
Dead-End (dnd) Gene Cloning and Gonad-Specific Expression Pattern in Starry Flounder (Platichthys stellatus)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sampling
2.2. Total RNA Extraction and cDNA Synthesis
2.3. PCR Amplification of cDNA Fragment
2.4. 5′ and 3′ Rapid Amplification of cDNA Ends (RACE) PCR
2.5. Cloning
2.6. Phylogenetic Analysis of psdnd
2.7. qRT-PCR
2.8. Whole Mount In Situ Hybridization (WISH)
2.9. Statistical Analysis
3. Results
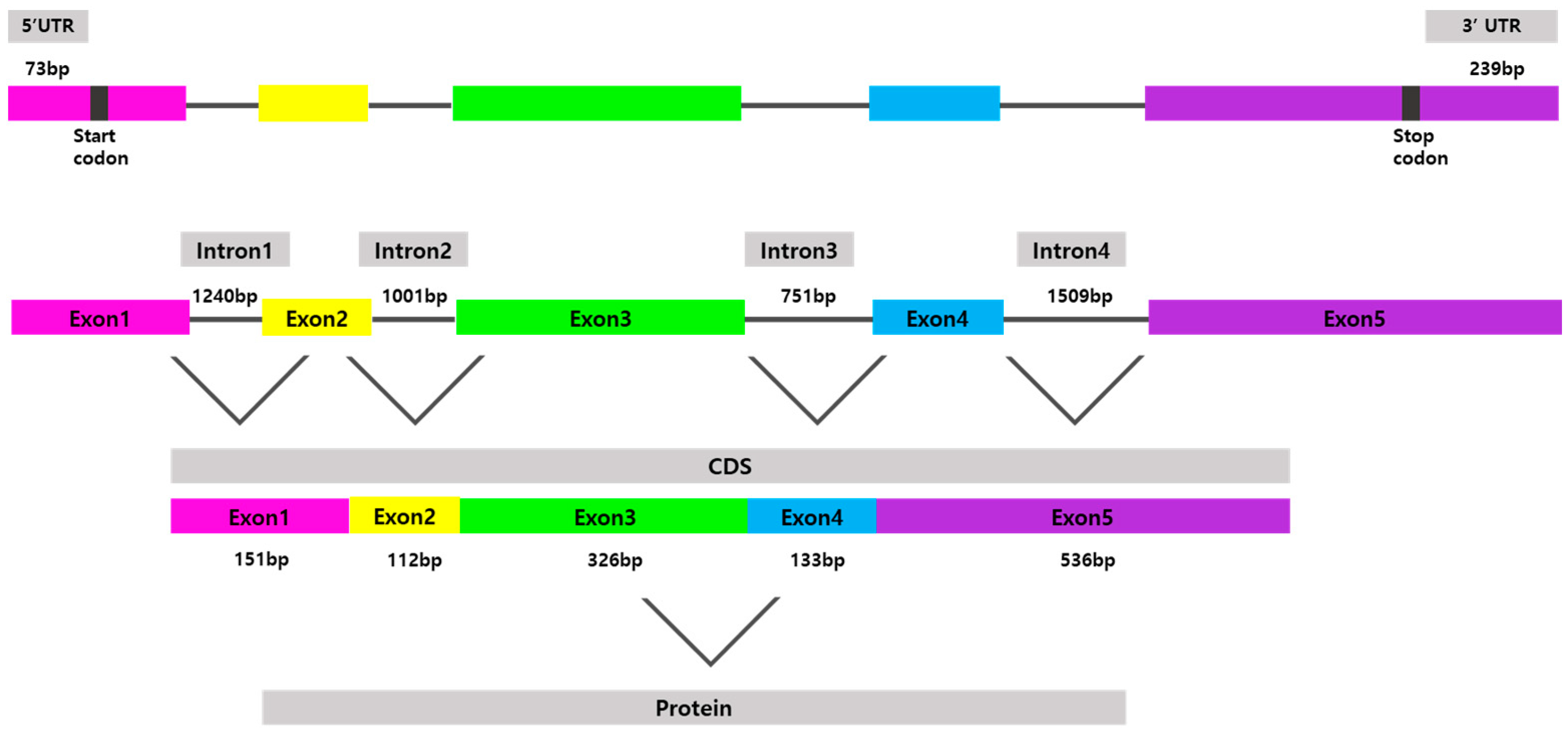
3.1. Identification of P. stellatus cDNA Sequence
3.2. Multiple Sequence Alignments
3.3. Phylogenetic Tree and Identity
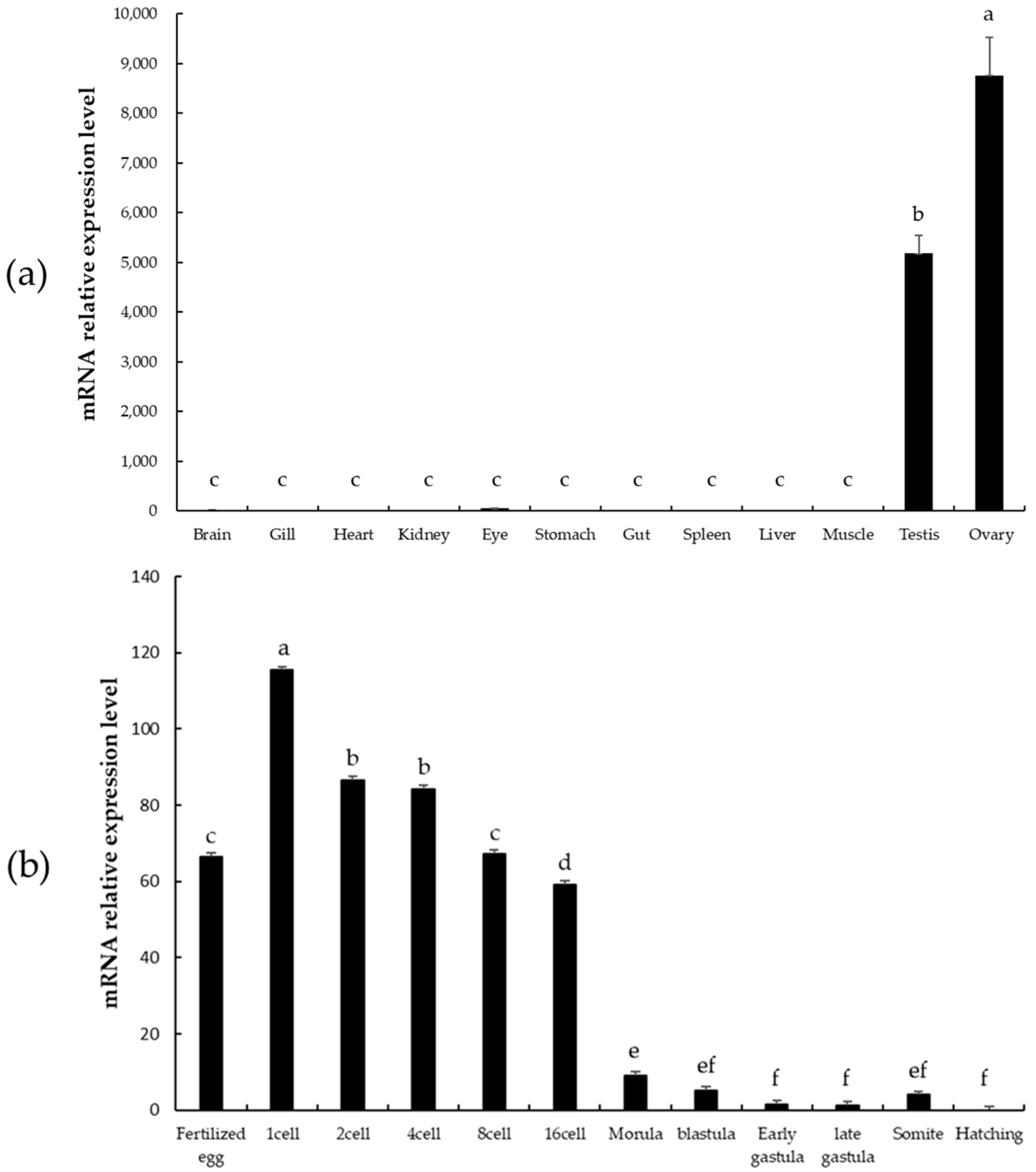
3.4. qRT-PCR
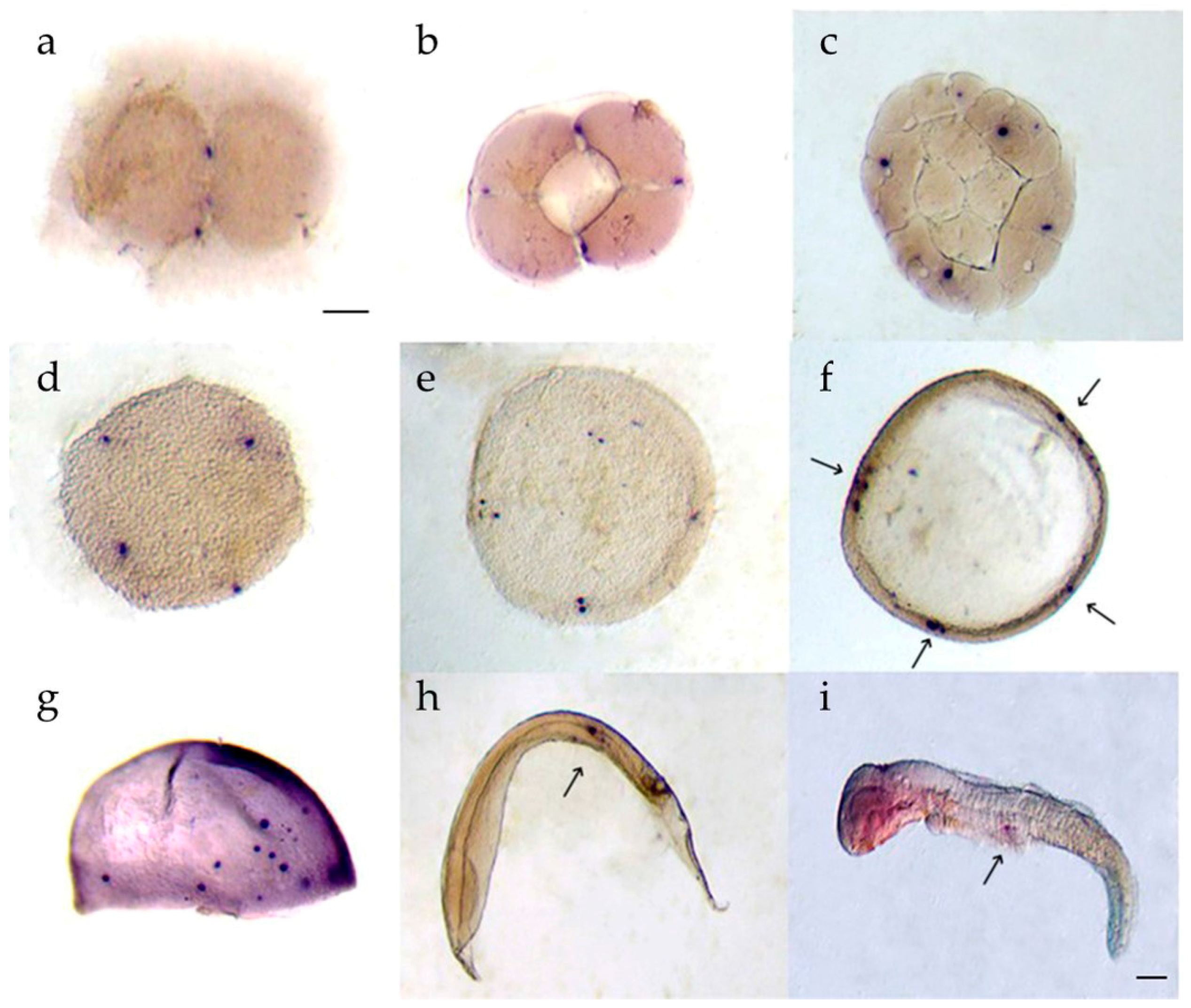
3.5. Whole-Mount In Situ Hybridization
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baloch, A.R.; Franěk, R.; Saito, T.; Pšenička, M. Dead-end (dnd) protein in fish—A review. Fish. Physiol. Biochem. 2019, 47, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Marlow, F. Primordial germ cell specification and migration. F1000Research 2015, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beer, R.L.; Draper, B.W. nanos3 maintains germline stem cells and expression of the conserved germline stem cell gene nanos2 in the zebrafish ovary. Dev. Biol. 2013, 374, 308–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komiya, T.; Itoh, K.; Ikenishi, K. Isolation and characterization of a novel gene of the DEAD box protein family which is specifically expressed in germ cells of Xenopus laevis. Dev. Biol. 1994, 162, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Houwing, S.; Kaaij, L.J. Tdrd1 acts as a molecular scaffold for Piwi proteins and piRNA targets in zebrafish. EMBO J. 2011, 30, 3298–3308. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.A.; Fulton, N.; Cowan, G. Conserved and divergent patterns of expression of DAZL, VASA and OCT4 in the germ cells of the human fetal ovary and testis. BMC Dev. Biol. 2007, 7, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Liu, Q.; Xiao, Y.; Yang, Y.; Wang, Y.; Song, Z.; You, F.; An, H.; Xiao, Z.; Xu, S. The dnd RNA identifies germ cell origin and migration in olive flounder (Paralichthys olivaceus). BioMed Res. Int. 2015, 2015, 428591. [Google Scholar]
- Kedde, M.; Strasser, M.J.; Boldajipour, B.; Oude Vrielink, J.A.F.; Slanchev, K.; le Sage, C.; Nagel, R.; Voorhoeve, P.M.; van Duijse, J.; Ørom, U.A.; et al. RNA-Binding Protein Dnd1 Inhibits MicroRNA Access to Target mRNA. Cell 2007, 131, 1273–1286. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, C.; Aggarwal, S.; Zhu, R. The mouse dead-end gene isoform α is necessary for germ cell and embryonic viability. Biochem. Biophys. Res. Commun. 2007, 355, 194–199. [Google Scholar] [CrossRef] [Green Version]
- Shinomiya, A.; Tanaka, M.; Kobayashi, T.; Nagahama, Y.; Hamaguchi, S. The vasa-like gene, olvas, identifies the migration path of primordial germ cells during embryonic body formation stage in the medaka, Oryzias latipes. Dev. Growth Differ. 2000, 42, 317–326. [Google Scholar] [CrossRef]
- Aramaki, S.; Sato, F.; Kato, T.; Soh, T.; Kato, Y.; Hattori, M.A. Molecular cloning and expression of dead end homologue in chicken primordial germ cells. Cell Tissue Res. 2007, 330, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Weidinger, G.; Stebler, J.; Slanchev, K.; Dumstrei, K.; Wise, C.; Lovell-Badge, R.; Thisse, C.; Thisse, B.; Raz, E. Dead end, a Novel Vertebrate Germ Plasm Component, Is Required for Zebrafish Primordial Germ Cell Migration and Survival. Curr. Biol. 2003, 13, 1429–1434. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Hong, N.; Xu, H.; Li, M.; Yan, Y.; Purwanti, Y.; Yi, M.; Li, Z.; Wang, L.; Hong, Y. Medaka dead end encodes a cytoplasmic protein and identifies embryonic and adult germ cells. Gene Expr. Patterns 2009, 9, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, T.; Nishimura, T.; Goto-Kazeto, R.; Kawakami, Y.; Yamaha, E.; Arai, K. Sexual dimorphism of gonadal structure and gene expression in germ cell-deficient loach, a teleost fish. Proc. Natl. Acad. Sci. USA 2010, 107, 17211–17216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, R.; Saito, T.; Takeda, T.; Fujimoto, T.; Takagi, M.; Arai, K.; Yamaha, E. Germ cells are not the primary factor for sexual fate determination in goldfish. Dev. Biol. 2012, 370, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Horvay, K.; Claußen, M.; Katzer, M.; Landgrebe, J.; Pieler, T. Xenopus Dead end mRNA is a localized maternal determinant that serves a conserved function in germ cell development. Dev. Biol. 2006, 291, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wong, T.T.; Zohar, Y. Production of reproductively sterile fish by a non-transgenic gene silencing technology. Sci. Rep. 2015, 5, 15822. [Google Scholar] [CrossRef] [Green Version]
- Hallerman, E.M.; Kapuscinski, A.R. Incorporating risk assessment and risk management into public policies on genetically modified finfish and shellfish. Aquaculture 1995, 137, 9–17. [Google Scholar] [CrossRef]
- Bartley, D.M.; Rana, K.; Immink, A.J. The use of inter-specific hybrids in aquaculture and fisheries. Rev. Fish. Biol. Fish. 2000, 10, 325–337. [Google Scholar] [CrossRef]
- Rahman, M.A.; Uehara, T.; Lawrence, J.M. Growth and heterosis of hybrids of two closely related species of Pacific sea urchins (Genus Echinometra) in Okinawa. Aquaculture 2005, 245, 121–133. [Google Scholar] [CrossRef]
- Kalsoom, U.; Salim, M.; Shahzadi, T.; Barlas, A. Growth performance and feed conversion ratio (FCR) in hybrid fish (Catla catle x Labeo rohita) fed on feed bran, rice broken and blood meal. Pak. Vet. J. 2009, 29, 55–58. [Google Scholar]
- Exadactylos, A.; Arvanitoyannis, I.S. Aquaculture Biotechnology for enhanced fish production for food consumption. In Microbial Biotechnology in Agriculture and Aquaculture, 2nd ed.; Ray, R.C., Ed.; Science Publishers Inc.: Enfield, NH, USA, 2006; pp. 453–510. [Google Scholar]
- Li, H.; Su, B.; Qin, G.; Ye, Z.; Elaswad, A.; Alsaqufi, A.; Perera, D.A.; Qin, Z.; Odin, R.; Vo, K. Repressible transgenic sterilization in channel catfish, Ictalurus punctatus, by knockdown of primordial germ cell genes with copper-sensitive constructs. Mar. Biotechnol. 2018, 20, 324–342. [Google Scholar] [CrossRef]
- Wargelius, A.; Leininger, S.; Skaftnesmo, K.O.; Kleppe, L.; Andersson, E.; Taranger, G.L.; Schulz, R.W.; Edvardsen, R.B. Dnd knockout ablates germ cells and demonstrates germ cell independent sex differentiation in Atlantic salmon. Sci. Rep. 2016, 6, 21284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linhartová, Z.; Saito, T.; Kašpar, V.; Rodina, M.; Prášková, E.; Hagihara, S.; Pšenička, M. Sterilization of sterlet Acipenser ruthenus by using knockdown agent, antisense morpholino oligonucleotide, against dead end gene. Theriogenology 2015, 84, 1246–1255. [Google Scholar] [CrossRef] [Green Version]
- Aguero, T.; Jin, Z.; Owens, D.; Malhotra, A.; Newman, K.; Yang, J.; King, M.L. Combined functions of two RRMs in Dead-end1 mimic helicase activity to promote nanos1 translation in the germline. Mol. Reprod. Dev. 2018, 85, 896–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duszczyk, M.M.; Wischnewski, H.; Kazeeva, T.; Loughlin, F.E.; von Schroetter, C.; Pradère, M.U.; Ciaudo, C.; Allain, F.H.T. The solution structure of Dead End bound to AU-rich RNA reveals an unprecedented mode of tandem RRM-RNA recognition required for mRNA repression. BioRxiv 2019, 572156. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Collodi, P. Zebrafish dead end possesses ATPase activity that is required for primordial germ cell development. FASEB J. 2010, 24, 2641–2650. [Google Scholar] [CrossRef] [Green Version]
- Lin, F.; Zhao, C.Y.; Xu, S.H.; Ma, D.Y.; Xiao, Z.Z.; Xiao, Y.S.; Xu, C.A.; Liu, Q.H.; Li, J. Germline-specific and sexually dimorphic expression of a dead end gene homologue in turbot (Scophthalmus maximus). Theriogenology 2013, 80, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Feng, G.; Chang, P.; Zhang, X.; Zhou, Q.; Zhong, X.; Qi, C.; Xie, S.; Zhao, H. Germ cell-specific expression of dead end (dnd) in rare minnow (Gobiocypris rarus). Fish. Physiol. Biochem. 2015, 41, 561–571. [Google Scholar] [CrossRef]
- Omura, C.S.; Lott, S.E. The conserved regulatory basis of mRNA contributions to the early Drosophila embryo differs between the maternal and zygotic genomes. PLoS Genet. 2020, 16, e1008645. [Google Scholar] [CrossRef] [Green Version]
- Mishima, Y.; Giraldez, A.J.; Takeda, Y.; Fujiwara, T.; Sakamoto, H.; Schier, A.F.; Inoue, K. Differential Regulation of Germline mRNAs in Soma and Germ Cells by Zebrafish miR-430. Curr. Biol. 2006, 16, 2135–2142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraldez, A.J.; Mishima, Y.; Rihel, J.; Grocock, R.J.; Van Dongen, S.; Inoue, K.; Enright, A.J.; Schier, A.F. Zebrafish MiR-430 promotes deadenylation and clearance of maternal mRNAs. Science 2006, 312, 75–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purpose | Primer Name | Primer Sequence (5′-3′) | Product Size_(bp) |
|---|---|---|---|
| Partial fragment | dnd-F | ATGAACACTGAGCGGGTGCAAGC | 1098 |
| dnd-R | CTAGTTGTTGTAAATTTCCTGCAGG | ||
| 5′ RACE | 5′ GSP1 | AGATTGCTTTTGTAAC | ≥238 |
| 5′ GSP2 | AGACAGGCCCCACAGAGCTGAAC | ≥145 | |
| 5′ GSP3 | AGCTGGTCCTCGTAGCTGTCCCGTG | ≥113 | |
| 3′ RACE | 3′ GSP1 | GCGTGCAGAACGTTTCACTGACAGCTGGG | ≥647 |
| 3′ GSP2 | TGCCACATCCTGAAGGCTCCACAGTCCTC | ≥408 | |
| Cloning | CL-F | GCGTGCAGAACGTTTCACTGACAGCTGGG | 1493 |
| CL-R | TGCCACATCCTGAAGGCTCCACAGTCCTC | ||
| qRT-PCR | dnd-qF | AGAACGTTTCACTGACAGC | 100 |
| dnd-qR | AGAACGTTTCACTGACAGC | ||
| qRT-PCR | GAPDH-qF | CCAGAACATCATCCCAGCTT | 185 |
| GAPDH-qR | GGCCTTCACAACCTTCTTGA | ||
| WISH | WISH-F | AGATCCCACGGGACAGCTAC | 761 |
| WISH-R | GTTGGTCAGCCACGCTATGAG |
| Species | Identity (%) |
|---|---|
| Starry flounder (Platichthys stellatus) | 100 |
| Atlantic halibut (Hippoglossus hippoglossus) | 85 |
| Olive flounder (Paralichthys olivaceus) | 77 |
| Turbot (Scophthalmus maximus) | 62 |
| Pacific blufin tuna (Thunnus orientalis) | 61 |
| Rainbow trout (Oncorhynchus mykiss) | 50 |
| Atlantic salmon (Salmo salar) | 49 |
| Pond loach (Misgurnus anguillicaudatus) | 39 |
| Channel catfish (Ictalurus punctatus) | 39 |
| Common carp (Cyprinus carpio) | 37 |
| Zebrafish (Danio rerio) | 35 |
| Human (Homo sapiens) | 33 |
| Mouse (Mus musculus) | 32 |
| Chicken (Gallus gallus) | 32 |
| Western clawed frog (Xenopus tropicalis) | 31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, J.-H.; Cho, Y.-S.; Lee, H.-B.; Park, J.-Y.; Lim, H.-K. Dead-End (dnd) Gene Cloning and Gonad-Specific Expression Pattern in Starry Flounder (Platichthys stellatus). Animals 2021, 11, 2256. https://doi.org/10.3390/ani11082256
Yoon J-H, Cho Y-S, Lee H-B, Park J-Y, Lim H-K. Dead-End (dnd) Gene Cloning and Gonad-Specific Expression Pattern in Starry Flounder (Platichthys stellatus). Animals. 2021; 11(8):2256. https://doi.org/10.3390/ani11082256
Chicago/Turabian StyleYoon, Ji-Hye, Youn-Su Cho, Hyo-Bin Lee, Jung-Yeol Park, and Han-Kyu Lim. 2021. "Dead-End (dnd) Gene Cloning and Gonad-Specific Expression Pattern in Starry Flounder (Platichthys stellatus)" Animals 11, no. 8: 2256. https://doi.org/10.3390/ani11082256
APA StyleYoon, J.-H., Cho, Y.-S., Lee, H.-B., Park, J.-Y., & Lim, H.-K. (2021). Dead-End (dnd) Gene Cloning and Gonad-Specific Expression Pattern in Starry Flounder (Platichthys stellatus). Animals, 11(8), 2256. https://doi.org/10.3390/ani11082256