
Growth Performance and Plasma Metabolites of Grazing Beef Cattle Backgrounded on Buffel or Buffel-Desmanthus Mixed Pastures

 , , and
, , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Animal Management
2.3. Pasture Sampling and Analysis
2.4. Faecal Sampling and Analysis
2.5. Plasma Metabolites Analysis
2.6. Statistical Analysis
3. Results
3.1. Rainfall and Pasture Characteristics
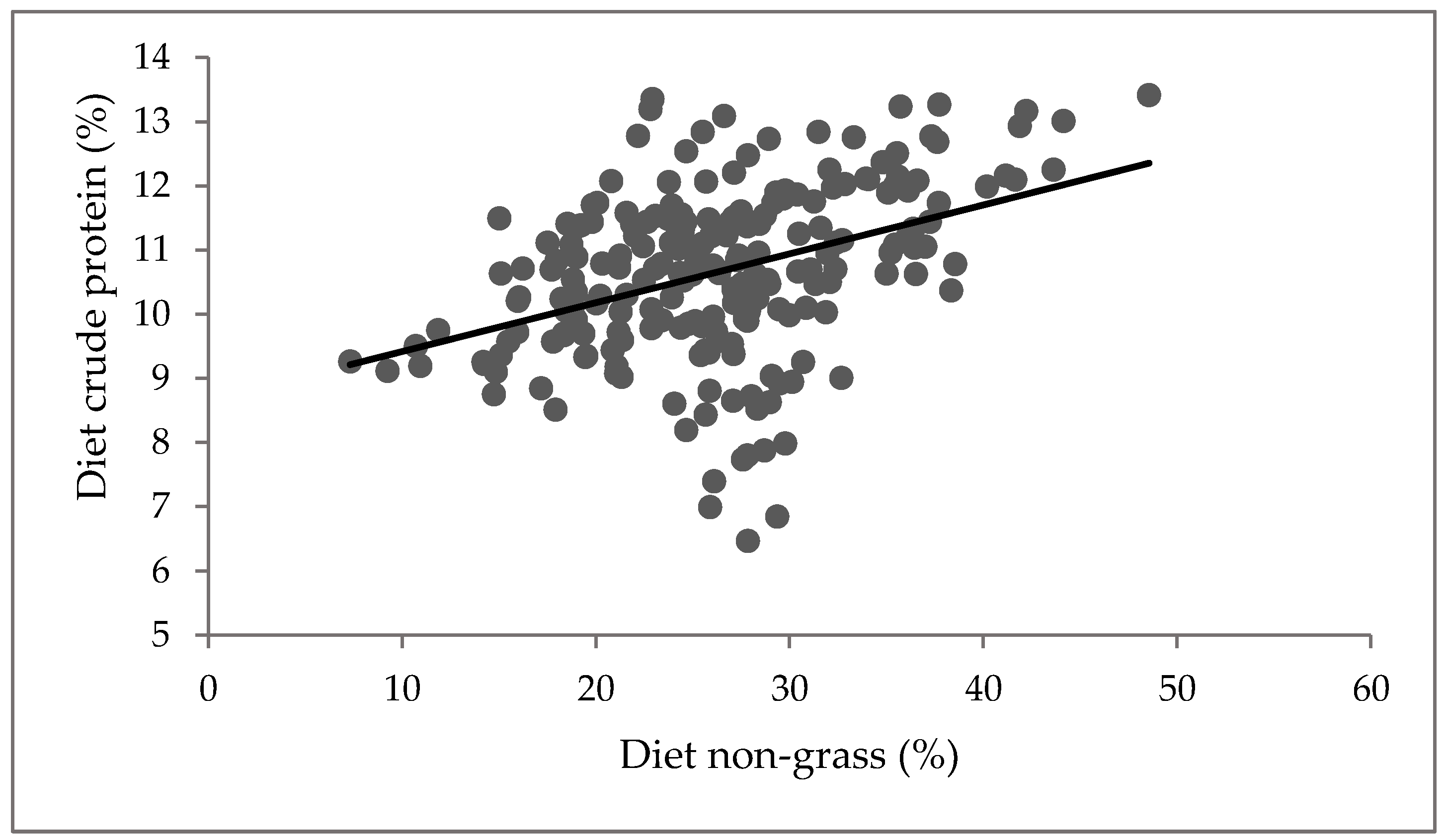
3.2. Diet Selected during Grazing
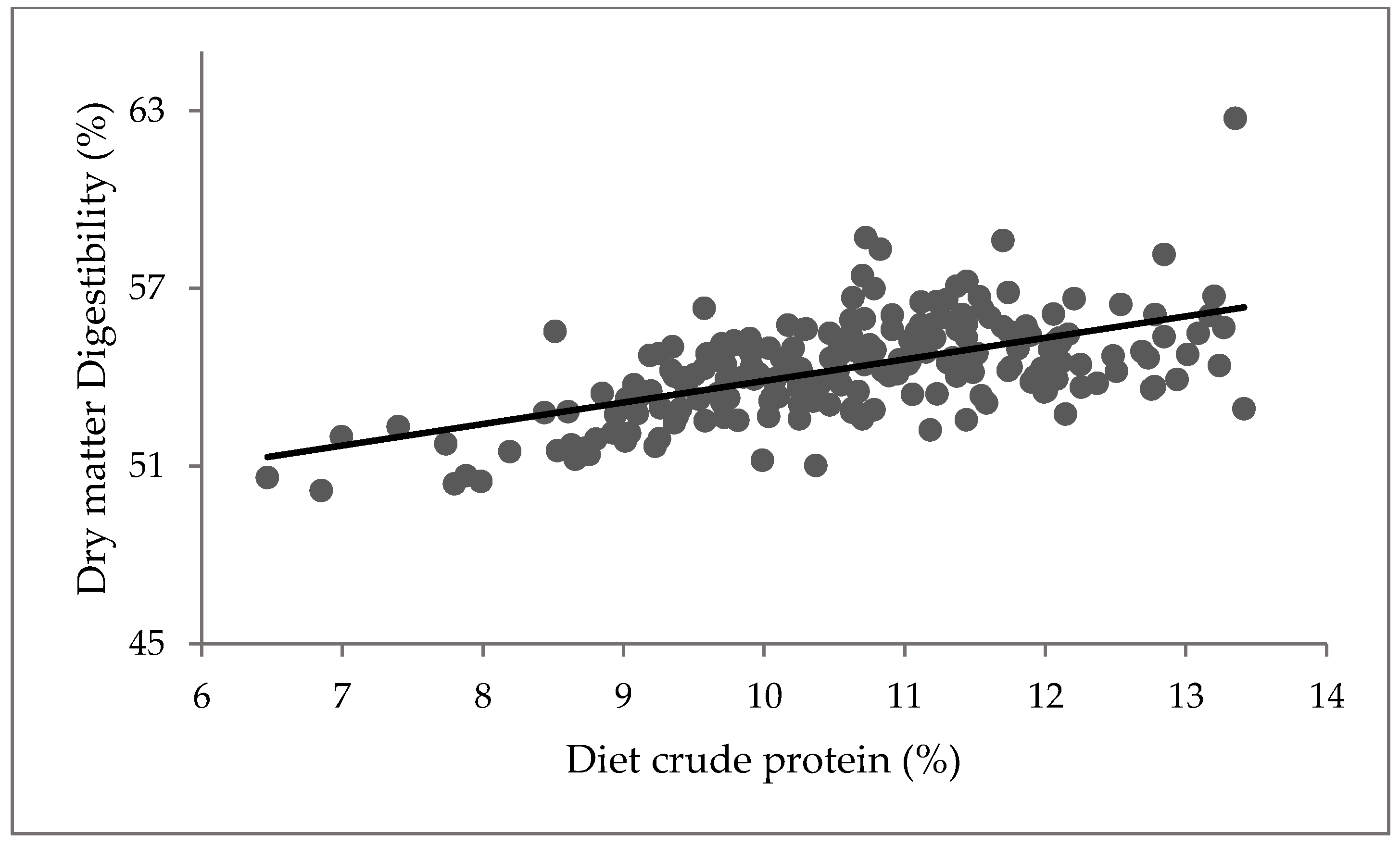
3.3. Plasma Metabolites
3.4. Growth Performance
4. Discussion
4.1. Pasture Characteristics
4.2. Diet Selected during Grazing
4.3. Plasma Metabolites
4.4. Growth Performance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oosting, S.J.; Udo, H.M.J.; Viets, T.C. Development of livestock production in the tropics: Farm and farmers’ perspectives. Animal 2014, 8, 1238–1248. [Google Scholar] [CrossRef] [PubMed]
- Winter, W.H.; Winks, L.; Seebeck, R.M. Sustaining productive pastures in the tropics 10. Forage and feeding systems for cattle. Trop. Grassl. 1991, 25, 145–152. [Google Scholar]
- Hattersley, P.W. The distribution of C3 and C4 grasses in Australia in relation to climate. Oecologia 1983, 57, 113–128. [Google Scholar] [CrossRef]
- Hunt, L.P.; Mcivor, J.G.; Grice, A.C.; Bray, S.G. Principles and guidelines for managing cattle grazing in the grazing lands of northern Australia: Stocking rates, pasture resting, prescribed fire, paddock size and water points: A review. Rangel. J. 2014, 36, 105–119. [Google Scholar] [CrossRef]
- Edwards, E.J.; Smith, S.A. Phylogenetic analyses reveal the shady history of C4 grasses. Proc. Natl. Acad. Sci. USA 2010, 107, 2532–2537. [Google Scholar] [CrossRef]
- Poppi, D.P.; Quigley, S.P.; Silva, T.A.C.C.; McLennan, S.R. Challenges of beef cattle production from tropical pastures. Rev. Bras. Zootec. 2018, 47, e20160419. [Google Scholar] [CrossRef]
- Allison, C.D. Factors affecting forage intake by range ruminants: A review. J. Range Manag. 1985, 38, 305–311. [Google Scholar] [CrossRef]
- McCown, R.L. The climatic potential for beef cattle production in tropical Australia: Part I-Simulating the annual cycle of liveweight change. Agric. Syst. 1981, 6, 303–317. [Google Scholar] [CrossRef]
- Hill, J.O.; Coates, D.B.; Whitbread, A.M.; Clem, R.L.; Robertson, M.J.; Pengelly, B.C. Seasonal changes in pasture quality and diet selection and their relationship with liveweight gain of steers grazing tropical grass and grass-legume pastures in northern Australia. Anim. Prod. Sci. 2009, 49, 983–993. [Google Scholar] [CrossRef]
- Prudhomme, R.; Brunelle, T.; Dumas, P.; Le Moing, A.; Zhang, X. Assessing the impact of increased legume production in Europe on global agricultural emissions. Reg. Environ. Change 2020, 20, 91. [Google Scholar] [CrossRef]
- Da Motta, E.A.M.; Dall-Agnol, M.; Rios, E.F.; Souza, C.H.L.; Weiler, R.L.; Brunes, A.P.; Simioni, C.; Rockenbach de Ávila, M.; Neto, D.; Felix, L.D.; et al. Agronomic performance of interspecific Paspalum hybrids under nitrogen fertilization or mixed with legumes. Agrosyst. Geosci. Environ. 2020, 3, e20127. [Google Scholar]
- Castro-Montoya, J.M.; Dickhoefer, U. The nutritional value of tropical legume forages fed to ruminants as affected by their growth habit and fed form: A systematic review. Anim. Feed Sci. Technol. 2020, 269, 114641. [Google Scholar] [CrossRef]
- Mannetje, L. Harry stobbs memorial lecture, 1994: Potential and prospects of legume-based pastures in the tropics. Trop. Grassl. 1997, 31, 81–94. [Google Scholar]
- Coates, D.B.; Miller, C.P.; Hendricksen, R.E.; Jones, R.J. Stability and productivity of Stylosanthes pastures in Australia. II. Animal production from Stylosanthes pastures. Trop. Grassl. 1997, 31, 494–502. [Google Scholar]
- Clem, R.L.; Hall, T.J. Persistence and productivity of tropical pasture legumes on three cracking clay soils (Vertisols) in north-eastern Queensland. Aust. J. Exp. Agric. 1994, 34, 161–171. [Google Scholar] [CrossRef]
- Pengelly, B.C.; Conway, M.J. Pastures on cropping soils: Which tropical pasture legume to use? Trop. Grassl. 2000, 34, 162–168. [Google Scholar]
- Meat & Livestock Australia (MLA). State of the Industry Report: The Australian Red Meat and Livestock Industry; Meat & Livestock Australia (MLA): Sydney, Australia, 2019. [Google Scholar]
- Soil Science Australia State Soils. Available online: https://www.soilscienceaustralia.org.au/about/about-soil/state-soil (accessed on 15 December 2020).
- Weston, E.J.; Ellis, C.M.; Harbison, J. Assessment of the Agricultural and Pastoral Potential of Queensland; Queensland Department of Primary Industries: Brisbane, Australia, 1983; ISBN 0724221433.
- Gardiner, C.P.; Swan, S.J. Abandoned pasture legumes offer potential economic and environmental benefits in semiarid clay soil rangelands. In Proceedings of the Australian Rangeland Society 15th Biennial Conference Proceedings, Charters Towers, Australia, 28 September–2 October 2008; p. 93. [Google Scholar]
- Meat & Livestock Australia (MLA) Legumes for Clay Soils. Available online: https://www.mla.com.au/download/finalreports?itemId=874 (accessed on 29 November 2020).
- Vandermeulen, S.; Singh, S.; Ramírez-Restrepo, C.A.; Kinley, R.D.; Gardiner, C.P.; Holtum, J.A.M.; Hannah, I.; Bindelle, J. In vitro assessment of ruminal fermentation, digestibility and methane production of three species of Desmanthus for application in northern Australian grazing systems. Crop Pasture Sci. 2018, 69, 797–807. [Google Scholar] [CrossRef]
- Suybeng, B.; Charmley, E.; Gardiner, C.P.; Malau-Aduli, B.S.; Malau-Aduli, A.E.O. Supplementing Northern Australian beef cattle with Desmanthus tropical legume reduces in-vivo methane emissions. Animals 2020, 10, 2097. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.O.; Clem, R.L.; Robertson, M.J.; Pengelly, B.C.; Whitbread, A. Beef production from tropical pasture legumes on cropping soils. In Proceedings of the 25th Biennial Conference of the Australian Society of Animal Production, Melbourne, Australia, 4–8 July 2004; CSIRO Publishing: Melbourne, Australia. [Google Scholar]
- Gardiner, C.; Kempe, C.; Kempe, N.; Campbell, G.; Heitor, F.; Malau-Aduli, A.E.O.; Walker, G.; Suybeng, B.; Mwangi, F. Progardes Desmanthus: An update. In Proceedings of the Northern Beef Research Update Conference, Brisbane, Australia, 19–22 August 2019; p. 33. [Google Scholar]
- Gardiner, C.; Parker, A. Steer liveweight gains on ProgardesTM Desmanthus/Buffel pastures in Queensland. In Proceedings of the 2nd Australian and New Zealand societies of animal production joint conference, Lincoln, New Zealand, 2–5 July 2012. [Google Scholar]
- Marsetyo, D.R.; Rusiyantono, Y.; Syukur, S.H. The effect of supplementation of different legume leaves on feed intake, digestion and growth of Kacang goats given Mulato grass. J. Agric. Sci. Technol. 2017, 7, 117–122. [Google Scholar] [CrossRef][Green Version]
- Rangel, J.H.D.A.; Gardiner, C.P. Stimulation of wool growth by Desmanthus spp. as a supplement to a diet of Mitchell grass hay. Trop. Grassl. 2009, 43, 106–111. [Google Scholar]
- Kanani, J.; Lukefahr, S.D.; Stanko, R.L. Evaluation of tropical forage legumes (Medicago sativa, Dolichos lablab, Leucaena leucocephala and Desmanthus bicornutus) for growing goats. Small Rumin. Res. 2006, 65, 1–7. [Google Scholar] [CrossRef]
- Ndlovu, T.; Chimonyo, M.; Okoh, A.I.; Muchenje, V.; Dzama, K.; Raats, J.G. Assessing the nutritional status of beef cattle: Current practices and future prospects. Afr. J. Biotechnol. 2007, 6, 2727–2734. [Google Scholar]
- Flier, J.; Maratos-Flier, E. Energy homeostasis and body weight. Curr. Biol. 2000, 10, R215–R217. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Beef Cattle, 7th ed.; National Academies Press: Washington, DC, USA, 1996; ISBN 978-0-309-06934-2. [Google Scholar]
- Nicholson, M.J.; Sayers, A.R. Repeatability, reproducibility and sequential use of condition scoring of Bos indicus cattle. Trop. Anim. Health Prod. 1987, 19, 127–135. [Google Scholar] [CrossRef]
- Chester-Jones, H.; Fontenot, J.P.; Veit, H.P. Physiological and pathological effects of feeding high levels of magnesium to steers. J. Anim. Sci. 1990, 68, 4400–4413. [Google Scholar] [CrossRef]
- Pambu-Gollah, R.; Cronjé, P.B.; Casey, N.H. An evaluation of the use of blood metabolite concentrations as indicators of nutritional status in free-ranging indigenous goats. S. Afr. J. Anim. Sci. 2000, 30, 115–120. [Google Scholar] [CrossRef]
- Murillo-Ortiz, M.; Mellado-Bosque, M.; Herrera-Torres, E.; Reyes-Estrada, O.; Carrete-Carreón, F.O. Seasonal diet quality and metabolic profiles of steers grazing on Chihuahuan desert rangeland. Livestig. Sci. 2014, 165, 61–65. [Google Scholar] [CrossRef]
- Abeni, F.; Bergoglio, G.; Masoero, G.; Terzano, G.M.; Allegrini, S. Plasma hormones and metabolites in Piedmontese cows during late pregnancy: Relationships with calf birth weight. J. Anim. Sci. 2004, 82, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Movahedi, B.; Foroozandeh, A.D.; Shakeri, P. Effects of different forage sources as a free-choice provision on the performance, nutrient digestibility, selected blood metabolites and structural growth of Holstein dairy calves. J. Anim. Physiol. Anim. Nutr. 2017, 101, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Soder, K.J.; Sanderson, M.A.; Stack, J.L.; Muller, L.D. Intake and performance of lactating cows grazing diverse forage mixtures. J. Dairy Sci. 2006, 89, 2158–2167. [Google Scholar] [CrossRef]
- Pembleton, K.G.; Hills, J.L.; Freeman, M.J.; McLaren, D.K.; French, M.; Rawnsley, R.P. More milk from forage: Milk production, blood metabolites, and forage intake of dairy cows grazing pasture mixtures and spatially adjacent monocultures. J. Dairy Sci. 2016, 99, 3512–3528. [Google Scholar] [CrossRef] [PubMed]
- Ragni, M.; Colonna, M.A.; Lestingi, A.; Tarricone, S.; Giannico, F.; Marsico, G.; Facciolongo, A.M. Effects of protein sources on performance, carcass composition, blood parameters and meat quality in Charolais heifers. S. Afr. J. Anim. Sci. 2018, 48, 683–694. [Google Scholar] [CrossRef]
- National Health and Medical Research Council. Australian Code of Practice for the Care and Use of Animals for Scientific Purposes, 8th ed.; National Health and Medical Research Council: Canberra, Australia, 2013; ISBN 1864965975.
- Collins, J.; Gardiner, C.P.; Kempe, N.; Hannah, I. Successful pasture development at Cungelella: A grazier, a researcher and a seed company’s perspective. In Proceedings of the Northern Beef Research Update Conference, Leichhardt Hotel, Rockhampton, Australia, 15–18 August 2016. [Google Scholar]
- Gardiner, C.; Kempe, N.; Hannah, I.; Mcdonald, J. PROGARDESTM: A legume for tropical/subtropical semi-arid clay soils. Trop. Grassl. Forrajes Trop. 2013, 1, 78–80. [Google Scholar] [CrossRef]
- Gardiner, C. Developing and commercializing new pasture legumes for clay soils in the semi-arid rangelands of northern Australia: The new Desmanthus cultivars JCU 1–5 and the Progardes story. In Tropical Forage Legumes. Harnessing the Potential of Desmanthus and Other Genera for Heavy Clay Soils; Lazier, J.R., Ahmad, N., Eds.; CABI: Boston, MA, USA, 2016; pp. 283–304. [Google Scholar]
- Miller, C.P.; Stockwell, T.G.H. Sustaining productive pastures in the tropics. 4. Augmenting native pasture with legumes. Trop. Grassl. 1991, 25, 98–103. [Google Scholar]
- Tothill, J.C.; Hargreaves, J.; Jones, R. Botanal-A Comprehensive Sampling and Computing Procedure for Estimating Pasture Yield and Composition. 1. Field Sampling; CSIRO Division of Tropical Crops and Pastures: Queensland, Australia, 1992. [Google Scholar]
- McDonald, C.K.; Corfield, J.P.; Hargreaves, J.N.G.; O’Toole, J.G. Botanal: A Comprehensive Sampling and Computing Procedure for Estimating Pasture Yield and Composition. 3. Field Recording Direct to Computer, 2nd ed.; CSIRO: Brisbane, Australia, 1996.
- Stoddart, L.A. Range capacity determination. Ecology 1935, 16, 531–532. [Google Scholar]
- Sweeney, R.A.; Rexroad, P.R. Comparison of LECO FP-228 “nitrogen determinator” with AOAC copper catalyst Kjeldahl method for crude protein. J. Assoc. Off. Anal. Chem. 1987, 70, 1028–1030. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Goering, H.K.; Van Soest, P.J. Forage Fiber Analysis (Apparatus, Reagents, Procedures, and Some Applications); ARS-USDA: Washington, DC, USA, 1970.
- Faichney, G.; White, G. Methods for the Analysis of Feeds Eaten by Ruminants; CSIRO: Melbourne, Australia, 1983; ISBN 0643035621.
- Clarke, T.; Flinn, P.C.; McGowan, A. Low-cost pepsin-cellulase assays for prediction of digestibility of herbage. Grass Forage Sci. 1982, 37, 147–150. [Google Scholar] [CrossRef]
- Commonwealth Scientific and Industrial Research Organisation. Nutrient Requirements of Domesticated Ruminants; CSIRO Publishing: Collingwood, Australia, 2007; ISBN 9780643092624. [Google Scholar]
- Coates, D.B.; Dixon, R.M. Faecal near infrared reflectance spectroscopy (F.NIRS) measurements of non-grass proportions in the diet of cattle grazing tropical rangelands. Rangel. J. 2007, 29, 51–63. [Google Scholar] [CrossRef]
- Dixon, R.M.; Coates, D.B. Diet quality and liveweight gain of steers grazing Leucaena-grass pasture estimated with faecal near infrared reflectance spectroscopy (F.NIRS). Aust. J. Exp. Agric. 2008, 48, 835–842. [Google Scholar] [CrossRef]
- Coates, D.B. Improving Nutritional Management of Grazing Cattle: Improving Reliability of Faecal NIRS Calibration Equations; Meat & Livestock Australia: North Sydney, Australia, 2004. [Google Scholar]
- Coates, D.B.; Dixon, R.M. Development of near infrared analysis of faeces to estimate non-grass proportions in diets selected by cattle grazing tropical pastures. J. Near Infrared Spectrosc. 2008, 16, 471–480. [Google Scholar] [CrossRef]
- Hacker, J.B.; Waite, R.B. Selecting buffel grass (Cenchrus ciliaris) with improved spring yield in subtropical Australia. Trop. Grassl. 2001, 35, 205–210. [Google Scholar]
- Cowie, B.A.; Thornton, C.M.; Radford, B.J. The Brigalow catchment study: I. Overview of a 40-year study of the effects of land clearing in the brigalow bioregion of Australia. Soil Res. 2007, 45, 479. [Google Scholar] [CrossRef]
- Gulwa, U.; Mgujulwa, N.; Beyene, S.T. Effect of grass-legume intercropping on dry matter yield and nutritive value of pastures in the Eastern Cape Province, South Africa. Univ. J. Agric. Res. 2017, 5, 355–362. [Google Scholar] [CrossRef]
- Lüscher, A.; Suter, M.; Finn, J.; Collins, R.; Gastal, F. Quantification of the Effect of Legume Proportion in the Sward on Yield Advantage and Options to Keep Stable Legume Proportions (over Climatic Zones Relevant for Livestock Production). HAL Arch. Ouv. 2014, 7.3, 35. [Google Scholar]
- Bork, E.W.; Gabruck, D.T.; McLeod, E.M.; Hall, L.M. Five-year forage dynamics arising from four legume–grass seed mixes. Agron. J. 2017, 109, 2789–2799. [Google Scholar] [CrossRef]
- Khatiwada, B.; Acharya, S.N.; Larney, F.J.; Lupwayi, N.Z.; Smith, E.G.; Islam, M.A.; Thomas, J.E. Benefits of mixed grass–legume pastures and pasture rejuvenation using bloat-free legumes in western Canada: A review. Can. J. Plant Sci. 2020, 100, 463–476. [Google Scholar] [CrossRef]
- Thomas, R.J. Role of legumes in providing N for sustainable tropical pasture systems. In Developments in Plant and Soil Sciences; Ladha, J.K., Peoples, M.B., Eds.; Springer Science and Business Media, B.V.: Dordrecht, The Netherlands, 1995; Volume 65, pp. 103–118. ISBN 978-94-010-4029-7. [Google Scholar]
- Thomas, R.J. The role of the legume in the nitrogen cycle of productive and sustainable pastures. Grass Forage Sci. 1992, 47, 133–142. [Google Scholar] [CrossRef]
- Durmic, Z.; Ramírez-Restrepo, C.A.; Gardiner, C.; O’Neill, C.J.; Hussein, E.; Vercoe, P.E. Differences in the nutrient concentrations, in vitro methanogenic potential and other fermentative traits of tropical grasses and legumes for beef production systems in northern Australia. J. Sci. Food Agric. 2017, 97, 4075–4086. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, D.W.; Williamson, P.J.; Nolan, J.V.; Kempton, T.J.; Leng, R.A. The roles of energy- or protein-rich supplements in the subtropics for young cattle consuming basal diets that are low in digestible energy and protein. J. Agric. Sci. 1983, 100, 657–666. [Google Scholar] [CrossRef]
- Leng, R.A. Factors affecting the utilization of ‘poor-quality’ forages by ruminants particularly under tropical conditions. Nutr. Res. Rev. 1990, 3, 277–303. [Google Scholar] [CrossRef] [PubMed]
- Dove, H.; Mayes, R.W. Using n-alkanes and other plant wax components to estimate intake, digestibility and diet composition of grazing/browsing sheep and goats. Small Rumin. Res. 2005, 59, 123–139. [Google Scholar] [CrossRef]
- Gordon, I.J. Animal-based techniques for grazing ecology research. Small Rumin. Res. 1995, 16, 203–214. [Google Scholar] [CrossRef]
- Dove, H. Pasture and grazing animals—The interaction continues. Anim. Prod. Aust. 1998, 22, 3–13. [Google Scholar]
- Bowen, M.K.; Chudleigh, F.; Buck, S.; Hopkins, K. Productivity and profitability of forage options for beef production in the subtropics of northern Australia. Anim. Prod. Sci. 2018, 58, 332–342. [Google Scholar] [CrossRef]
- White, I.A.; Hunt, L.P.; Poppi, D.P.; Petty, S.R. Sampling requirements for predicting cattle diet quality using faecal near-infrared reflectance spectroscopy (F.NIRS) in heterogeneous tropical rangeland pastures. Rangel. J. 2010, 32, 435–441. [Google Scholar] [CrossRef]
- Dixon, R.M.; Coates, D.B. The use of faecal NIRS to improve nutritional management of cattle in northern Australia. Recent Adv. Anim. Nutr. Aust. 2005, 15, 65–75. [Google Scholar]
- Orr, D.; Evenson, C.; Jordan, D.; Bowly, P.; Lehane, K.; Cowan, D. Sheep productivity in an Astrebla grassland of south-west Queensland. Rangel. J. 1988, 10, 39–47. [Google Scholar] [CrossRef]
- Holm, A.; Eliot, G. Seasonal changes in the nutritive value of some native pasture species in North-western Australia. Rangel. J. 1980, 2, 175–182. [Google Scholar] [CrossRef]
- Hall, T. The nitrogen and phosphorus concentrations of some pasture species in the Dichanthium-Eulalia Grasslands of North-West Queensland. Rangel. J. 1981, 3, 67. [Google Scholar] [CrossRef]
- Hersom, M.J.; Wettemann, R.P.; Krehbiel, C.R.; Horn, G.W.; Keisler, D.H. Effect of live weight gain of steers during winter grazing: III. Blood metabolites and hormones during feedlot finishing. J. Anim. Sci. 2004, 82, 2059–2068. [Google Scholar] [CrossRef]
- Murillo, M.; Herrera, E.; Ruiz, O.; Reyes, O.; Carrete, F.O.; Gutierrez, H. Effect of supplemental corn dried distillers grains with solubles fed to beef steers grazing native rangeland during the forage dormant season. Asian Australas. J. Anim. Sci. 2015, 29, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, J.J. Carbohydrate Metabolism and Its Diseases, 5th ed.; Kaneko, J., Harvey, J., Bruss, M., Eds.; Elsevier: San Diego, CA, USA, 1997; ISBN 978-0-12-396305-5. [Google Scholar]
- Mitruka, B.M.; Rawnsley, H.M. Clinical Biochemical and Hematological Reference Values in Normal Experimental Animals; Masson Publishing USA Inc.: New York, USA, 1977; ISBN 9780893520663. [Google Scholar]
- Asano, K.; Ishida, M.; Ishida, M. Effects of inclusion levels of pelleted silvergrass (Miscanthus sinensis Andress.) in the diet on digestibility, chewing activity, ruminal fermentation and blood metabolites in breeding Japanese Black cows. Anim. Sci. J. 2017, 88, 468–475. [Google Scholar] [CrossRef]
- Beeby, J.M.; Haresign, W.; Swan, H. Endogenous hormone and metabolite concentrations in different breeds of beef steer on two systems of production. Anim. Prod. 1988, 47, 231–244. [Google Scholar] [CrossRef]
- Benedet, A.; Manuelian, C.L.; Zidi, A.; Penasa, M.; De Marchi, M. Invited review: β-hydroxybutyrate concentration in blood and milk and its associations with cow performance. Animal 2019, 13, 1676–1689. [Google Scholar] [CrossRef] [PubMed]
- Braun, J.-P.; Lefebvre, H.P. Kidney function and damage. In Clinical Biochemistry of Domestic Animals; Kaneko, J.J., Harvey, J.W., Bruss, M.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 485–528. ISBN 9780123704917. [Google Scholar]
- Grünwaldt, E.G.; Guevara, J.C.; Estévez, O.R.; Vicente, A.; Rousselle, H.; Alcuten, N.; Aguerregaray, D.; Stasi, C.R. Biochemical and haematological measurements in beef cattle in Mendoza plain rangelands (Argentina). Trop. Anim. Health Prod. 2005, 37, 527–540. [Google Scholar] [CrossRef]
- Otto, F.; Vilela, F.; Harun, M.; Taylor, G.; Baggasse, P.; Bogin, E. Biochemical blood profile of Angoni catle in Mozambique. Isr. J. Vet. Med. 2000, 55, 1–9. [Google Scholar]
- Agenas, S.; Heath, M.; Nixon, R.; Wilkinson, J.; Phillips, C. Indicators of undernutrition in cattle. Anim. Welf. 2006, 15, 149–160. [Google Scholar]
- Issi, M.; Gül, Y.; Başbuğ, O. Evaluation of renal and hepatic functions in cattle with subclinical and clinical ketosis. Turk. J. Vet. Anim. Sci. 2016, 40, 47–52. [Google Scholar] [CrossRef]
- Ngo, T. The Effects of Diet Preference on Feed Intake, Digestibility and Nitrogen Balance of Sheep Given Flinders Grass (Iseilema spp.) Hay and/or Desmanthus Leptophyllus ad Libitum. Master’s Thesis, James Cook University, Townsville, Australia, 2017. [Google Scholar]
- Aoetpah, A.; Gardiner, C.; Gummow, B.; Walker, G. Growth and eye muscle area of cross-bred Boer goats fed Desmanthus cultivar JCU 1 hay. In Proceedings of the 32nd Biennial Conference of the Australian Society of Animal Production, Wagga Wagga, Australia, 2–4 July 2018. [Google Scholar]
- Radrizzani, A.; Nasca, J.A. The effect of Leucaena leucocephala on beef production and its toxicity in the Chaco Region of Argentina. Trop. Grassl.-Forrajes Trop. 2014, 2, 127–129. [Google Scholar] [CrossRef]
- Castro-Montoya, J.; Dickhoefer, U. Effects of tropical legume silages on intake, digestibility and performance in large and small ruminants: A review. Grass Forage Sci. 2018, 73, 26–39. [Google Scholar] [CrossRef]
- Cheffins, R. Nutritional and Managerial Opportunities for Meeting Beef Markets. Information Series QI 96112; Department of Primary Industries Queensland Meat Research Corporation: Brisbane, Australia, 1996; ISBN 0724266909 (pbk.).
- Rubanza, C.D.K.; Shem, M.N.; Otsyina, R.; Fujihara, T. Performance of Zebu steers grazing on western Tanzania native forages supplemented with Leucaena leucocephala leaf meal. Agrofor. Syst. 2005, 65, 165–174. [Google Scholar] [CrossRef]
- Pen, M.; Savage, D.B.; Nolan, J.V.; Seng, M. Effect of Stylosanthes guianensis supplementation on intake and nitrogen metabolism of Bos indicus cattle offered a basal diet of mixed rice straw and tropical grass. Anim. Prod. Sci. 2013, 53, 453–457. [Google Scholar] [CrossRef]
- Bowman, J.G.P.; Sowell, B.F.; Paterson, J.A. Liquid supplementation for ruminants fed low-quality forage diets: A review. Anim. Feed Sci. Technol. 1995, 55, 105–138. [Google Scholar] [CrossRef]
- Detmann, E.; Valente, É.E.L.; Batista, E.D.; Huhtanen, P. An evaluation of the performance and efficiency of nitrogen utilization in cattle fed tropical grass pastures with supplementation. Livestig. Sci. 2014, 162, 141–153. [Google Scholar] [CrossRef]
- McCown, R.L.; McLean, R.W. An analysis of cattle live-weight changes on tropical grass pasture during the dry and early wet seasons in northern Australia: 2. Relations to trends in the pasture, diet and grazing behaviour. J. Agric. Sci. 1983, 101, 17–24. [Google Scholar] [CrossRef]
- Schatz, T.; Ffoulkes, D.; Shotton, P.; Hearnden, M. Effect of high-intensity rotational grazing on the growth of cattle grazing buffel pasture in the Northern Territory and on soil carbon sequestration. Anim. Prod. Sci. 2020, 1814–1821. [Google Scholar] [CrossRef]
- Dixon, R.M.; Shotton, P.; Mayer, R. Diets selected and growth of steers grazing buffel grass (Cenchrus ciliaris cv. Gayndah)-Centro (Centrosema brasilianum cv. Oolloo) pastures in a seasonally dry tropical environment. Anim. Prod. Sci. 2020, 60, 1459–1468. [Google Scholar] [CrossRef]
- Ash, A.J.; McIvor, J.G.; Corfield, J.P.; Winter, W.H. How land condition alters plant-animal relationships in Australia’s tropical rangelands. Agric. Ecosyst. Environ. 1995, 56, 77–92. [Google Scholar] [CrossRef]
- Clements, R.J. Pastures for prosperity. 3. The future for new tropical pasture plants. Trop. Grassl. 1996, 30, 31–46. [Google Scholar]
- Brandão, R.K.C.; de Carvalho, G.; Silva, R.; Dias, D.; Mendes, F.; Lins, T.; Pereira, M.; Guimarães, J.; Tosto, M.; Rufino, L.; et al. Correlation between production performance and feeding behavior of steers on pasture during the rainy-dry transition period. Trop. Anim. Health Prod. 2018, 50, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Drouillard, J.S.; Kuhl, G.L. Effects of previous grazing nutrition and management on feedlot performance of cattle. J. Anim. Sci. 1999, 77, 136–146. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Year | Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec | Annual Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2017 | 68 | 39 | 105 | 6 | 8 | 0 | 22 | 0 | 0 | 107 | 42 | 24 | 421 |
| 2018 | 19 | 118 | 36 | 6 | 0 | 24 | 4 | 10 | 4 | 68 | 41 | 38 | 368 |
| 2019 | 0 | 1 | 111 | 82 | 3 | 5 | 24 | 15 | 0 | 40 | 20 | 5 | 306 |
| Variable | Buffel Grass Paddock | Desmanthus Paddock | ||
|---|---|---|---|---|
| Pre-Grazing | End of Grazing | Pre-Grazing | End of Grazing | |
| Ground cover | 63.7 | 38.0 | 68.7 | 29.7 |
| Woody Cover | 0.7 | 0.7 | 0.5 | 0.7 |
| Dry matter yield (kg/ha) | ||||
| Total yield | 4066 | 1854 | 4509 | 1425 |
| Cenchrus ciliaris | 3532 | 1700 | 3372 | 1260 |
| Desmanthus spp. | 502 | 88.6 | ||
| Salsola kali | 8.0 | 4.2 | 131 | 3.9 |
| Urochloa mosambicensis | 158 | 57.2 | 112 | 28.1 |
| Portulaca spp. | 18.0 | 5.5 | 80.0 | 2.1 |
| Botanical composition | ||||
| Cenchrus ciliaris | 90.1 | 91.7 | 77.2 | 88.4 |
| Desmanthus spp. | 11.5 | 6.2 | ||
| Salsola kali | 0.2 | 0.3 | 3.0 | 0.3 |
| Urochloa mosambicensis | 4.0 | 3.1 | 2.6 | 2.0 |
| Portulaca spp. | 0.5 | 0.3 | 1.8 | 0.2 |
| Variable | Buffel Grass | Desmanthus | Brigalow |
|---|---|---|---|
| DM (%) | 84.9 ± 3.1 | 68.3 ± 3.4 | 64.6 ± 1.6 |
| Neutral detergent fibre | 73.9 ± 1.0 | 62.8 ± 2.0 | 38.7 ± 0.5 |
| Acid detergent fibre | 43.4 ± 1.1 | 40.9 ± 1.6 | 25.5 ± 1.5 |
| Dry matter digestibility | 46.9 ± 1.1 | 48.4 ± 1.2 | 60.6 ± 1.1 |
| Organic matter | 93.1 ± 0.3 | 94.6 ± 0.5 | 91.8 ± 0.3 |
| Ash | 7.2 ± 0.2 | 5.4 ± 0.5 | 8.2 ± 0.3 |
| Hemicellulose | 30.5 ± 1.3 | 21.9 ± 2.0 | 13.2 ± 2.0 |
| Crude Protein | 4.4 ± 0.9 | 8.5 ± 1.4 | 7.5 ± 0.3 |
| Metabolizable energy (Mj·kg−1 DM) 1 | 6.9 ± 0.1 | 6.8 ± 0.128 | 8.7 ± 0.2 |
| Variable | Paddock | Days Since the Onset of Grazing | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 49 | 79 | 147 | P | D | P*D | |||
| Diet CP (%) | Buffel grass | 11.26 ab | 11.70 a | 10.99 ab | 10.92 ab | 0.995 | 0.001 | 0.001 | 0.001 |
| Desmanthus | 11.28 ab | 8.78 d | 10.61 bc | 9.94 c | |||||
| Faecal N (%) | Buffel grass | 1.75 a | 1.75 a | 1.73 a | 1.56 b | 0.150 | 0.68 | 0.001 | 0.14 |
| Desmanthus | 1.74 a | 1.73 a | 1.79 a | 1.49 b | |||||
| DMD (%) | Buffel grass | 53.94 c | 55.11 ab | 55.07 ab | 55.92 a | 1.24 | 0.001 | 0.001 | 0.001 |
| Desmanthus | 54.16 bc | 52.09 d | 54.72 bc | 53.83 c | |||||
| ME (MJ/Kg DM) 1 | Buffel grass | 7.57 c | 7.77 ab | 7.76 ab | 7.91 a | 0.0381 | 0.001 | 0.001 | 0.001 |
| Desmanthus | 7.61 bc | 7.25 d | 7.71 bc | 7.55 bc | |||||
| DNG (%) | Buffel grass | 32.40 a | 28.09 bc | 26.27 c | 19.94 d | 4.91 | 0.65 | 0.001 | 0.089 |
| Desmanthus | 31.87 ab | 27.72 c | 28.52 abc | 17.37 d | |||||
| Metabolite | Pasture | Sampling Period | SEM | p-Value | |||
|---|---|---|---|---|---|---|---|
| Day 0 | Day 147 | P | D | P*D | |||
| Total Bilirubin (µmol/L) | Buffel grass | 2.93 | 2.29 | 1.21 | 0.41 | 0.041 | 0.094 |
| Desmanthus | 2.81 | 2.30 | |||||
| BHB (mmol/L) | Buffel grass | 0.22 | 0.28 | 0.0603 | 0.35 | 0.001 | 0.68 |
| Desmanthus | 0.21 | 0.25 | |||||
| Creatinine (µmol/L) | Buffel grass | 94.48 | 109.15 | 16.27 | 0.12 | 0.001 | 0.011 |
| Desmanthus | 89.25 | 122.70 | |||||
| NEFA (mmol/L) | Buffel grass | 0.45 | 0.36 | 0.177 | 0.058 | 0.21 | 0.31 |
| Desmanthus | 0.36 | 0.33 | |||||
| Glucose (mmol/L) | Buffel grass | 5.9 | 4.8 | 1.00 | 0.46 | 0.001 | 0.40 |
| Desmanthus | 5.7 | 4.8 | |||||
| Variable | Pasture | Days Since the Onset of Grazing | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 49 | 79 | 147 | P | Days | P*D | |||
| LW (kg) | Buffel grass | 319 d | 372 c | 392 b | 431 a | 18.9 | 0.14 | 0.001 | 0.21 |
| Desmanthus | 322 d | 369 c | 396 b | 433 a | |||||
| BCS | Buffel grass | 3.46 c | 3.60 bc | 3.58 bc | 4.10 a | 0.38 | 0.51 | 0.001 | 0.36 |
| Desmanthus | 3.46 c | 3.65 bc | 3.59 bc | 3.90 ab | |||||
| Overall ADG (kg/day) | Buffel grass | 0.74 | 0.13 | 0.78 | |||||
| Desmanthus | 0.75 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mwangi, F.W.; Gardiner, C.P.; Walker, G.; Hall, T.J.; Malau-Aduli, B.S.; Kinobe, R.T.; Malau-Aduli, A.E.O. Growth Performance and Plasma Metabolites of Grazing Beef Cattle Backgrounded on Buffel or Buffel-Desmanthus Mixed Pastures. Animals 2021, 11, 2355. https://doi.org/10.3390/ani11082355
Mwangi FW, Gardiner CP, Walker G, Hall TJ, Malau-Aduli BS, Kinobe RT, Malau-Aduli AEO. Growth Performance and Plasma Metabolites of Grazing Beef Cattle Backgrounded on Buffel or Buffel-Desmanthus Mixed Pastures. Animals. 2021; 11(8):2355. https://doi.org/10.3390/ani11082355
Chicago/Turabian StyleMwangi, Felista W., Christopher P. Gardiner, Glen Walker, Trevor J. Hall, Bunmi S. Malau-Aduli, Robert T. Kinobe, and Aduli E. O. Malau-Aduli. 2021. "Growth Performance and Plasma Metabolites of Grazing Beef Cattle Backgrounded on Buffel or Buffel-Desmanthus Mixed Pastures" Animals 11, no. 8: 2355. https://doi.org/10.3390/ani11082355
APA StyleMwangi, F. W., Gardiner, C. P., Walker, G., Hall, T. J., Malau-Aduli, B. S., Kinobe, R. T., & Malau-Aduli, A. E. O. (2021). Growth Performance and Plasma Metabolites of Grazing Beef Cattle Backgrounded on Buffel or Buffel-Desmanthus Mixed Pastures. Animals, 11(8), 2355. https://doi.org/10.3390/ani11082355