Epithelial Barrier Function in Steers. Animals 2021, 11, 2545. https://doi.org/10.3390/ani11092545
Wu H, Li Y, Meng Q, Zhou Z. Effect of High Sulfur Diet on Rumen Fermentation, Microflora, and Epithelial Barrier Function in Steers. Animals. 2021; 11(9):2545. https://doi.org/10.3390/ani11092545
Chicago/Turabian StyleWu, Hao, Yan Li, Qingxiang Meng, and Zhenming Zhou. 2021. "Effect of High Sulfur Diet on Rumen Fermentation, Microflora, and Epithelial Barrier Function in Steers" Animals 11, no. 9: 2545. https://doi.org/10.3390/ani11092545
APA StyleWu, H., Li, Y., Meng, Q., & Zhou, Z. (2021). Effect of High Sulfur Diet on Rumen Fermentation, Microflora, and Epithelial Barrier Function in Steers. Animals, 11(9), 2545. https://doi.org/10.3390/ani11092545