
Dietary Yeast Cell Wall Improves Growth Performance and Prevents of Diarrhea of Weaned Pigs by Enhancing Gut Health and Anti-Inflammatory Immune Responses

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sample Collection
2.2. Diets
2.3. Growth Performance
2.4. Nutrient Digestibility
2.5. Intestinal Morphology
2.6. Immune Responses
2.7. Gene Expression Profiles in Ileum
2.8. 16s rRNA Sequencing for Fecal Microbiota
2.9. Statistical Analyses
3. Results
3.1. Growth Performance
3.2. Nutrient Digestibility and Intestinal Morphology
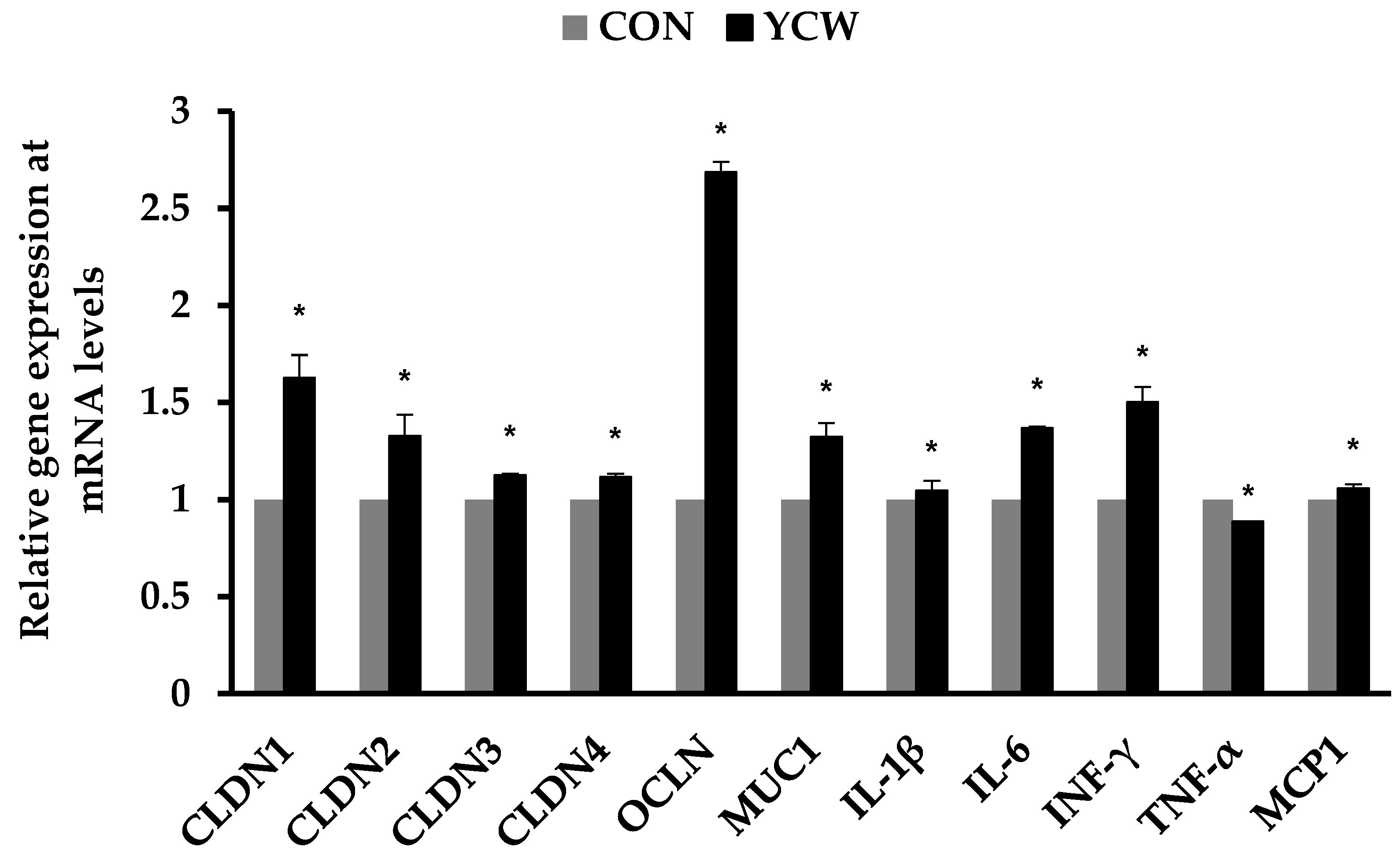
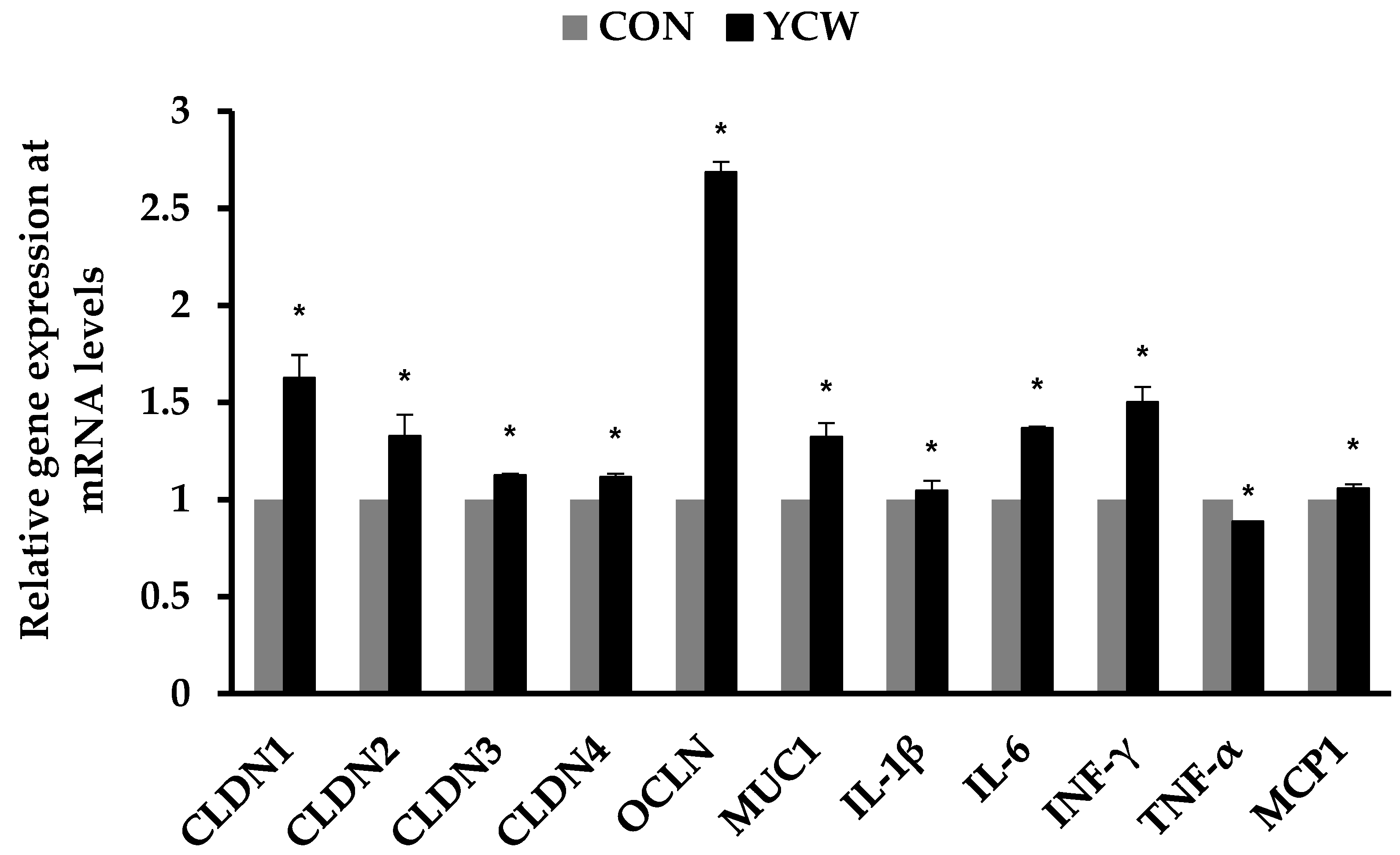
3.3. Immune Responses and Gene Expression of Tight Junction Proteins
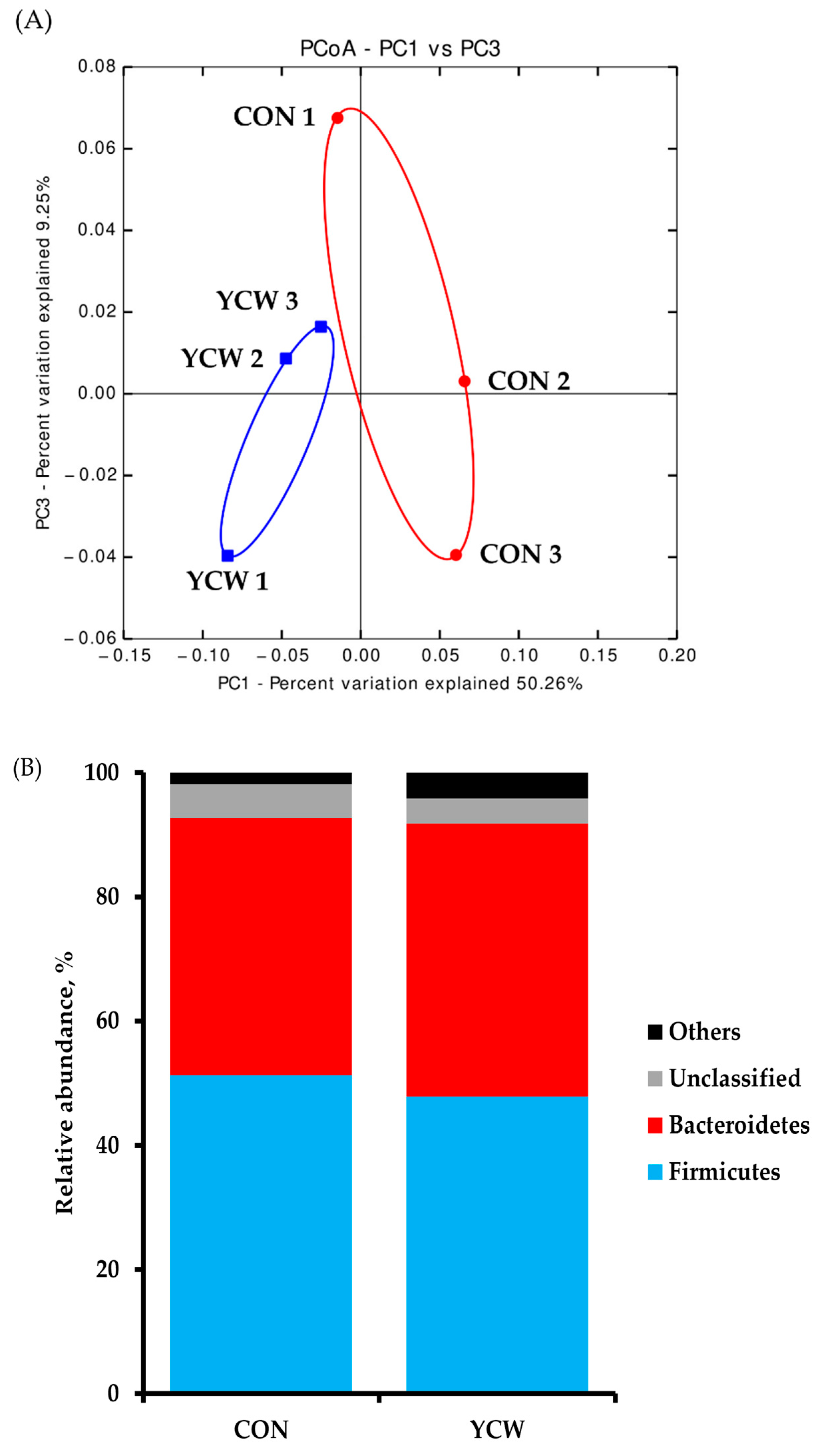
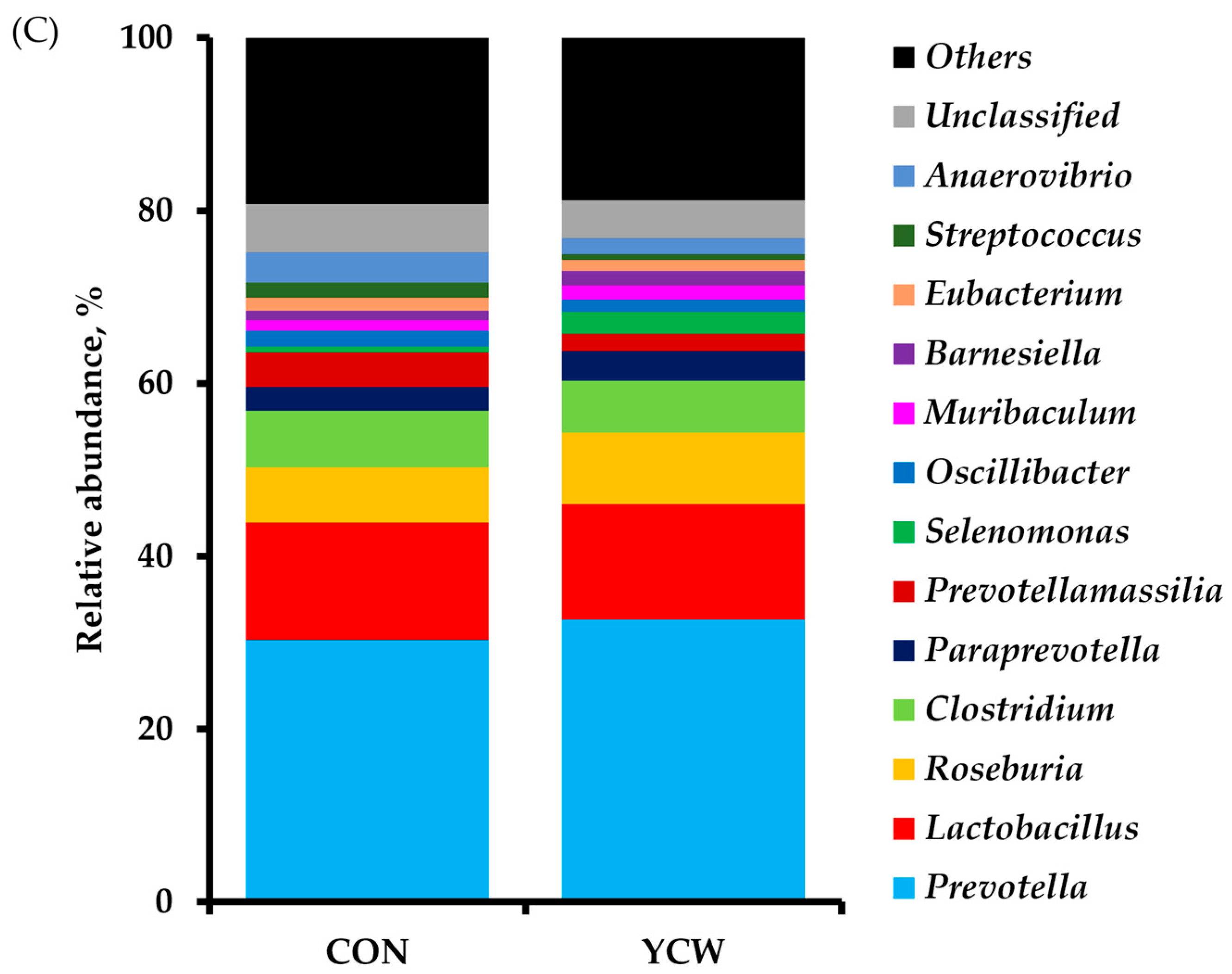
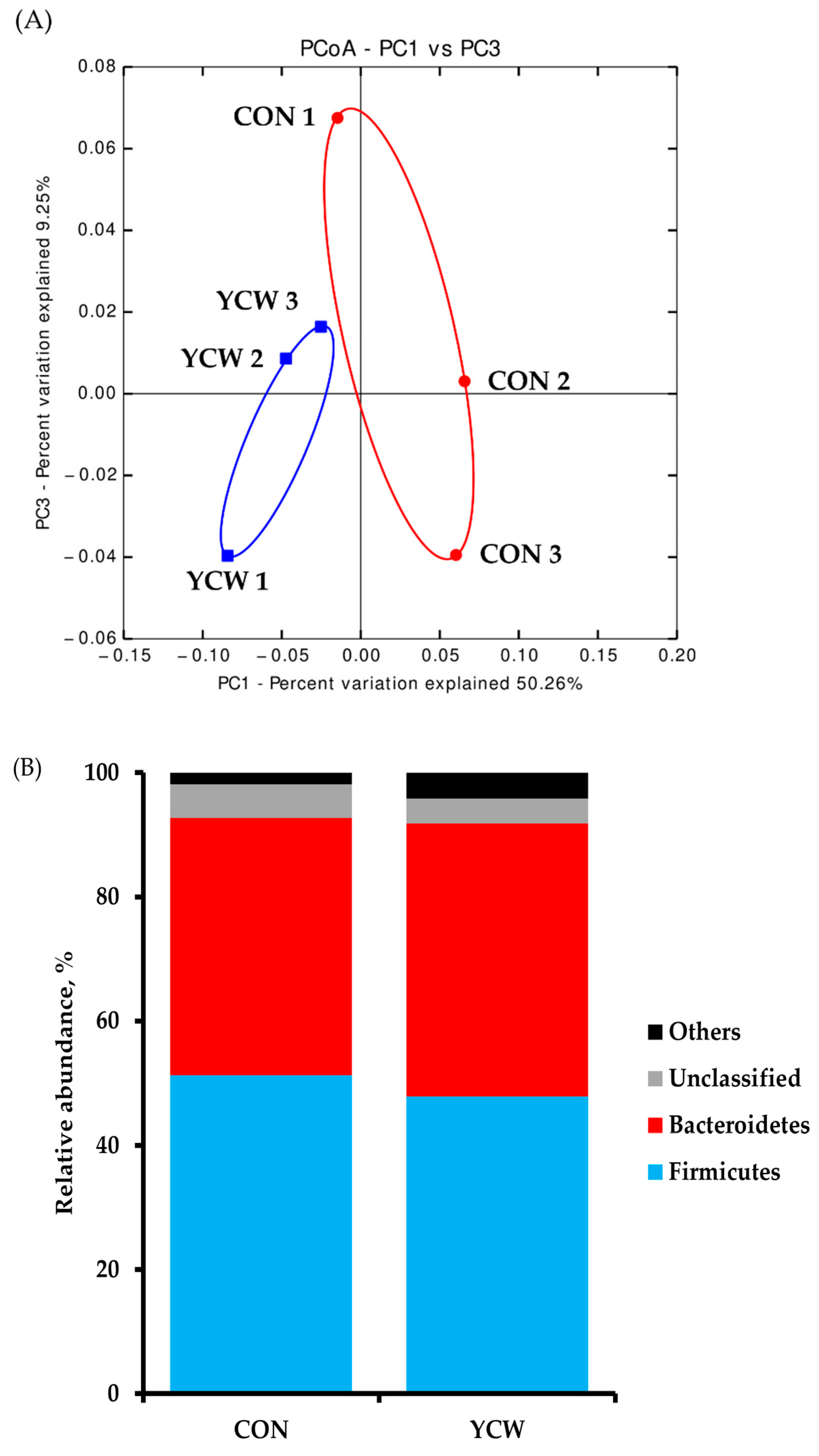
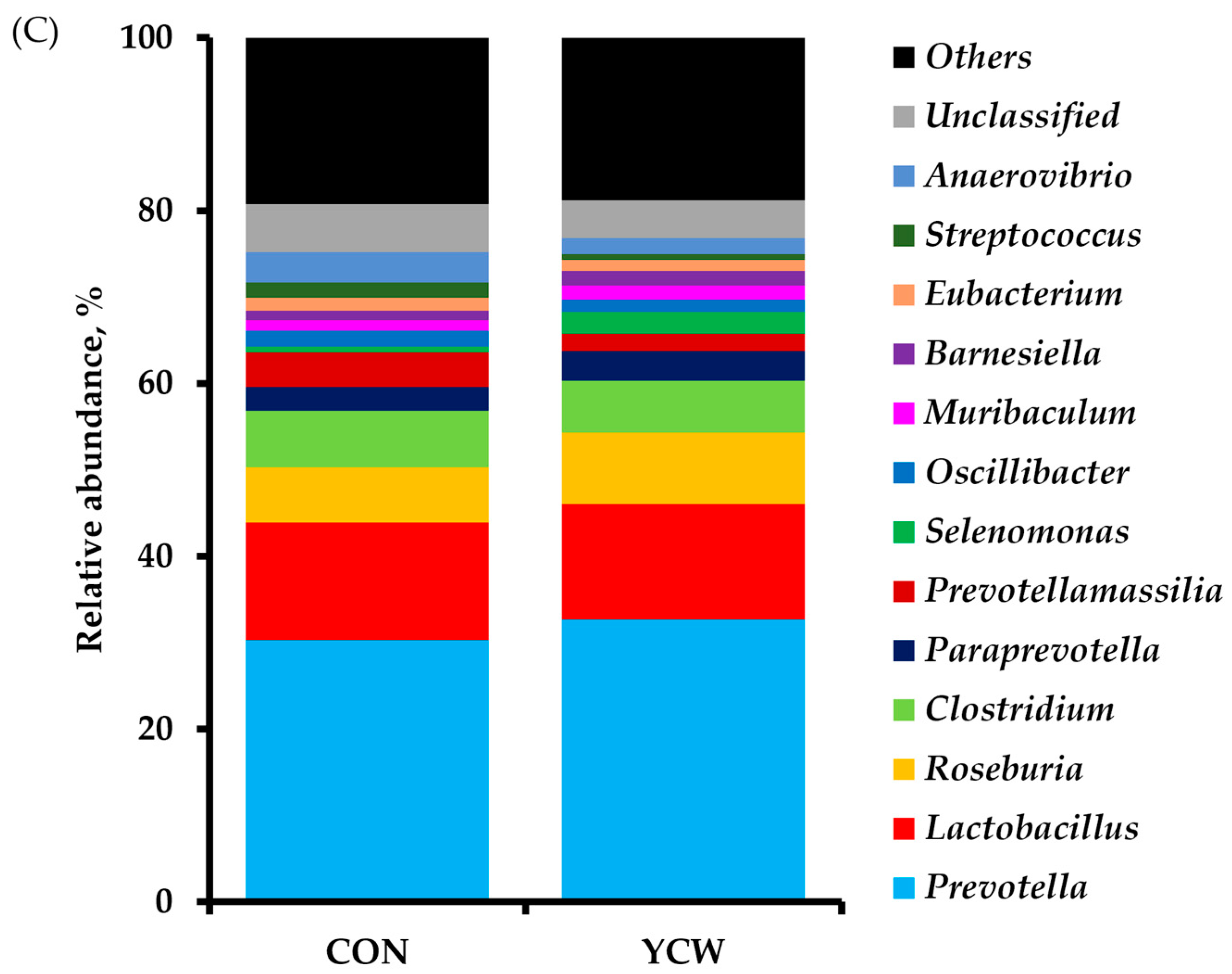
3.4. Diversity of Intestinal Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Madec, F.; Bridoux, N.; Bounaix, S.; Jestin, A. Measurement of digestive disorders in the piglet at weaning and related risk factors. Prev. Vet. Med. 1998, 35, 53–72. [Google Scholar] [CrossRef]
- Lalles, J.P.; Boudry, G.; Favier, C.; Le Floc’h, N.; Lurona, I.; Montagne, L.; Oswald, I.P.; Pie, S.; Piel, C.; Seve, B. Gut function and dysfunction in young pigs: Physiology. Anim. Res. 2004, 53, 301–316. [Google Scholar] [CrossRef] [Green Version]
- Smith, F.; Clark, J.E.; Overman, B.L.; Tozel, C.C.; Huang, J.H.; Rivier, J.E.F.; Blisklager, A.T.; Moeser, A.J. Early weaning stress impairs development of mucosal barrier function in the porcine intestine. Am. J. Physiol.-Gastroint. Liver Physiol. 2010, 298, 352–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trckova, M.; Faldyna, M.; Alexa, P.; Zajacova, Z.S.; Gopfert, E.; Kumprechtova, D.; Auclair, E.; D’Inca, R. The effects of live yeast Saccharomyces cerevisiae on postweaning diarrhea, immune response, and growth performance in weaned piglets. J. Anim. Sci. 2014, 92, 767–774. [Google Scholar] [CrossRef]
- Che, L.Q.; Xu, Q.; Wu, C.; Luo, Y.H.; Huang, X.B.; Zhang, B.; Auclair, E.; Kiros, T.; Fang, Z.F.; Lin, Y.; et al. Effects of dietary live yeast supplementation on growth performance, diarrhoea severity, intestinal permeability and immunological parameters of weaned piglets challenged with enterotoxigenic Escherichia coli K88. Br. J. Nutr. 2017, 118, 949–958. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.H.; Fleet, G.H.; Rogers, P.L. Composition of the cell walls of several yeast species. Appl. Microbiol. Biotechnol. 1998, 50, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.B.; Piao, X.S.; Kim, S.W.; Wang, L.; Liu, P.; Yoon, I.; Zhen, Y.G. Effects of yeast culture supplementation on growth performance, intestinal health, and immune response of nursery pigs. J. Anim. Sci. 2009, 87, 2614–2624. [Google Scholar] [CrossRef]
- Pourabedin, M.; Zhao, X. Prebiotics and gut microbiota in chickens. FEMS Microbiol. Lett. 2015, 362, fnv122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bontempo, V.; Di Giancamillo, A.; Savoini, G.; Dell’Orto, V.; Domeneghini, C. Live yeast dietary supplementation acts upon intestinal morpho-functional aspects and growth in weanling piglets. Anim. Feed Sci. Technol. 2006, 129, 224–236. [Google Scholar] [CrossRef]
- Kogan, G.; Kocher, A. Role of yeast cell wall polysaccharides in pig nutrition and health protection. Livest. Sci. 2007, 109, 161–165. [Google Scholar] [CrossRef]
- Molist, F.; van Eerden, E.; Parmentier, H.K.; Vuorenmaa, J. Effects of inclusion of hydrolyzed yeast on the immune response and performance of piglets after weaning. Anim. Feed Sci. Technol. 2014, 195, 136–141. [Google Scholar] [CrossRef]
- Park, S.; Lee, J.J.; Yang, B.M.; Cho, J.H.; Kim, S.; Kang, J.; Oh, S.; Park, D.-J.; Perez-Maldonado, R.; Cho, J.-Y.; et al. Dietary protease improves growth performance, nutrient digestibility, and intestinal morphology of weaned pigs. J. Anim. Sci. Technol. 2020, 62, 21–30. [Google Scholar] [CrossRef] [Green Version]
- NRC. Nutrient Requirements of Swine, 11th ed.; The National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- AOAC. International Official Methods of Analysis, 21st ed.; AOAC International: Gaithersburg, MD, USA, 2019. [Google Scholar]
- Williams, C.H.; David, D.J.; Iismaa, O. The determination of chromic oxide in faeces samples by atomic absorption spectrophotometry. J. Agric. Sci. 1962, 59, 381–385. [Google Scholar] [CrossRef]
- Stein, H.H.; Seve, B.; Fuller, M.F.; Moughan, P.J.; de Lange, C.F.M. Invited review: Amino acid bioavailability and digestibility in pig feed ingredients: Terminology and application. J. Anim. Sci. 2007, 85, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Liu, Y.; Lee, J.J.; Che, T.M.; Soares-Almeida, J.A.; Chun, J.L.; Campbell, J.M.; Polo, J.; Crenshaw, J.D.; Seo, S.W.; et al. Spray-dried plasma attenuates inflammation and improves pregnancy rate of mated female mice. J. Anim. Sci. 2015, 93, 298–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuijk, E.W.; du Puy, L.; van Tol, H.T.A.; Haagsman, H.P.; Colenbrander, B.; Roelen, B.A.J. Validation of reference genes for quantitative RT-PCR studies in porcine oocytes and preimplantation embryos. BMC Dev. Biol. 2007, 7, 58. [Google Scholar] [CrossRef] [Green Version]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a Dual-Index Sequencing Strategy and Curation Pipeline for Analyzing Amplicon Sequence Data on the MiSeq Illumina Sequencing Platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.J.; Kang, J.; Park, S.; Cho, J.H.; Oh, S.; Park, D.-J.; Perez-Maldonado, R.; Cho, J.-Y.; Park, I.-H.; Kim, H.B.; et al. Effects of dietary protease on immune responses of weaned pigs. J. Anim. Sci. Technol. 2020, 62, 174–179. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Kim, I.H. Effects of Saccharomyces cerevisiae cell wall extract and poplar propolis ethanol extract supplementation on growth performance, digestibility, blood profile, fecal microbiota and fecal noxious gas emissions in growing pigs. Anim. Sci. J. 2014, 85, 698–705. [Google Scholar] [CrossRef]
- Lu, H.; Wilcock, P.; Adeola, O.; Ajuwon, K.M. Effect of live yeast supplementation to gestating sows and nursery piglets on postweaning growth performance and nutrient digestibility. J. Anim. Sci. 2019, 97, 2534–2540. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Yang, H.S.; Li, B.; Liu, G.; Huang, R.L.; Li, F.N.; Liao, P.; Zhang, Y.Z.; Nyachoti, C.M.; Deng, D. Dietary supplementation with yeast product improves intestinal function, and serum and ileal amino acid contents in weaned piglets. Livest. Sci. 2015, 171, 20–27. [Google Scholar] [CrossRef]
- Waititu, S.M.; Yin, F.; Patterson, R.; Yitbareka, A.; Rodriguez-Lecompte, J.C.; Nyachoti, C.M. Dietary supplementation with a nucleotide-rich yeast extract modulates gut immune response and microflora in weaned pigs in response to a sanitary challenge. Animal 2017, 11, 2156–2164. [Google Scholar] [CrossRef]
- Al-Sadi, R.; Khatib, K.; Guo, S.H.; Ye, D.M.; Youssef, M.; Ma, T. Occludin regulates macromolecule flux across the intestinal epithelial tight junction barrier. Am. J. Physiol.-Gastroint. Liver Physiol. 2011, 300, 1054–1064. [Google Scholar] [CrossRef] [Green Version]
- Sivignon, A.; de Vallee, A.; Barnich, N.; Denizot, J.; Darcha, C.; Pignede, G.; Vandekerckove, P.; Darfeuille-Michaud, A. Saccharomyces cerevisiae CNCM I-3856 prevents colitis Induced by AIEC bacteria in the transgenic mouse model mimicking crohn’s disease. Inflamm. Bowel Dis. 2015, 21, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Ji, W.; Wang, J.; Li, B.; Hu, J.; Wu, X. Effects of dietary supplementation with yeast glycoprotein on growth performance, intestinal mucosal morphology, immune response and colonic microbiota in weaned piglets. Food Funct. 2019, 10, 2359–2371. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, Y.; Yang, Z.; Li, C.; Liang, H.; Wu, Z.; Pu, W. Yeast probiotics shape the gut microbiome and improve the health of early-weaned piglets. Front. Microbiol. 2018, 9, 2011. [Google Scholar] [CrossRef]
- Looft, T.; Johnson, T.A.; Allen, H.K.; Bayles, D.O.; Alt, D.P.; Stedtfeld, R.D.; Sul, W.J.; Stedtfeld, T.M.; Chai, B.; Cole, J.R.; et al. In-feed antibiotic effects on the swine intestinal microbiome. Proc. Natl. Acad. Sci. USA 2012, 109, 1691–1696. [Google Scholar] [CrossRef] [Green Version]
- Tamanai-Shacoori, Z.; Smida, I.; Bousarghin, L.; Loreal, O.; Meuric, V.; Fong, S.B.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Roseburia spp.: A marker of health? Future Microbiol. 2017, 12, 157–170. [Google Scholar] [CrossRef]
- Lee, J.J.; Choi, S.H.; Cho, J.H.; Choe, J.; Kang, J.; Kim, S.; Park, S.; Kyoung, H.; Seo, D.; Cho, J.-Y.; et al. Effects of dietary carbohydrases on productive performance and immune responses of lactating sows and their piglets. J. Anim. Sci. Technol. 2019, 61, 359–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yimin; Kohanawa, M. A Regulatory Effect of the Balance between TNF-α and IL-6 in the Granulomatous and Inflammatory Response to Rhodococcus aurantiacus Infection in Mice. J. Immunol. 2006, 177, 642–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.S.; Hosseindoust, A.; Lee, S.H.; Choi, Y.H.; Kim, M.J.; Lee, J.H.; Kwon, I.K.; Chae, B.J. Bacteriophage cocktail and multi-strain probiotics in the feed for weanling pigs: Effects on intestine morphology and targeted intestinal coliforms and clostridium. Animal 2017, 11, 45–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| Items | Basal Diet |
|---|---|
| Ingredient, % | |
| Corn | 53.90 |
| Soybean meal (44%) | 15.00 |
| Soy protein concentrate | 7.50 |
| Whey powder | 12.50 |
| Soybean oil | 2.30 |
| Spray dried porcine plasma | 2.50 |
| Fish meal, combined | 3.00 |
| Limestone | 1.20 |
| Monocalcium phosphate | 0.80 |
| Vitamin-mineral premix 1 | 0.40 |
| L-Lysine-HCl | 0.35 |
| DL-Methionine | 0.15 |
| L-Threonine | 0.10 |
| Zinc oxide | 0.30 |
| Total | 100.00 |
| Calculated energy and nutrient contents | |
| Metabolizable energy, MJ/kg | 14.24 |
| Crude protein, % | 21.69 |
| Calcium, % | 0.89 |
| Phosphorus, % | 0.68 |
| Lysine, % | 1.55 |
| Items 1 | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| CLDN1 | AGAAGATGCGGATGGCTGTC | CCCAGAAGGCAGAGAGAAGC |
| CLDN2 | TCCTCCCTGTTCTCCCTGATAG | CCTTGCAGTGGGCAGGAA |
| CLDN3 | GATGCAGTGCAAAGTGTACGA | GTCCTGCACGCAGTTGGT |
| CLDN4 | TATCATCCTGGCCGTGCTA | CATCATCCACGCAGTTGGT |
| OCLN | GGAGTGATTCGGATTCTGTCTATGCT | CGCCTGGGCTGTTGGGTTGA |
| MUC1 | CCCTGGCCATCATCTATGTC | TGCCCACAGTTCTTTCGTC |
| IL-1β | GCCCTGTACCCCAACTGGTA | CCCAGGAAGACGGGCTTT |
| IL-6 | GCGCAGCCTTGAGGATTTC | CCCAGCTACATTATCCGAATGG |
| INF-γ | GAGCCAAATTGTCTCCTTCTAC | CGAAGTCATTCAGTTTCCCAG |
| TNF-α | CTTGGGTTTGGATTCCTGGAT | CTTCCCTGGCAGCCACAT |
| MCP1 | TCCCACACCGAAGCTTGAAT | CACAGGAGGGCTGCAGAGA |
| 18S rRNA | GGCTACCACATCCAAGGAAG | TCCAATGGATCCTCGCGGAA |
| Item 1 | CON | YCW | SEM | p-Value |
|---|---|---|---|---|
| Day 1 to 7 | ||||
| Initial BW, kg | 7.99 | 7.98 | 0.43 | 0.995 |
| Final BW, kg | 9.21 | 9.33 | 0.40 | 0.842 |
| ADG, g/d | 203.93 | 223.69 | 15.81 | 0.394 |
| ADFI, g/d | 258.44 | 278.57 | 13.93 | 0.327 |
| G:F, g/g | 0.78 | 0.80 | 0.03 | 0.710 |
| Diarrhea 2, % | 6.12 | 0.00 | 0.243 | |
| Day 8 to 14 | ||||
| Initial BW, kg | 9.21 | 9.33 | 0.40 | 0.842 |
| Final BW, kg | 12.60 | 12.94 | 0.45 | 0.598 |
| ADG, g/d | 376.19 | 401.59 | 11.96 | 0.159 |
| ADFI, g/d | 512.31 | 545.45 | 14.10 | 0.123 |
| G:F, g/g | 0.73 | 0.74 | 0.01 | 0.882 |
| Diarrhea 2, % | 18.37 | 10.20 | 0.317 | |
| Day 1 to 14 | ||||
| Initial BW, kg | 7.99 | 7.98 | 0.43 | 0.995 |
| Final BW, kg | 12.60 | 12.94 | 0.45 | 0.598 |
| ADG, g/d | 307.29 | 330.43 | 11.68 | 0.186 |
| ADFI, g/d | 410.76 | 438.70 | 11.82 | 0.121 |
| G:F, g/g | 0.75 | 0.75 | 0.01 | 0.783 |
| Diarrhea 2, % | 12.24 | 5.10 | 0.083 | |
| Day 15 to 28 | ||||
| Initial BW, kg | 12.60 | 12.94 | 0.45 | 0.598 |
| Final BW, kg | 19.77 | 20.53 | 0.65 | 0.422 |
| ADG, g/d | 512.50 | 542.35 | 16.83 | 0.234 |
| ADFI, g/d | 856.03 | 863.88 | 30.49 | 0.859 |
| G:F, g/g | 0.60 | 0.63 | 0.01 | 0.118 |
| Day 1 to 28 | ||||
| Initial BW, kg | 7.99 | 7.98 | 0.43 | 0.995 |
| Final BW, kg | 19.77 | 20.53 | 0.65 | 0.422 |
| ADG, g/d | 420.87 | 448.19 | 10.41 | 0.088 |
| ADFI, g/d | 648.07 | 666.96 | 18.36 | 0.481 |
| G:F, g/g | 0.65 | 0.67 | 0.01 | 0.101 |
| Item 1 | CON | YCW | SEM | p-Value |
|---|---|---|---|---|
| Apparent ileal digestibility, % | ||||
| Dry matter | 79.76 | 80.75 | 0.27 | 0.024 |
| Crude protein | 72.27 | 71.68 | 2.56 | 0.873 |
| Energy | 75.77 | 78.14 | 0.78 | 0.052 |
| Apparent total tract digestibility, % | ||||
| Dry matter | 84.06 | 86.02 | 0.90 | 0.147 |
| Crude protein | 76.17 | 76.11 | 1.02 | 0.972 |
| Energy | 81.61 | 83.46 | 0.69 | 0.085 |
| Duodenum | ||||
| Villus height, μm | 317.73 | 356.68 | 16.12 | 0.113 |
| Crypt depth, μm | 266.48 | 236.14 | 12.31 | 0.107 |
| VH:CD, μm/μm | 1.19 | 1.52 | 0.05 | 0.001 |
| Villus width, μm | 137.83 | 162.61 | 6.76 | 0.024 |
| Villus area, μm2 | 38,286 | 46,226 | 1688 | 0.006 |
| Goblet cell, n | 9.93 | 13.95 | 1.10 | 0.024 |
| Jejunum | ||||
| Villus height, μm | 307.05 | 338.81 | 19.42 | 0.349 |
| Crypt depth, μm | 263.97 | 238.44 | 19.85 | 0.381 |
| VH:CD, μm/μm | 1.19 | 1.42 | 0.08 | 0.077 |
| Villus width, μm | 159.44 | 173.99 | 8.58 | 0.254 |
| Villus area, μm2 | 40,550 | 42,568 | 1491 | 0.357 |
| Goblet cell, n | 9.03 | 13.27 | 1.05 | 0.014 |
| Ileum | ||||
| Villus height, μm | 393.54 | 416.01 | 10.95 | 0.172 |
| Crypt depth, μm | 256.93 | 242.39 | 13.98 | 0.476 |
| VH:CD, μm/μm | 1.55 | 1.74 | 0.09 | 0.153 |
| Villus width, μm | 143.70 | 173.31 | 10.08 | 0.060 |
| Villus area, μm2 | 32,201 | 41,849 | 3691 | 0.089 |
| Goblet cell, n | 14.71 | 14.94 | 0.86 | 0.851 |
| Item 1 | CON | YCW | SEM | p-Value |
|---|---|---|---|---|
| White blood cell, ×103/μL | ||||
| Day 0 | 12.51 | 10.43 | 1.51 | 0.348 |
| Day 7 | 14.90 | 15.31 | 0.93 | 0.757 |
| Day 14 | 21.81 | 16.95 | 1.79 | 0.081 |
| Cortisol, ng/mL | ||||
| Day 0 | 44.73 | 45.40 | 2.03 | 0.821 |
| Day 7 | 48.23 | 46.87 | 1.00 | 0.363 |
| Day 14 | 50.45 | 49.86 | 1.74 | 0.467 |
| Tumor necrosis factor-α, pg/mL | ||||
| Day 0 | 65.84 | 64.12 | 5.65 | 0.836 |
| Day 7 | 126.57 | 81.91 | 13.48 | 0.039 |
| Day 14 | 119.43 | 111.88 | 13.63 | 0.702 |
| Transforming growth factor-β1, pg/mL | ||||
| Day 0 | 1397.69 | 1310.72 | 106.45 | 0.576 |
| Day 7 | 1248.89 | 1200.29 | 95.41 | 0.726 |
| Day 14 | 1129.77 | 1474.58 | 107.88 | 0.050 |
| Interleukin-1β, pg/mL | ||||
| Day 0 | 28.97 | 25.73 | 4.69 | 0.635 |
| Day 7 | 39.29 | 41.06 | 2.10 | 0.564 |
| Day 14 | 38.88 | 27.20 | 2.49 | 0.008 |
| Interleukin-6, pg/mL | ||||
| Day 0 | 139.41 | 140.44 | 2.90 | 0.808 |
| Day 7 | 147.84 | 143.56 | 3.62 | 0.420 |
| Day 14 | 143.12 | 138.80 | 3.90 | 0.448 |
| Item 1 | CON | YCW | p-Value |
|---|---|---|---|
| Number of read sequences | 128,647 ± 759 | 122,242 ± 44,506 | 0.535 |
| OTUs | 286.00 ± 48.82 | 263.30 ± 31.09 | 0.432 |
| Chao1 | 315.80 ± 51.97 | 285.10 ± 31.59 | 0.445 |
| Shannon | 5.33 ± 0.54 | 5.62 ± 0.26 | 0.193 |
| Inverse Simpson | 0.93 ± 0.03 | 0.96 ± 0.01 | 0.826 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.J.; Kyoung, H.; Cho, J.H.; Choe, J.; Kim, Y.; Liu, Y.; Kang, J.; Lee, H.; Kim, H.B.; Song, M. Dietary Yeast Cell Wall Improves Growth Performance and Prevents of Diarrhea of Weaned Pigs by Enhancing Gut Health and Anti-Inflammatory Immune Responses. Animals 2021, 11, 2269. https://doi.org/10.3390/ani11082269
Lee JJ, Kyoung H, Cho JH, Choe J, Kim Y, Liu Y, Kang J, Lee H, Kim HB, Song M. Dietary Yeast Cell Wall Improves Growth Performance and Prevents of Diarrhea of Weaned Pigs by Enhancing Gut Health and Anti-Inflammatory Immune Responses. Animals. 2021; 11(8):2269. https://doi.org/10.3390/ani11082269
Chicago/Turabian StyleLee, Jeong Jae, Hyunjin Kyoung, Jin Ho Cho, Jeehwan Choe, Younghoon Kim, Yanhong Liu, Joowon Kang, Hanbae Lee, Hyeun Bum Kim, and Minho Song. 2021. "Dietary Yeast Cell Wall Improves Growth Performance and Prevents of Diarrhea of Weaned Pigs by Enhancing Gut Health and Anti-Inflammatory Immune Responses" Animals 11, no. 8: 2269. https://doi.org/10.3390/ani11082269
APA StyleLee, J. J., Kyoung, H., Cho, J. H., Choe, J., Kim, Y., Liu, Y., Kang, J., Lee, H., Kim, H. B., & Song, M. (2021). Dietary Yeast Cell Wall Improves Growth Performance and Prevents of Diarrhea of Weaned Pigs by Enhancing Gut Health and Anti-Inflammatory Immune Responses. Animals, 11(8), 2269. https://doi.org/10.3390/ani11082269