
Impact of Different Durations of Fasting on Intestinal Autophagy and Serum Metabolome in Broiler Chicken

 , , , , ,
, , , , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
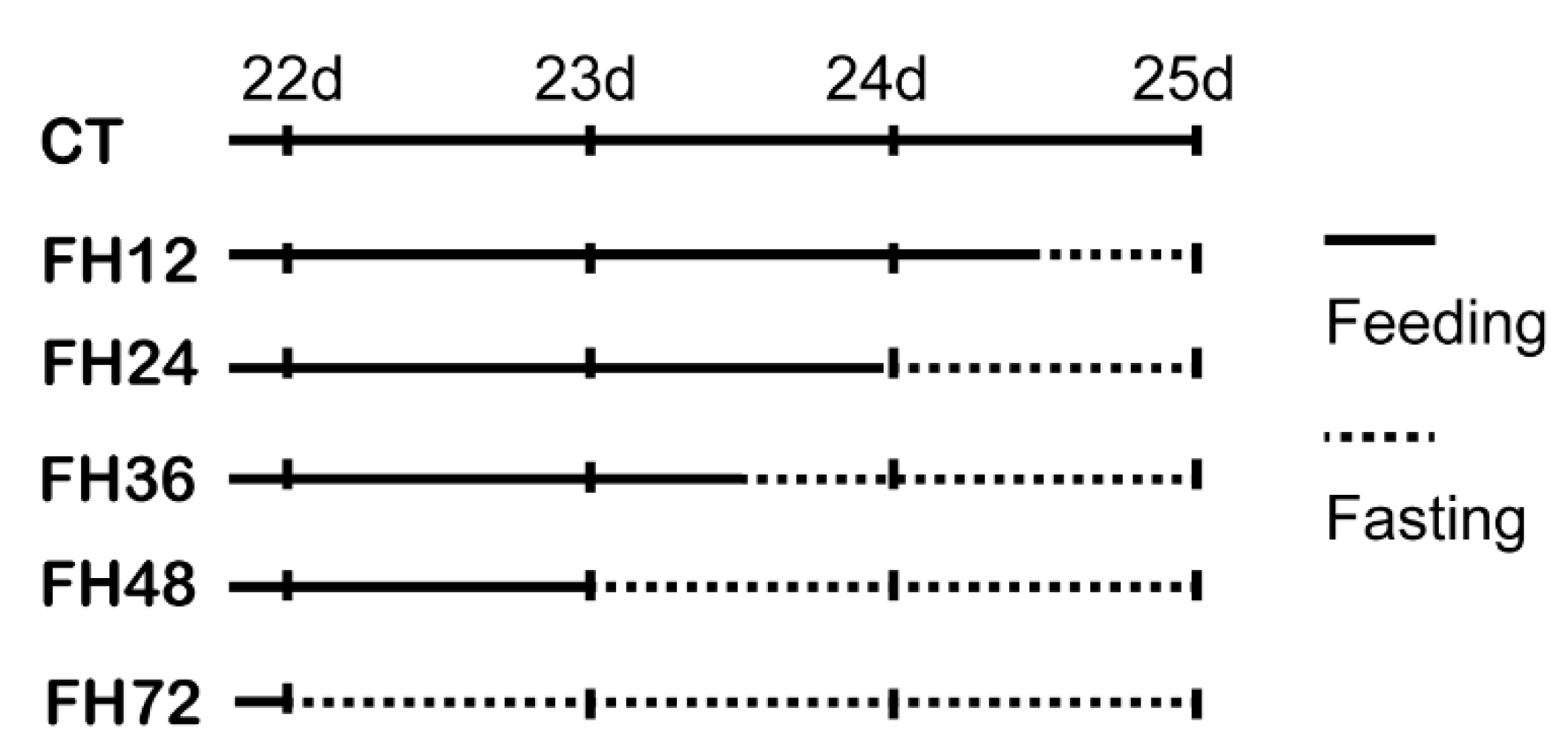
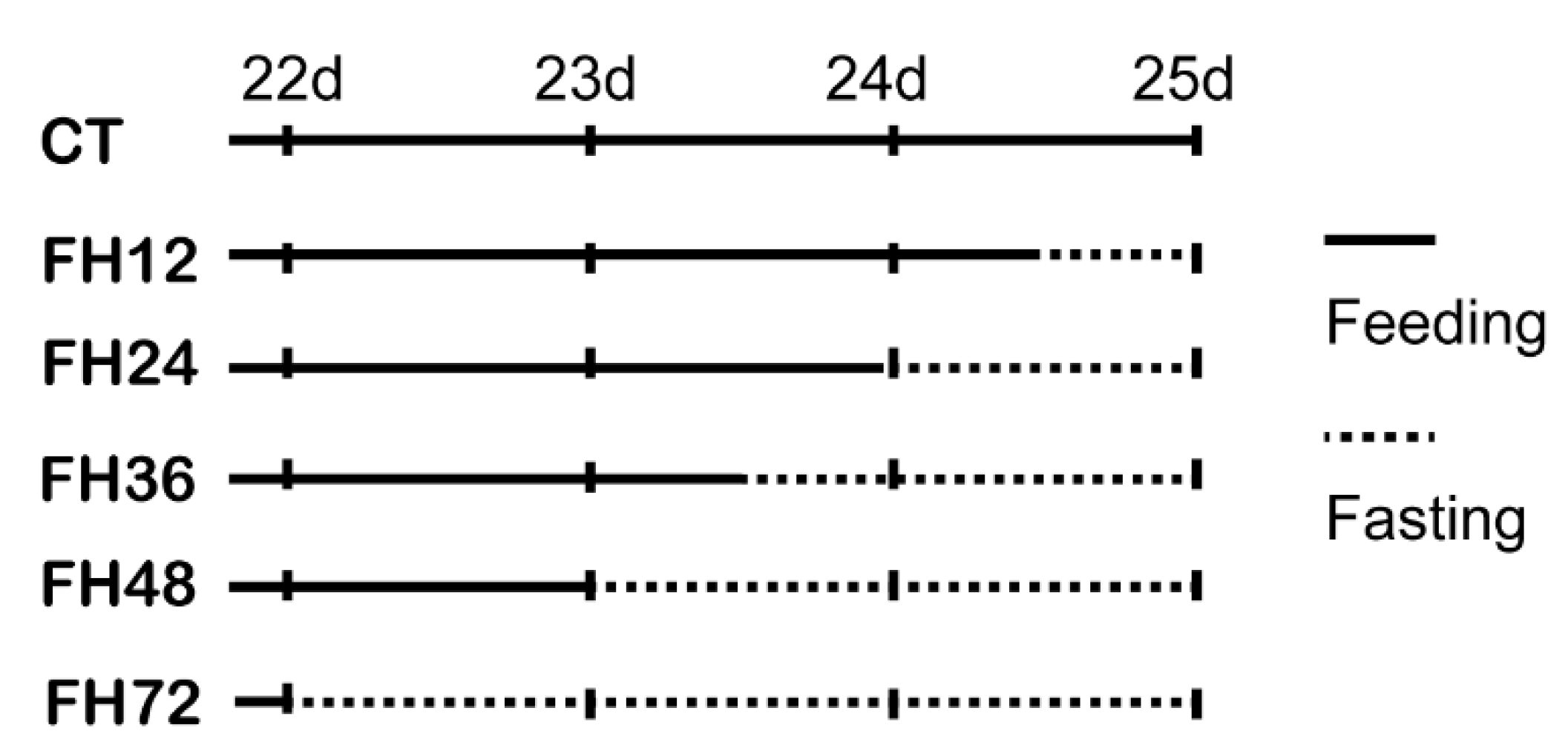
2.1. Bird Management
2.2. Sample Collection
2.3. Chemical Analysis
2.4. Autophagy-Related Gene Expression
2.5. Blood Serum Preparation
2.6. Liquid Chromatography–Mass Spectrometry Analysis
2.7. Data Preprocessing and Statistical Analysis
3. Results
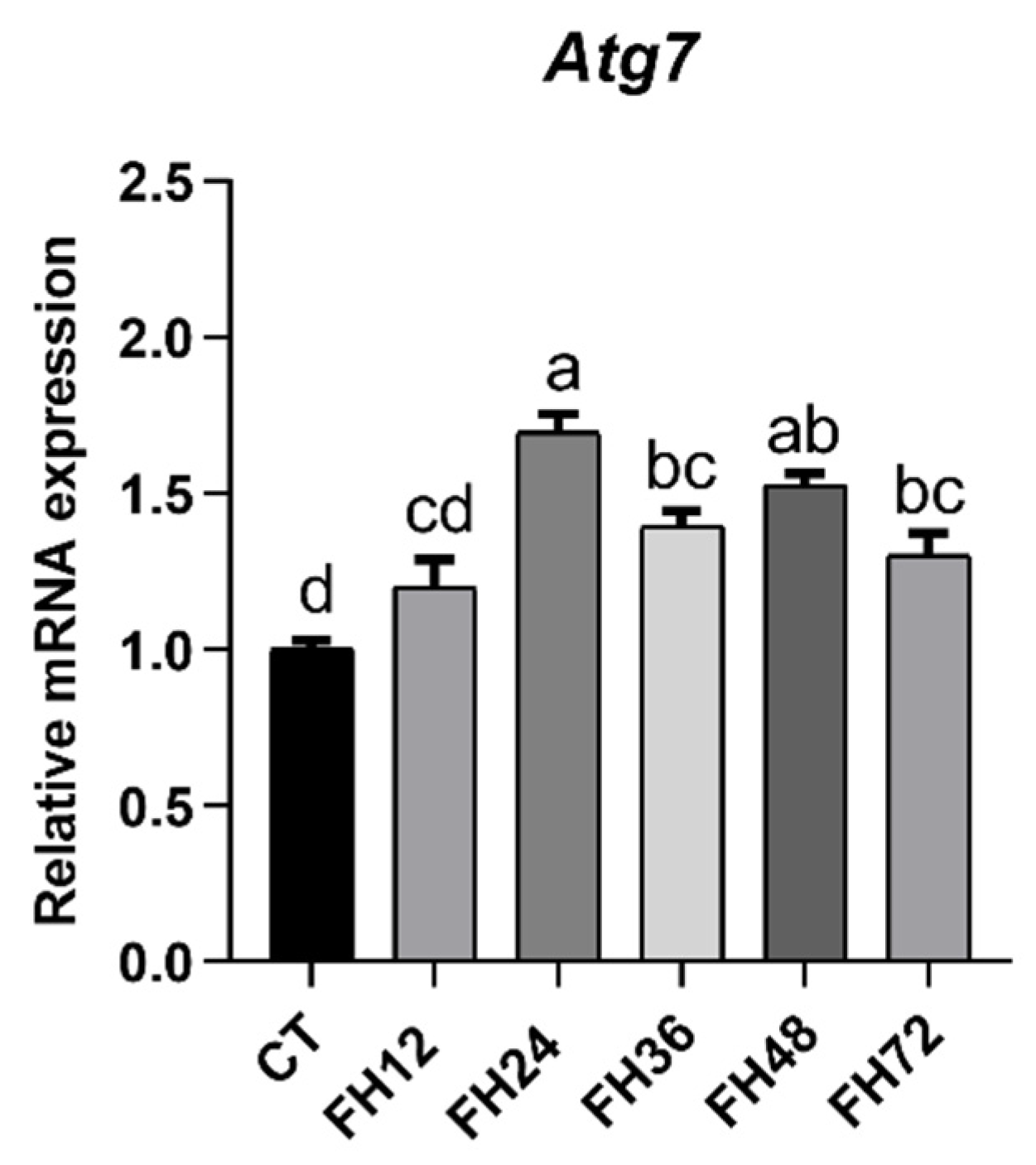
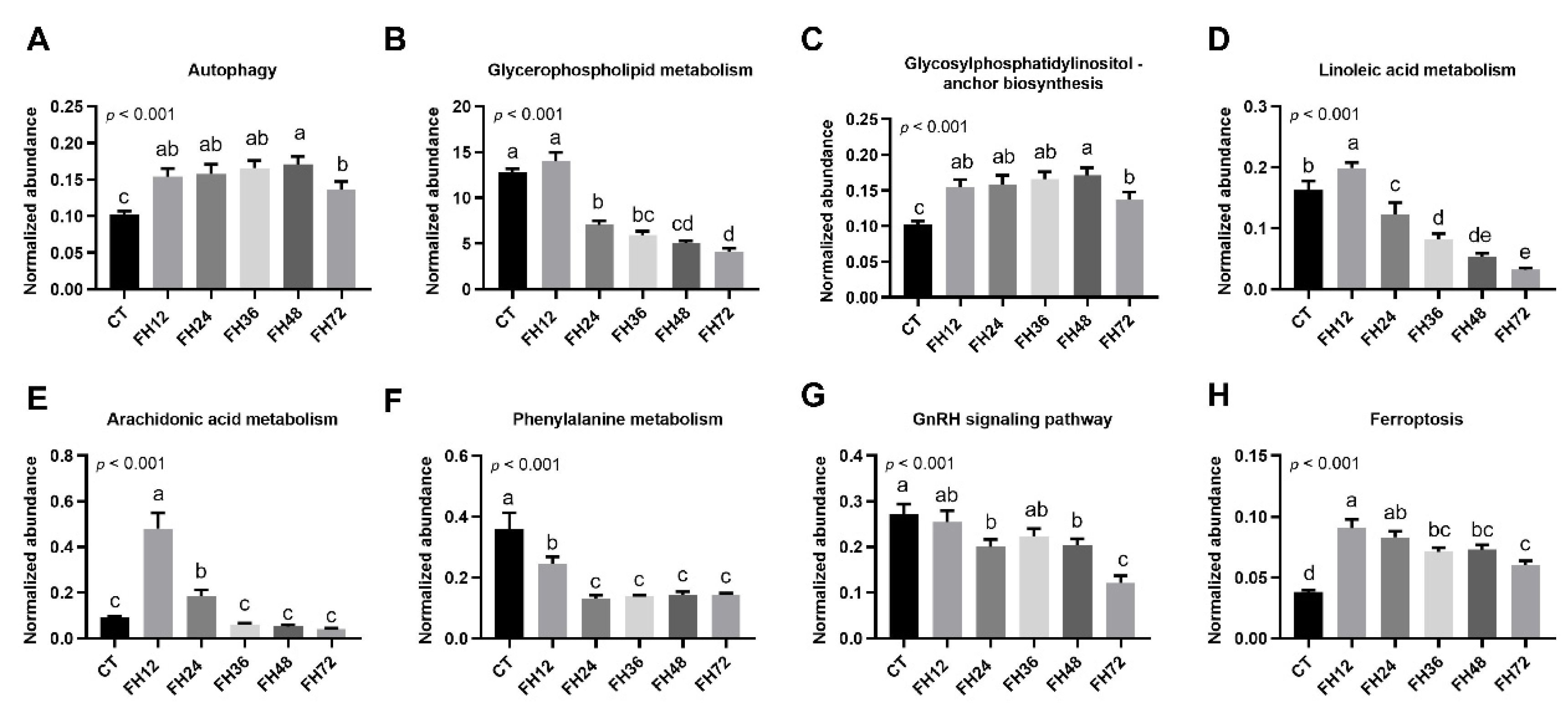
3.1. Intestinal Autophagy at Different Durations of Fasting
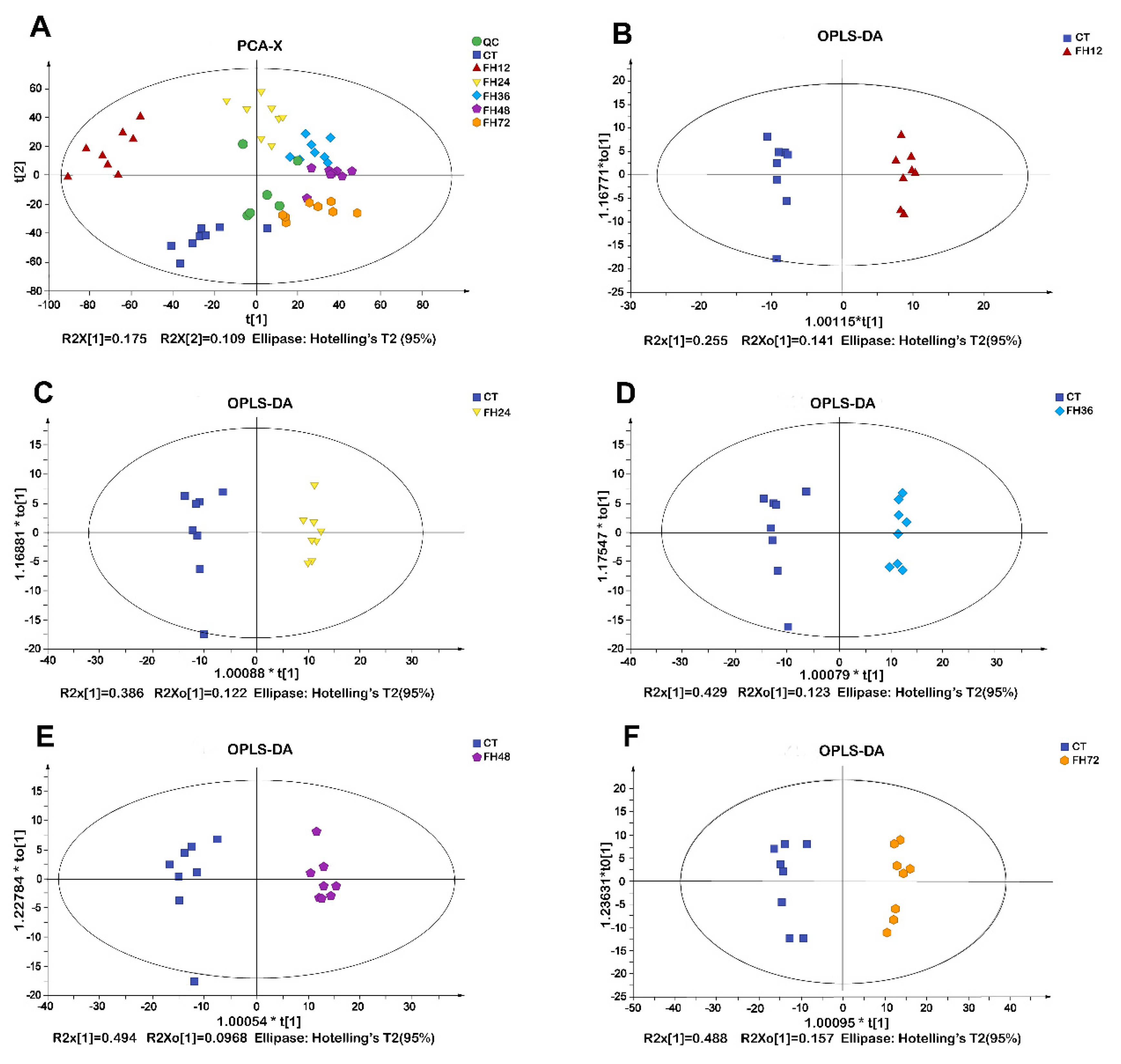
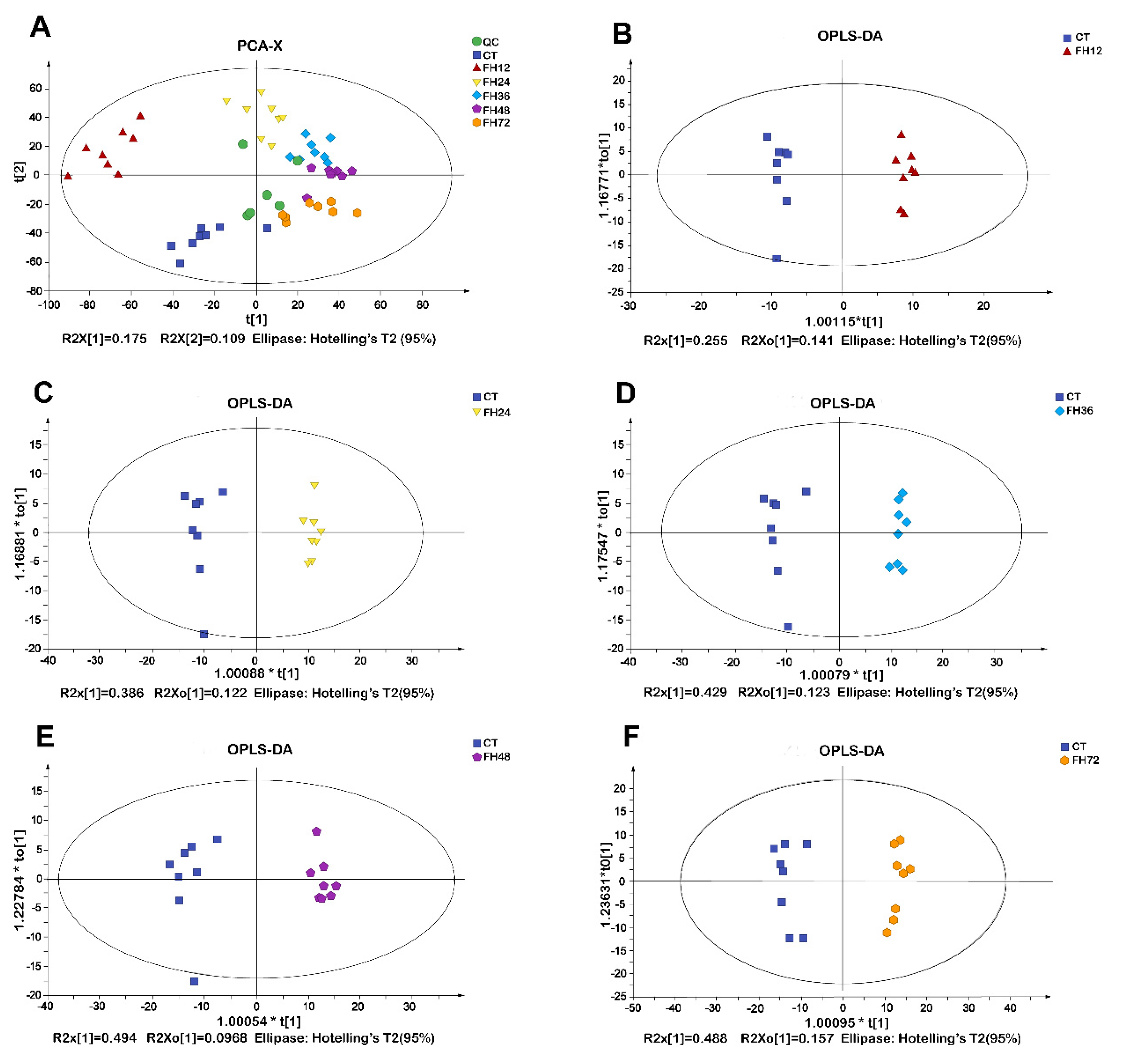
3.2. PCA and OPLS-DA Analysis of Chicken Serum at Different Durations of Fasting
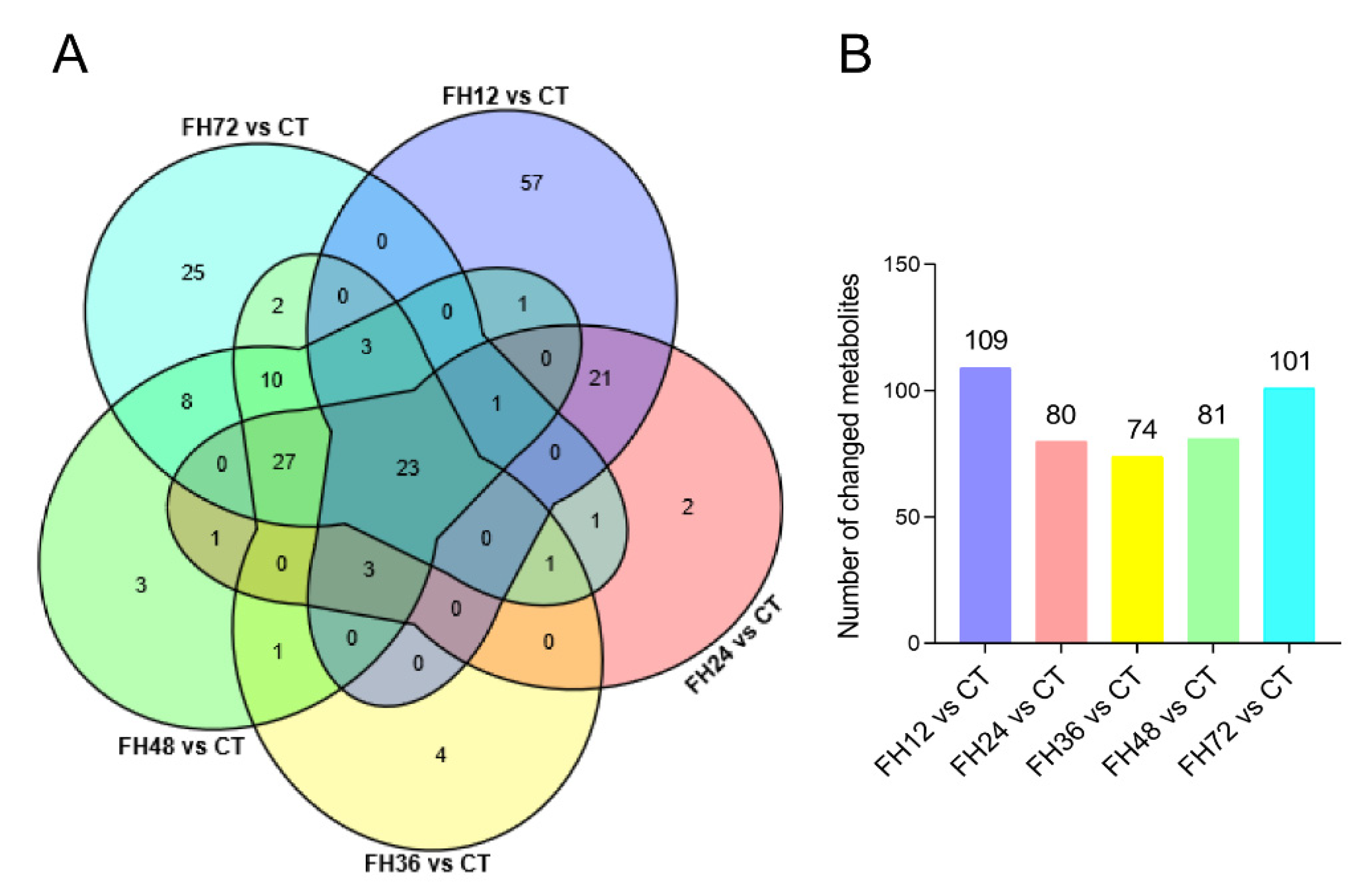
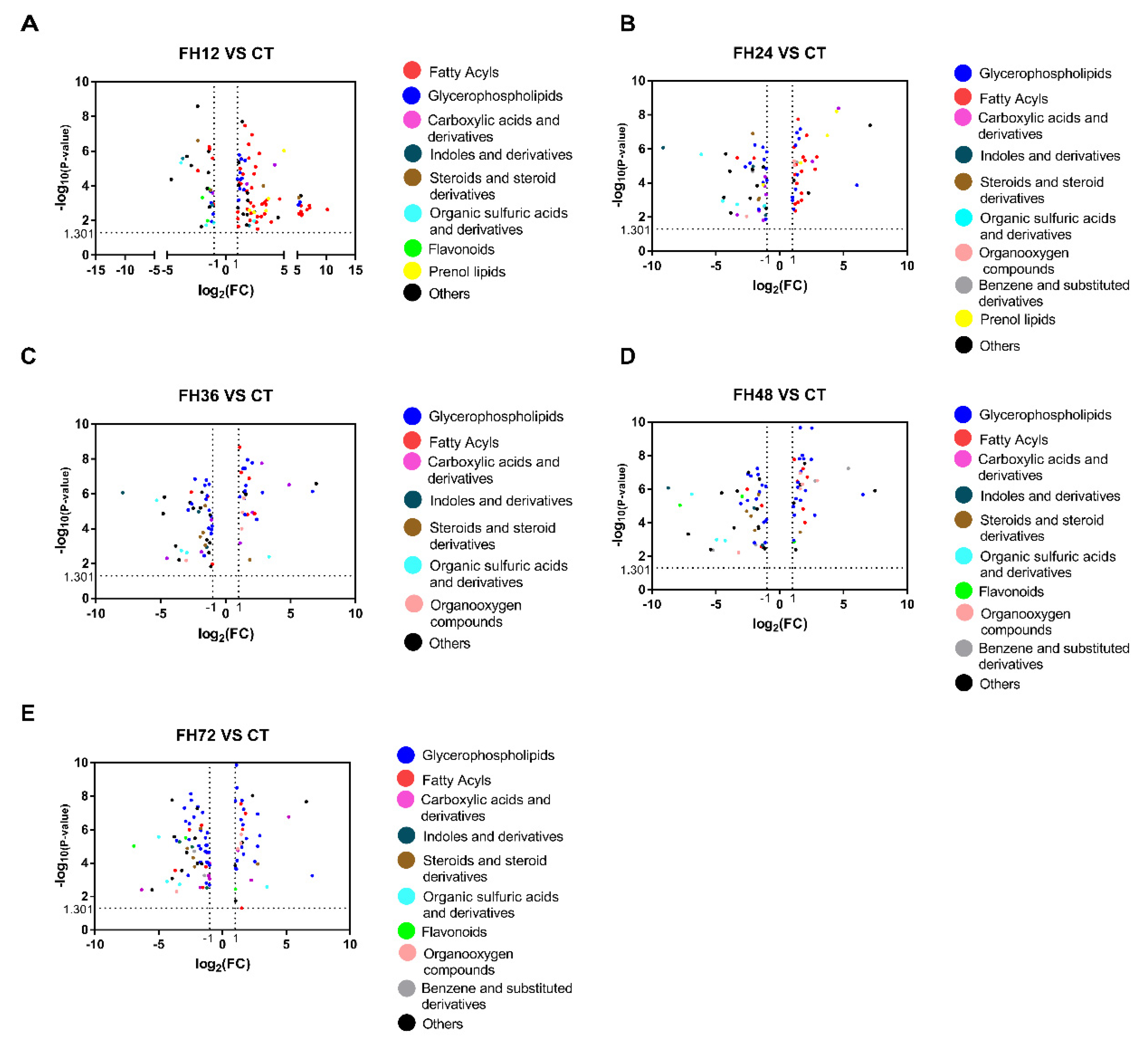
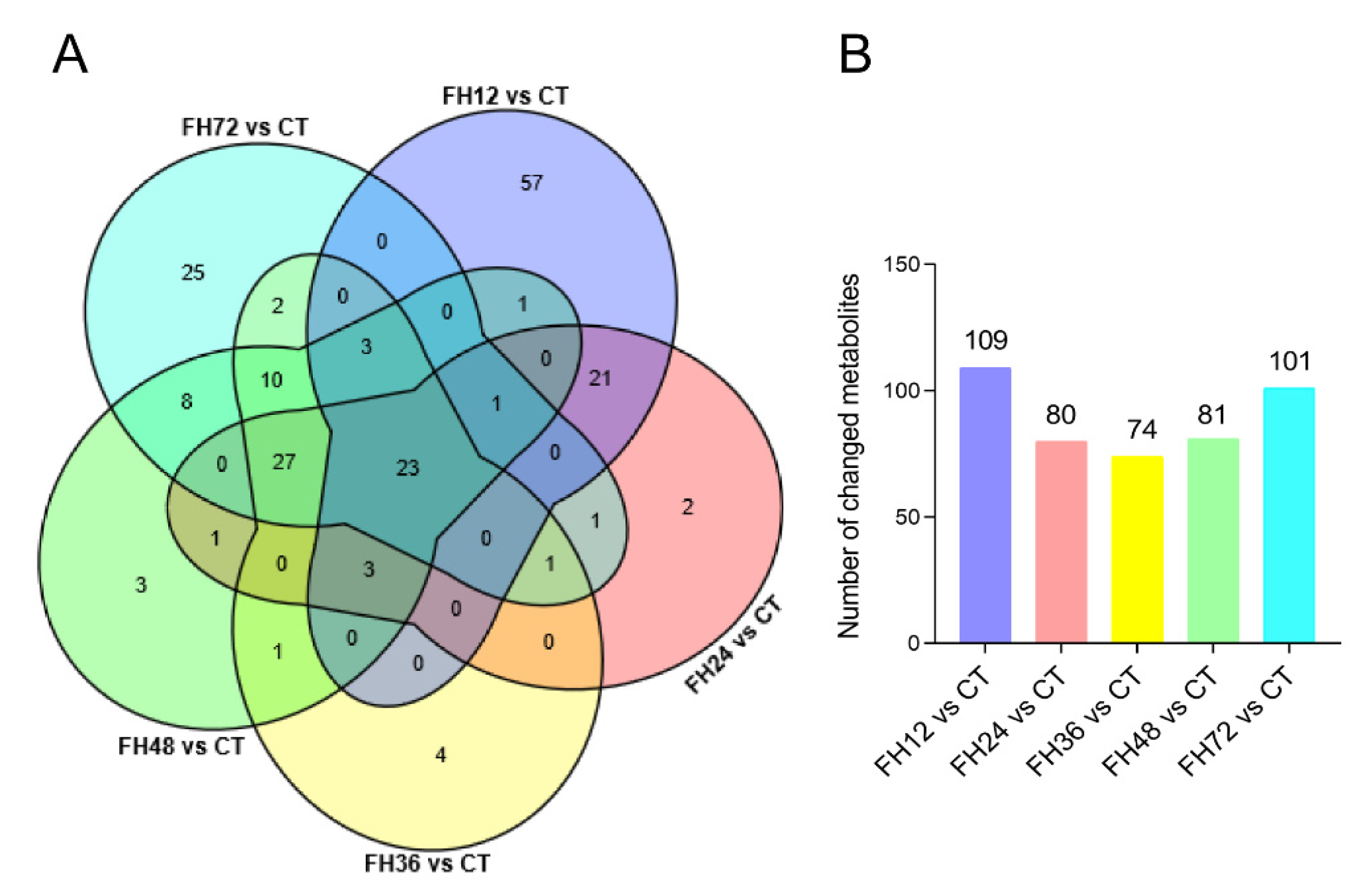
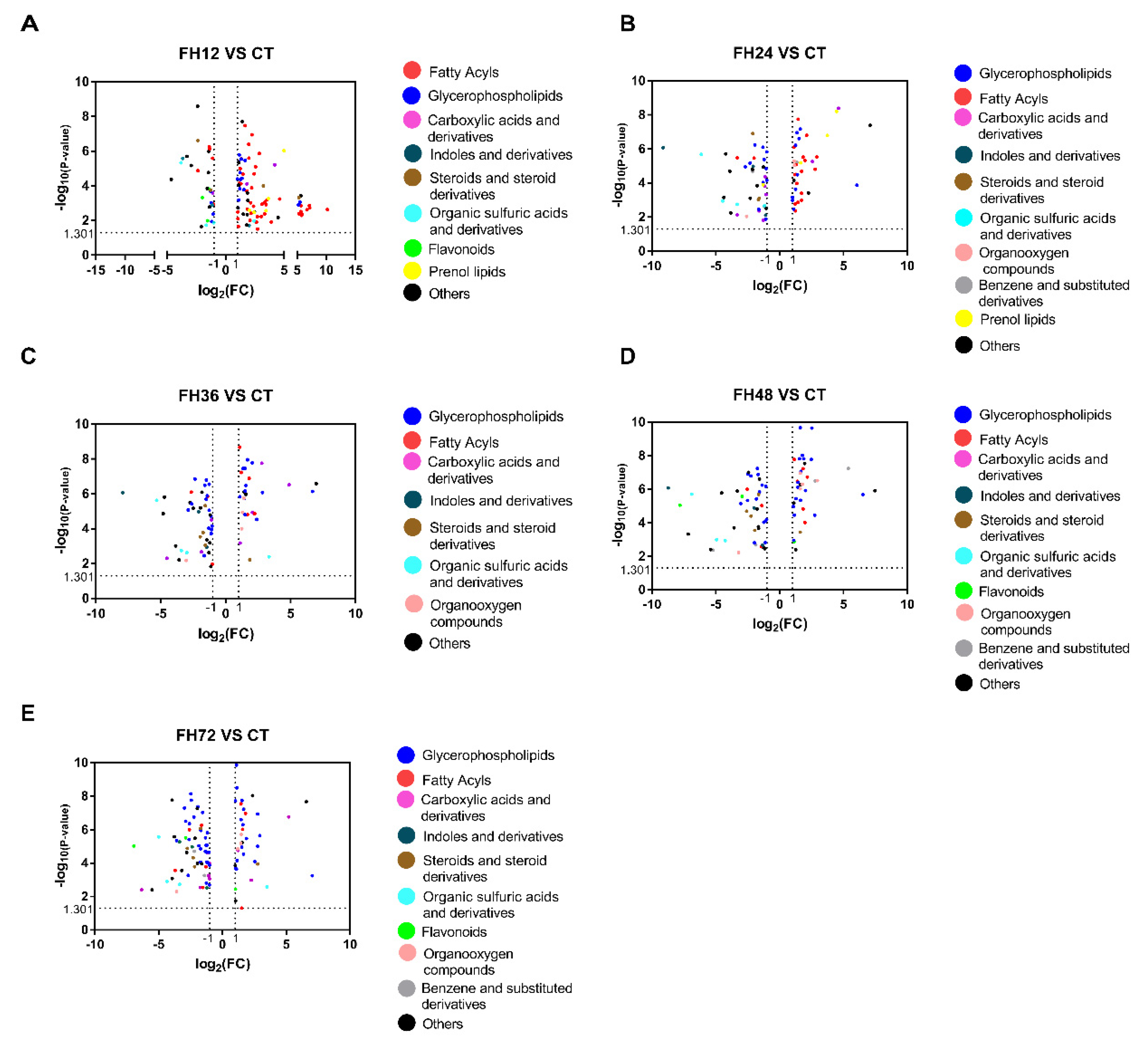
3.3. Serum Metabolic Profiles Changes with Different Durations of Fasting
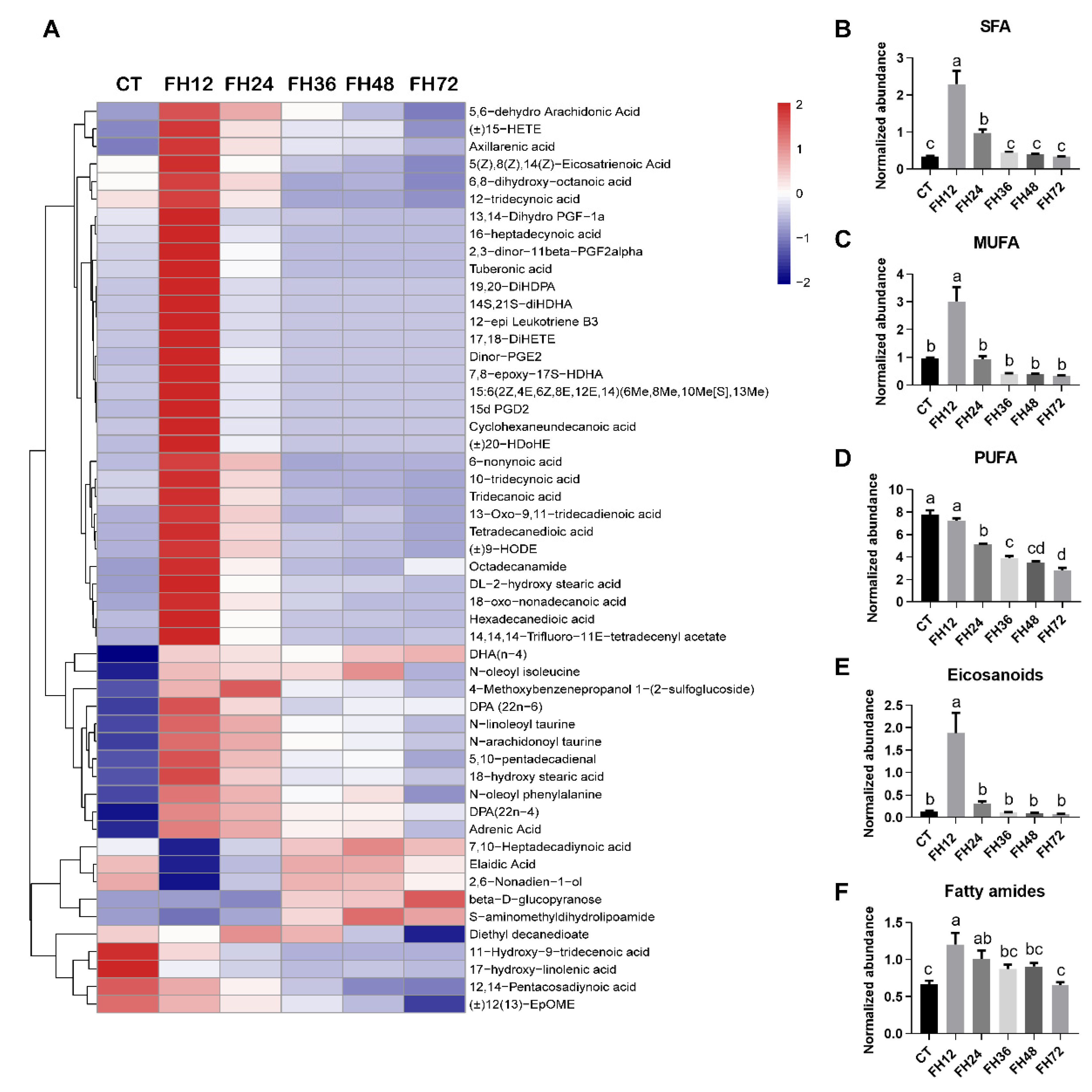
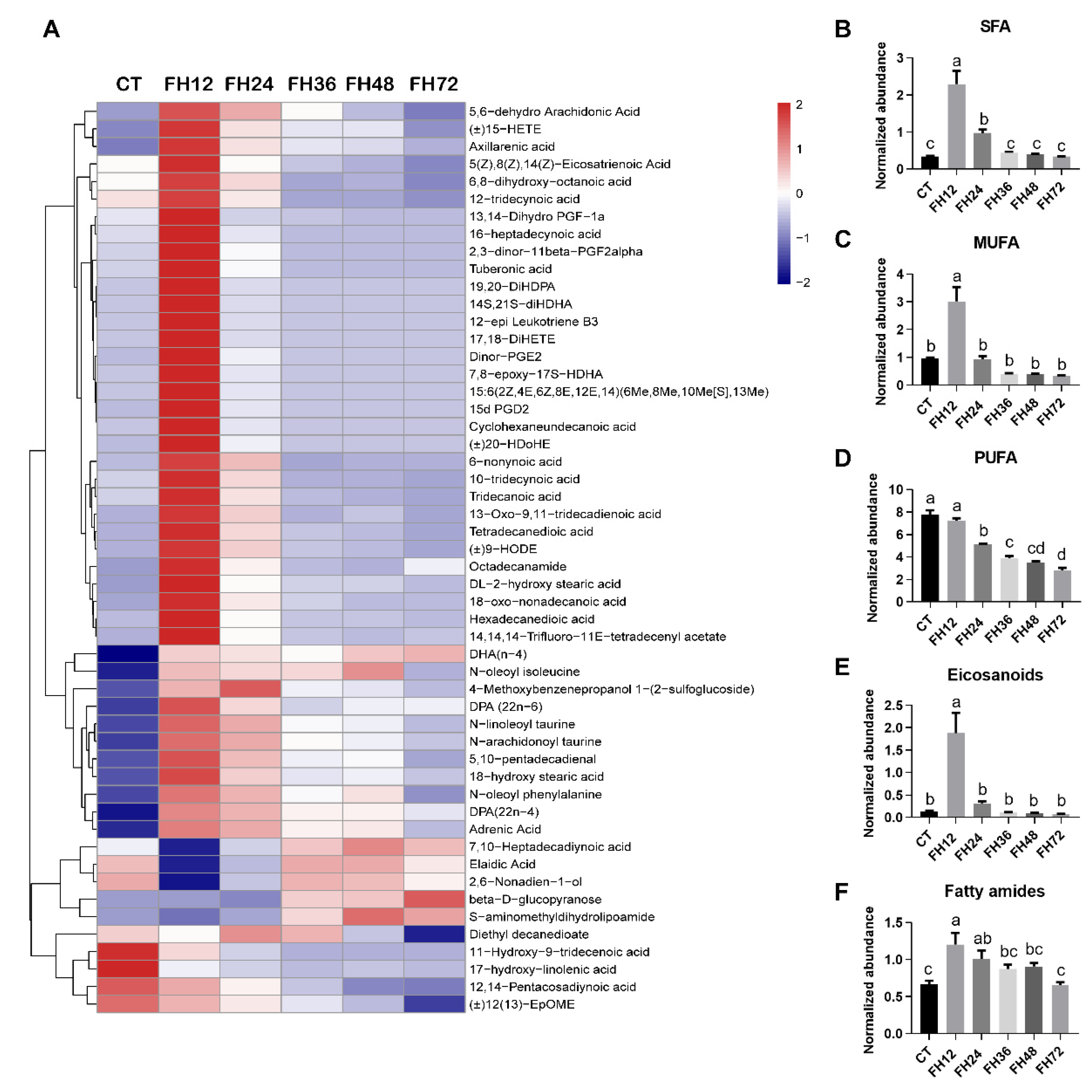
3.4. Changes in Serum Fatty Acyl Metabolism with Different Durations of Fasting
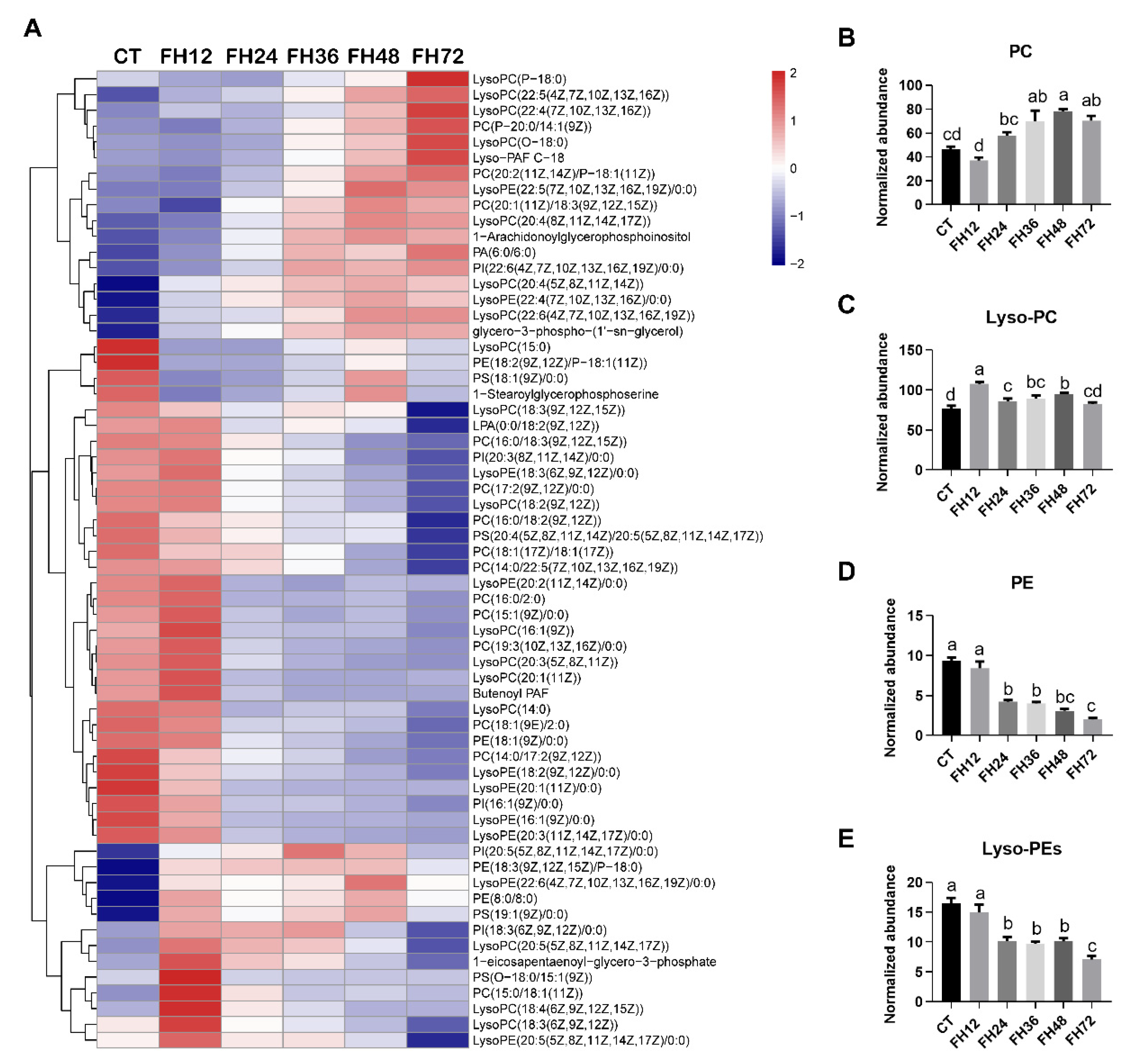
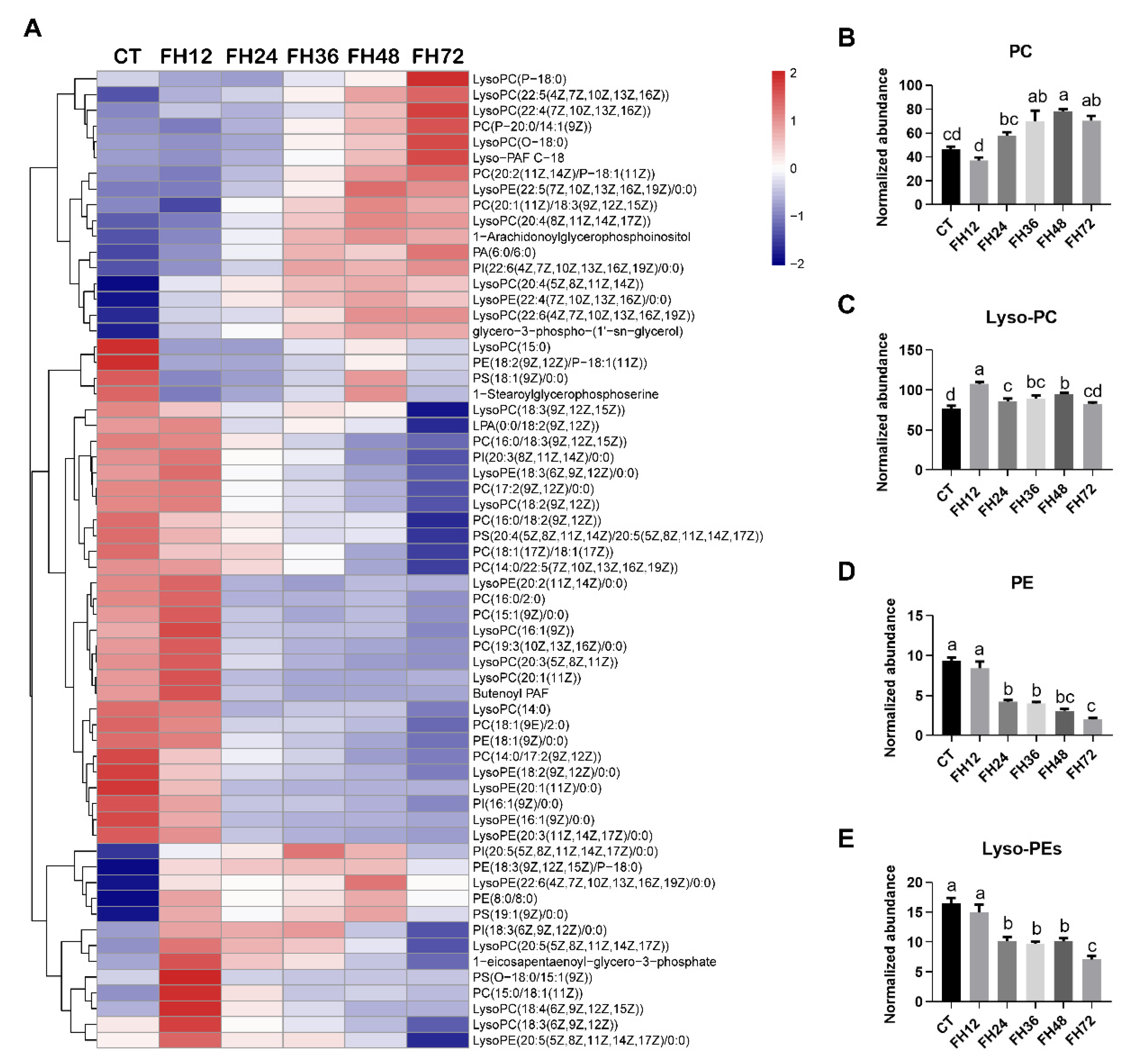
3.5. Changes in Serum Glycerophospholipids Metabolism with Different Durations of Fasting
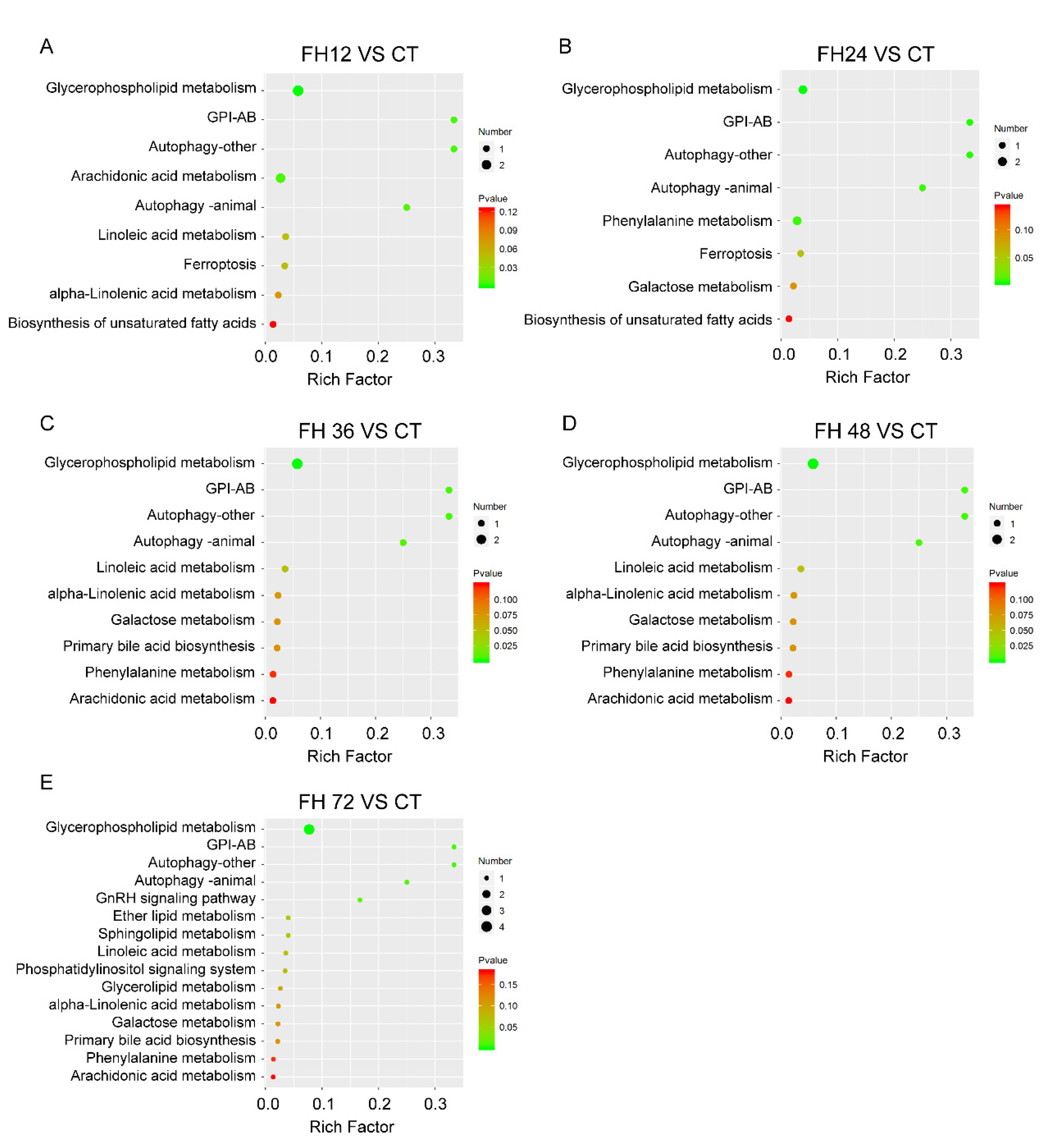
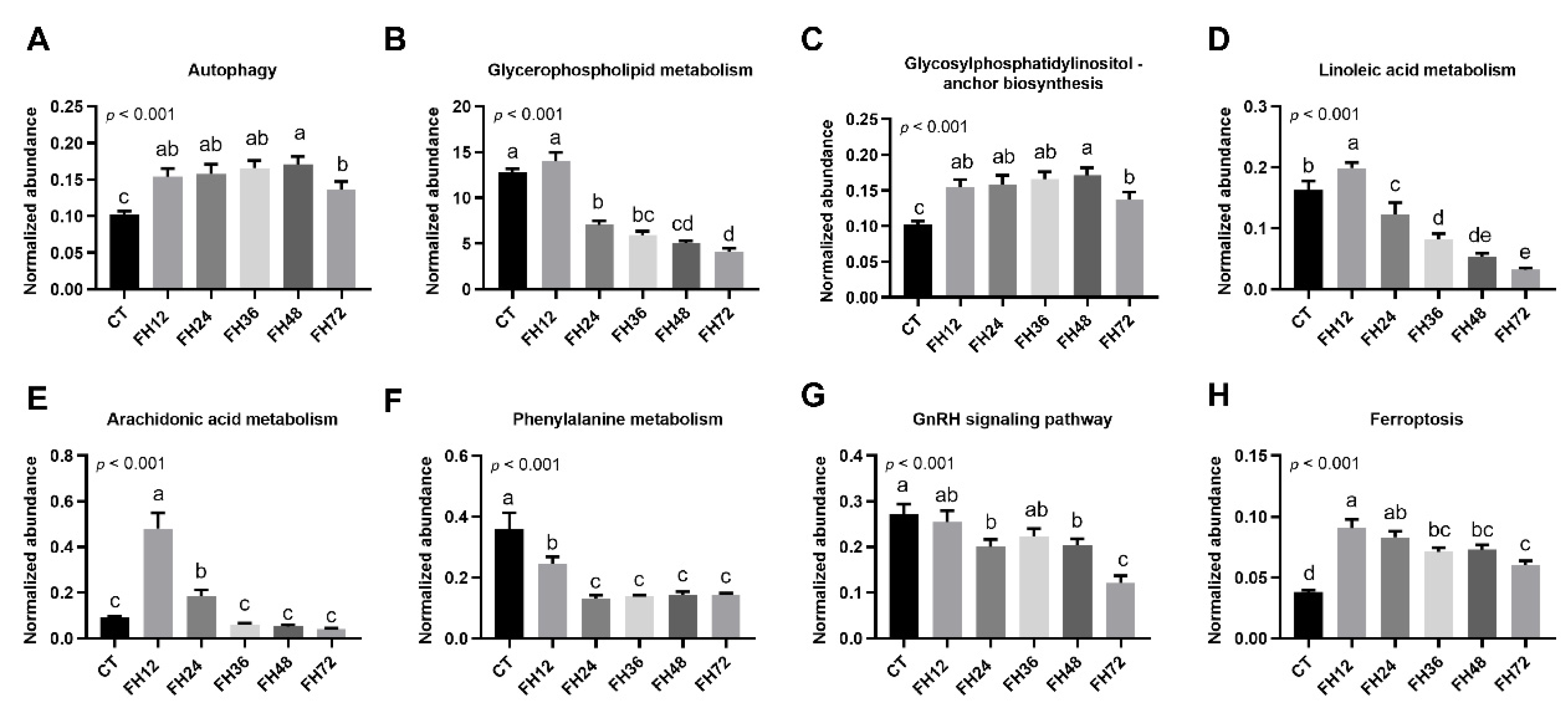
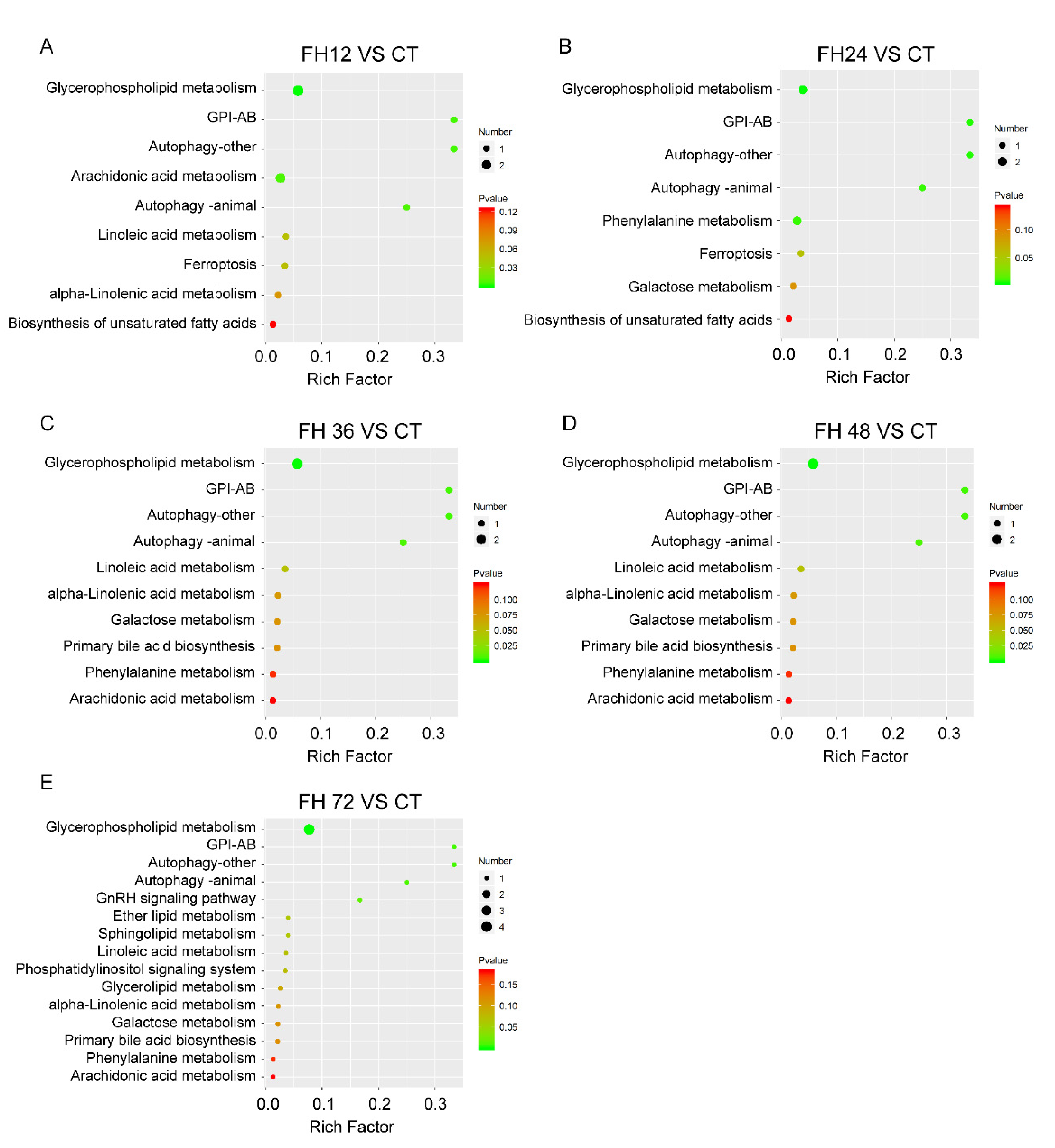
3.6. Changes in Serum Metabolic Pathway with Different Durations of Fasting
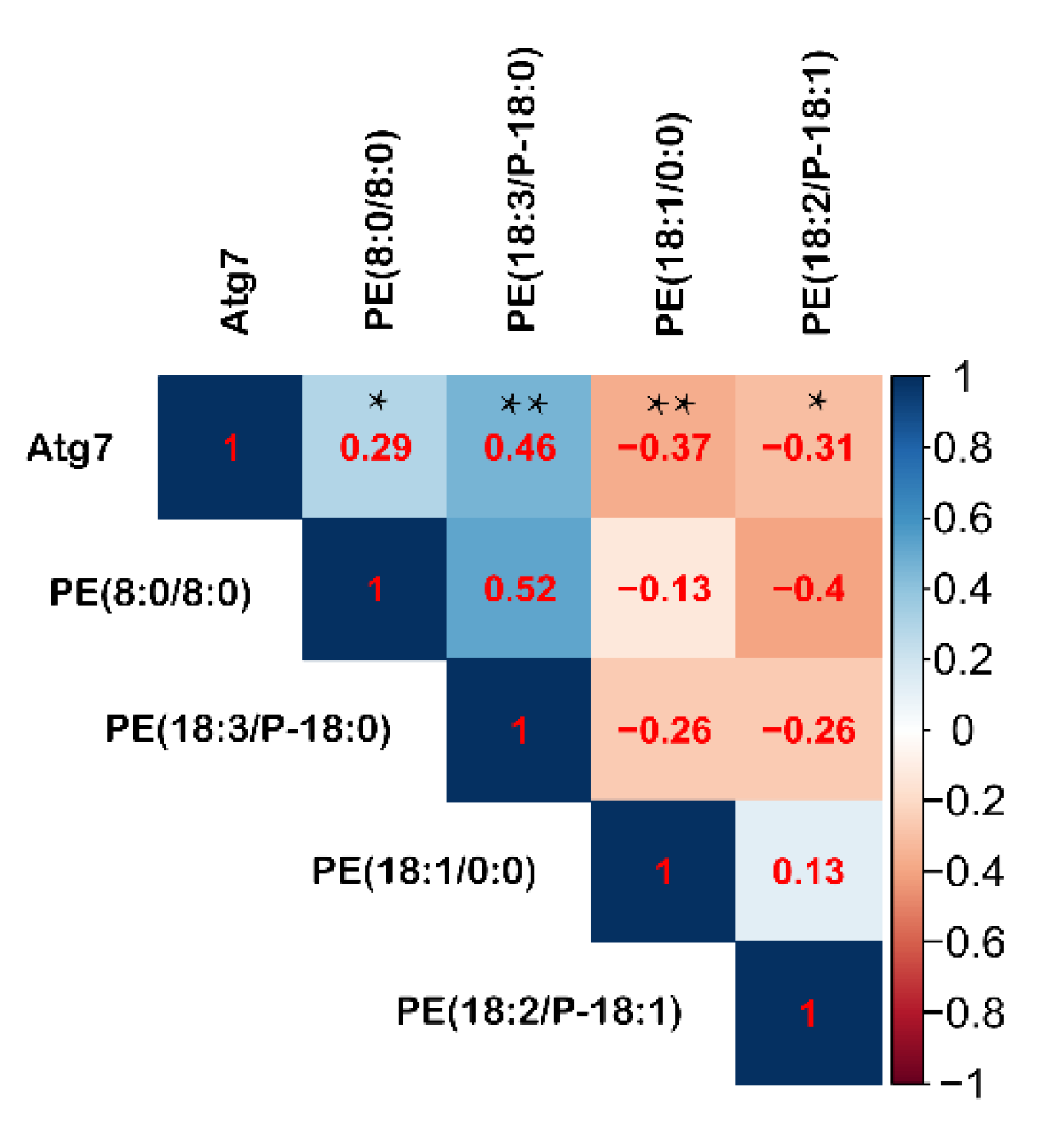
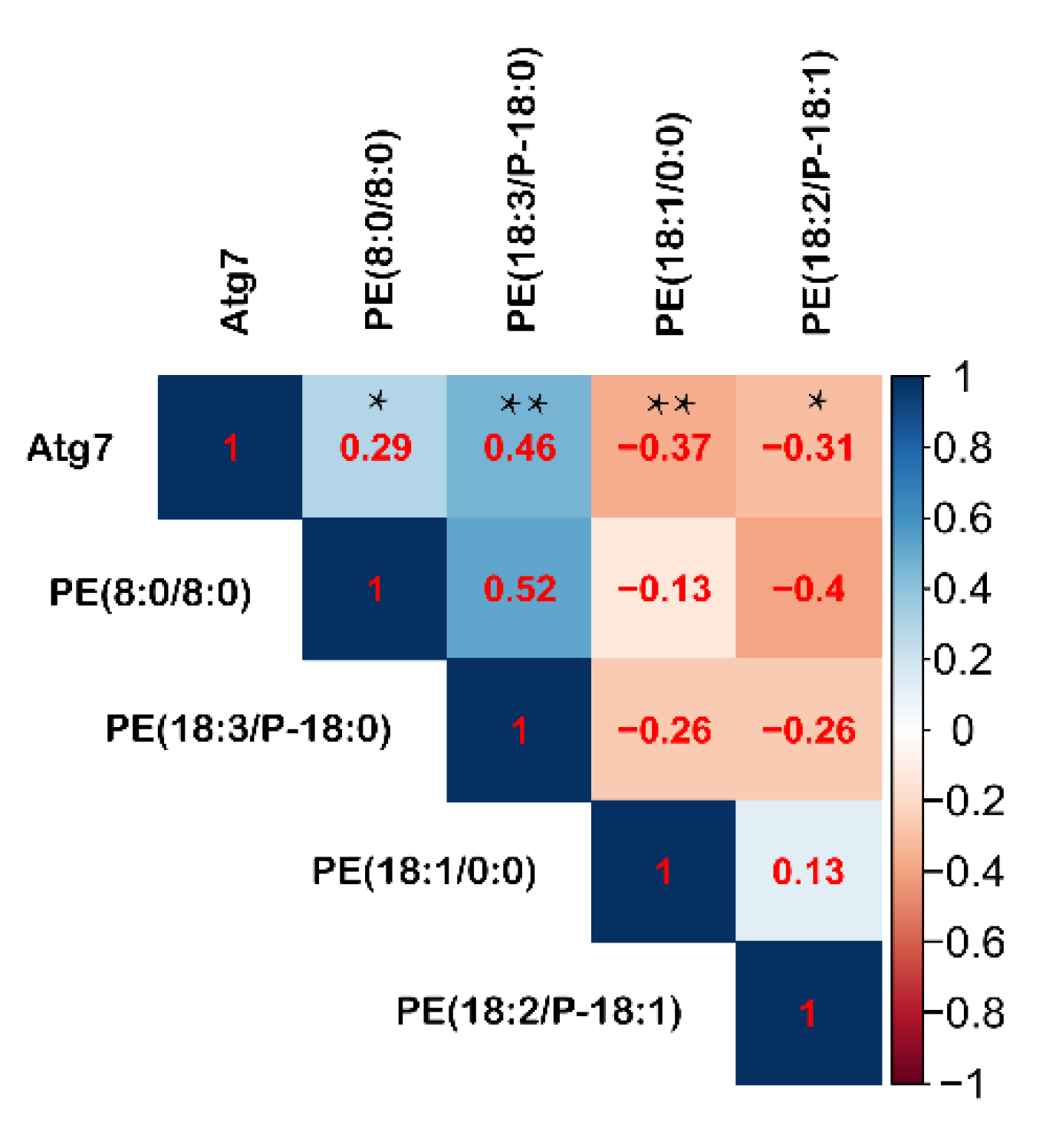
3.7. Pearson’s Correlations between Intestinal Autophagy and Serum PE during Fasting
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Malinowski, B.; Zalewska, K.; Węsierska, A.; Sokołowska, M.M.; Socha, M.; Liczner, G.; Pawlak-Osińska, K.; Wiciński, M. Intermittent fasting in cardiovascular disorders—An overview. Nutrients 2019, 11, 673. [Google Scholar] [CrossRef] [Green Version]
- Bourdillon, A.; Carre, B.; Conan, L.; Duperray, J.; Huyghebaert, G.; Leclercq, B.; Lessire, M.; McNab, J.; Wiseman, J. European reference method for the in vivo determination of metabolisable energy with adult cockerels: Reproducibility, effect of food intake and comparison with individual laboratory methods. Br. Poult. Sci. 1990, 31, 557–565. [Google Scholar] [CrossRef]
- Aardsma, M.P.; Mitchell, R.D.; Parsons, C.M. Relative metabolizable energy values for fats and oils in young broilers and adult roosters. Poult. Sci. 2017, 96, 2320–2329. [Google Scholar] [CrossRef]
- McNab, J.M.; Blair, J.C. Modified assay for true and apparent metabolisable energy based on tube feeding. Br. Poult. Sci. 1988, 29, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Farrell, D.J. Rapid determination of metabolisable energy of foods using cockerels. Br. Poult. Sci. 1978, 19, 303–308. [Google Scholar] [CrossRef]
- Inoki, K.; Kim, J.; Guan, K. AMPK and mTOR in cellular energy homeostasis and drug targets. Annu. Rev. Pharmacol. 2012, 52, 381–400. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N. Autophagy: Process and function. Genes Dev. 2007, 21, 2861–2873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, T.; Nan, Z.; Miao, C.; Jin, X.; Yang, W.; Wang, Z.; Tu, Y.; Bao, H.; Lyu, J.; Zheng, H.; et al. The autophagy-related gene Atg101 in Drosophila regulates both neuron and midgut homeostasis. J. Biol. Chem. 2019, 294, 5666–5676. [Google Scholar] [CrossRef]
- Baxt, L.A.; Xavier, R.J. Role of autophagy in the maintenance of intestinal homeostasis. Gastroenterology 2015, 149, 553–562. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Klionsky, D.J. Mammalian autophagy: Core molecular machinery and signaling regulation. Curr. Opin. Cell Biol. 2010, 22, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Pott, J.; Kabat, A.M.; Maloy, K.J. Intestinal Epithelial Cell Autophagy Is Required to Protect against TNF-Induced Apoptosis during Chronic Colitis in Mice. Cell Host Microbe 2018, 23, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Gelino, S.; Chang, J.T.; Kumsta, C.; She, X.; Davis, A.; Nguyen, C.; Panowski, S.; Hansen, M. Intestinal autophagy improves healthspan and longevity in C. elegans during dietary restriction. PLoS Genet. 2016, 12, e1006135. [Google Scholar]
- Hay-Koren, A.; Bialik, S.; Levin-Salomon, V.; Kimchi, A. Changes in cIAP2, survivin and BimEL expression characterize the switch from autophagy to apoptosis in prolonged starvation. J. Intern. Med. 2017, 281, 458–470. [Google Scholar] [CrossRef] [Green Version]
- Longo, V.D.; Mattson, M.P. Fasting: Molecular mechanisms and clinical applications. Cell Metab. 2014, 19, 181–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Yu, C.; Pan, Y.; Zhou, X.; Xin, R.; Wang, Y.; Ma, W.; Gao, R.; Wang, C.; Wu, Y. Metabolomics analysis of serum from subjects after occupational exposure to acrylamide using UPLC-MS. Mol. Cell Endocrinol. 2017, 444, 67–75. [Google Scholar] [CrossRef]
- Lu, Z.; He, X.; Ma, B.; Zhang, L.; Li, J.; Jiang, Y.; Zhou, G.; Gao, F. Serum metabolomics study of nutrient metabolic variations in chronic heat-stressed broilers. Br. J. Nutr. 2018, 119, 771–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papandreou, C.; Bulló, M.; Ruiz-Canela, M.; Dennis, C.; Deik, A.; Wang, D.; Guasch-Ferré, M.; Yu, E.; Razquin, C.; Corella, D.; et al. Plasma metabolites predict both insulin resistance and incident type 2 diabetes: A metabolomics approach within the Prevención con Dieta Mediterránea (PREDIMED) study. Am. J. Clin. Nutr. 2019, 109, 626–634. [Google Scholar] [CrossRef]
- Mignon-Grasteau, S.; Beauclercq, S.; Urvoix, S.; Le Bihan-Duval, E. Interest in the serum color as an indirect criterion of selection of digestive efficiency in chickens. Poult. Sci. 2020, 99, 702–707. [Google Scholar] [CrossRef]
- Vizcarra, F.R.; Verghese, M.; Vizcarra, J.A. Effect of short- and long-term feed restriction on ghrelin concentrations in turkeys. Poult. Sci. 2018, 97, 2183. [Google Scholar] [CrossRef] [PubMed]
- Agostini, P.S.; Dalibard, P.; Mercier, Y.; Van der Aar, P.; Van der Klis, J.D. Comparison of methionine sources around requirement levels using a methionine efficacy method in 0 to 28 day old broilers. Poult. Sci. 2016, 95, 560–569. [Google Scholar] [CrossRef]
- Kim, E.J.; Utterback, P.L.; Parsons, C.M. Development of a precision-fed ileal amino acid digestibility assay using 3-week-old broiler chicks. Poult. Sci. 2011, 90, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Nordestgaard, B.G.; Langsted, A.; Mora, S.; Kolovou, G.; Baum, H.; Bruckert, E.; Watts, G.F.; Sypniewska, G.; Wiklund, O.; Borén, J.; et al. fasting is not routinely required for determination of a lipid profile: Clinical and laboratory implications including flagging at desirable concentration cut-points—a joint consensus statement from the European Atherosclerosis Society and European Federation of Clinical Chemistry and Laboratory Medicine. Eur. Heart J. 2016, 37, 1944–1958. [Google Scholar] [PubMed] [Green Version]
- Teruya, T.; Chaleckis, R.; Takada, J.; Yanagida, M.; Kondoh, H. Diverse metabolic reactions activated during 58-hr fasting are revealed by non-targeted metabolomic analysis of human blood. Sci. Rep. 2019, 9, 854. [Google Scholar] [CrossRef] [Green Version]
- Pietrocola, F.; Demont, Y.; Castoldi, F.; Enot, D.; Durand, S.; Semeraro, M.; Baracco, E.E.; Pol, J.; Bravo-San Pedro, J.M.; Bordenave, C.; et al. Metabolic effects of fasting on human and mouse blood in vivo. Autophagy 2017, 13, 567–578. [Google Scholar] [CrossRef]
- Beauclercq, S.; Nadal-Desbarats, L.; Hennequet-Antier, C.; Gabriel, I.; Tesseraud, S.; Calenge, F.; Le Bihan-Duval, E.; Mignon-Grasteau, S. Relationships between digestive efficiency and metabolomic profiles of serum and intestinal contents in chickens. Sci. Rep. 2018, 8, 6678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauclercq, S.; Lefevre, A.; Nadal-Desbarats, L.; Germain, K.; Praud, C.; Emond, P.; Bihan-Duval, E.L.; Mignon-Grasteau, S. Does lipidomic serum analysis support the assessment of digestive efficiency in chickens? Poult. Sci. 2019, 98, 1425–1431. [Google Scholar] [CrossRef] [PubMed]
- Psychogios, N.; Hau, D.D.; Peng, J.; Guo, A.C.; Mandal, R.; Bouatra, S.; Sinelnikov, I.; Krishnamurthy, R.; Eisner, R.; Gautam, B.; et al. The human serum metabolome. PLoS ONE 2011, 6, e16957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mountzouris, K.C.; Tsitrsikos, P.; Palamidi, I.; Arvaniti, A.; Mohnl, M.; Schatzmayr, G.; Fegeros, K. Effects of probiotic inclusion levels in broiler nutrition on growth performance, nutrient digestibility, plasma immunoglobulins, and cecal microflora composition. Poult. Sci. 2010, 89, 58–67. [Google Scholar] [CrossRef]
- Wang, Y.; Yin, X.; Yin, D.; Lei, Z.; Mahmood, T.; Yuan, J. Antioxidant response and bioavailability of methionine hydroxy analog relative to DL-methionine in broiler chickens. Anim. Nutr. 2019, 5, 241–247. [Google Scholar] [CrossRef]
- Haq, S.; Grondin, J.; Banskota, S.; Khan, W.I. Autophagy: Roles in intestinal mucosal homeostasis and inflammation. J. Biomed. Sci. 2019, 26, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piekarski, A.; Khaldi, S.; Greene, E.; Lassiter, K.; Mason, J.G.; Anthony, N.; Bottje, W.; Dridi, S. Tissue distribution, gender- and genotype-dependent expression of autophagy-related genes in avian species. PLoS ONE 2014, 9, e112449. [Google Scholar] [CrossRef]
- Kadowaki, M.; Razaul Karim, M.; Carpi, A.; Miotto, G. Nutrient control of macroautophagy in mammalian cells. Mol. Asp. Med. 2006, 27, 426–443. [Google Scholar] [CrossRef]
- Yamauchi, K.; Kamisoyama, H.; Isshiki, Y. Effects of fasting and refeeding on structures of the intestinal villi and epithelial cells in White Leghorn hens. Br. Poult. Sci. 1996, 37, 909–921. [Google Scholar] [CrossRef] [PubMed]
- Ra, E.A.; Lee, T.A.; Won Kim, S.; Park, A.; Choi, H.J.; Jang, I.; Kang, S.; Hee Cheon, J.; Cho, J.W.; Eun Lee, J.; et al. TRIM31 promotes Atg5/Atg7-independent autophagy in intestinal cells. Nat. Commun. 2016, 7, 11726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Zhang, J.; Zhao, J.; Ma, N.; Kim, S.W.; Qiao, S.; Ma, X. Autophagy: The last defense against cellular nutritional stress. Adv. Nutr. 2018, 9, 493–504. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Huang, C.; Wu, Y.; Chen, B.; Zhang, W.; Zhang, J. Autophagy protects the blood-brain barrier through regulating the dynamic of claudin-5 in short-term starvation. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef]
- Noblet, J.; Dubois, S.; Lasnier, J.; Warpechowski, M.; Dimon, P.; Carré, B.; van Milgen, J.; Labussière, E. Fasting heat production and metabolic BW in group-housed broilers. Animal 2015, 9, 1138–1144. [Google Scholar] [CrossRef]
- Cosin-Roger, J.; Simmen, S.; Melhem, H.; Atrott, K.; Frey-Wagner, I.; Hausmann, M.; de Vallière, C.; Spalinger, M.R.; Spielmann, P.; Wenger, R.H.; et al. Hypoxia ameliorates intestinal inflammation through NLRP3/mTOR downregulation and autophagy activation. Nat. Commun. 2017, 8, 98. [Google Scholar] [CrossRef] [Green Version]
- Rangan, P.; Choi, I.; Wei, M.; Navarrete, G.; Guen, E.; Brandhorst, S.; Enyati, N.; Pasia, G.; Maesincee, D.; Ocon, V.; et al. Fasting-mimicking diet modulates microbiota and promotes intestinal regeneration to reduce inflammatory bowel disease pathology. Cell Rep. 2019, 26, 2704–2719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanada, T.; Satomi, Y.; Takao, T.; Ohsumi, Y. The amino-terminal region of Atg3 is essential for association with phosphatidylethanolamine in Atg8 lipidation. FEBS Lett. 2009, 583, 1078–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maity, S.; Jannasch, A.; Adamec, J.; Nalepa, T.; Höök, T.O.; Sepúlveda, M.S. Starvation causes disturbance in amino acid and fatty acid metabolism in Diporeia. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2012, 161, 348–355. [Google Scholar] [CrossRef] [Green Version]
- Weir, J.M.; Wong, G.; Barlow, C.K.; Greeve, M.A.; Kowalczyk, A.; Almasy, L.; Comuzzie, A.G.; Mahaney, M.C.; Jowett, J.B.M.; Shaw, J.; et al. Plasma lipid profiling in a large population-based cohort. J. Lipid Res. 2013, 54, 2898–2908. [Google Scholar]
- Shrestha, A.; Müllner, E.; Poutanen, K.; Mykkänen, H.; Moazzami, A.A. Metabolic changes in serum metabolome in response to a meal. Eur. J. Nutr. 2017, 56, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Dashti, M.; Kulik, W.; Hoek, F.; Veerman, E.C.; Peppelenbosch, M.P.; Rezaee, F. A phospholipidomic analysis of all defined human plasma lipoproteins. Sci. Rep. 2011, 1, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonoki, K.; Iwase, M.; Sasaki, N.; Ohdo, S.; Higuchi, S.; Matsuyama, N.; Iida, M. Relations of lysophosphatidylcholine in low-density lipoprotein with serum lipoprotein-associated phospholipase A2, paraoxonase and homocysteine thiolactonase activities in patients with type 2 diabetes mellitus. Diabetes Res. Clin. Pract. 2009, 86, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Daly, C.; Fitzgerald, A.P.; O’Callaghan, P.; Collins, P.; Cooney, M.T.; Graham, I.M. Homocysteine increases the risk associated with hyperlipidaemia. Eur. J. Cardiovasc. Prev. Rehabil. 2009, 16, 150–155. [Google Scholar] [CrossRef]
- Tepperman, B.L.; Soper, B.D. The role of phospholipase A2 in calcium-ionophore-mediated injury to rat gastric mucosal cells. Dig. Dis. Sci. 1999, 44, 494–502. [Google Scholar] [CrossRef]
- Nishina, A.; Kimura, H.; Sekiguchi, A.; Fukumoto, R.; Nakajima, S.; Furukawa, S. Lysophosphatidylethanolamine in Grifola frondosa as a neurotrophic activator via activation of MAPK. J. Lipid Res. 2006, 47, 1434–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchida, H.; Nakajima, Y.; Ohtake, K.; Ito, J.; Morita, M.; Kamimura, A.; Kobayashi, J. Protective effects of oral glutathione on fasting-induced intestinal atrophy through oxidative stress. World J. Gastroenterol. 2017, 23, 6650–6664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, D.; Witt, S.N. Ethanolamine and phosphatidylethanolamine: Partners in health and disease. Oxidative Med. Cell. Longev. 2017, 2017, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagan, V.E.; Mao, G.; Qu, F.; Angeli, J.P.F.; Doll, S.; Croix, C.S.; Dar, H.H.; Liu, B.; Tyurin, V.A.; Ritov, V.B.; et al. Oxidized arachidonic and adrenic PEs navigate cells to ferroptosis. Nat. Chem. Biol. 2017, 13, 81–90. [Google Scholar] [CrossRef]
- Doll, S.; Proneth, B.; Tyurina, Y.Y.; Panzilius, E.; Kobayashi, S.; Ingold, I.; Irmler, M.; Beckers, J.; Aichler, M.; Walch, A.; et al. ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition. Nat. Chem. Biol. 2017, 13, 91–98. [Google Scholar] [CrossRef]
- Consortium, T.U. UniProt: A hub for protein information. Nucleic Acids Res. 2015, 43, D204–D212. [Google Scholar] [CrossRef]
- Pitari, G.; Malergue, F.; Martin, F.; Philippe, J.M.; Massucci, M.T.; Chabret, C.; Maras, B.; Dupre, S.; Naquet, P.; Galland, F. Pantetheinase activity of membrane-bound Vanin-1: Lack of free cysteamine in tissues of Vanin-1 deficient mice. FEBS Lett. 2000, 483, 149–154. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Gao, Y.; Ma, Z.; Li, Z.; Tang, C.; Qin, X.; Zhang, Z.; Wang, G.; Du, L.; Li, M. Bioluminescent probe for detection of starvation-induced pantetheinase upregulation. Anal. Chem. 2018, 90, 9545–9550. [Google Scholar] [CrossRef]
- Daugherty, M.; Polanuyer, B.; Farrell, M.; Scholle, M.; Lykidis, A.; de Crécy-Lagard, V.; Osterman, A. Complete reconstitution of the human coenzyme A biosynthetic pathway via comparative genomics. J. Biol. Chem. 2002, 277, 21431–21439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahill, J.G.F. Fuel metabolism in starvation. Annu. Rev. Nutr. 2006, 26, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, X.; Zhu, X.; Chen, S.; Zhang, Z.; Zeng, Q.; Deng, L.; Peng, J.; Yu, J.; Wang, L.; Wang, S.; et al. Differential gene expression pattern in hypothalamus of chickens during fasting-induced metabolic reprogramming: Functions of glucose and lipid metabolism in the feed intake of chickens. Poult. Sci. 2014, 93, 2841–2854. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Zhou, K.; Zhang, Y.; Han, X.; Zhao, A.; Liu, J.; Qu, C.; Ge, K.; Huang, F.; Hernandez, B.; et al. Food withdrawal alters the gut microbiota and metabolome in mice. FASEB J. 2018, 32, 4878–4888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordestgaard, B.G. A test in context: Lipid profile, fasting versus non-fasting. J. Am. Coll. Cardiol. 2017, 70, 1637–1646. [Google Scholar] [CrossRef]
- Langsted, A.; Nordestgaard, B.G. Nonfasting versus fasting lipid profile for cardiovascular risk prediction. Pathology 2019, 51, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Whelan, J.; Fritsche, K. Linoleic Acid. Adv. Nutr. 2013, 4, 311–312. [Google Scholar] [CrossRef]
- Yan, S.; Zhang, Q.; Zhong, X.; Tang, J.; Wang, Y.; Yu, J.; Zhou, Y.; Zhang, J.; Guo, F.; Liu, Y.; et al. I prostanoid receptor-mediated inflammatory pathway promotes hepatic gluconeogenesis through activation of PKA and inhibition of AKT. Diabetes 2014, 63, 2911–2923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Yan, S.; Xiao, B.; Zuo, S.; Zhang, Q.; Chen, G.; Yu, Y.; Chen, D.; Liu, Q.; Liu, Y.; et al. Prostaglandin F2alpha facilitates hepatic glucose production through CaMKIIgamma/p38/FOXO1 signaling pathway in fasting and obesity. Diabetes 2018, 67, 1748–1760. [Google Scholar] [CrossRef] [Green Version]
- Arisaka, M.; Arisaka, O.; Fukuda, Y.; Yamashiro, Y. Prostaglandin metabolism in children with diabetes mellitus. I. plasma prostaglandin E2, F2α, TXB2, and serum fatty acid levels. J. Pediatr. Gastr. Nutr. 1986, 5, 878–882. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.H.; Zhang, Y.; Cui, J.; Liu, Y.; McAllan, B.M.; Liao, C.C.; Zhang, S. Adaptation of phenylalanine and tyrosine catabolic pathway to hibernation in bats. PLoS ONE 2013, 8, e62039. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Composition |
|---|---|
| Ingredient (%) | |
| Corn | 50.70 |
| Soybean meal (44% CP 2) | 41.60 |
| Soybean oil | 3.30 |
| Limestone | 1.35 |
| Di-calcium phosphate | 1.43 |
| Sodium chloride | 0.35 |
| Vitamins premix 3 | 0.03 |
| Mineral premix 4 | 0.20 |
| Phytase | 0.02 |
| Choline chloride (60%) | 0.20 |
| DL-Methionine (98%) | 0.30 |
| Antioxidant | 0.02 |
| Titanium dioxide | 0.50 |
| Analyzed nutrient content | |
| Gross energy (Mcal/kg) | 4.03 |
| Crude Protein (%) | 21.99 |
| Ether extract | 6.00 |
| Total starch | 39.12 |
| Calcium (%) | 1.15 |
| Total phosphorus (%) | 0.56 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Xu, Y.; Wu, Y.; Mahmood, T.; Chen, J.; Guo, X.; Wu, W.; Wang, B.; Guo, Y.; Yuan, J. Impact of Different Durations of Fasting on Intestinal Autophagy and Serum Metabolome in Broiler Chicken. Animals 2021, 11, 2183. https://doi.org/10.3390/ani11082183
Wang Y, Xu Y, Wu Y, Mahmood T, Chen J, Guo X, Wu W, Wang B, Guo Y, Yuan J. Impact of Different Durations of Fasting on Intestinal Autophagy and Serum Metabolome in Broiler Chicken. Animals. 2021; 11(8):2183. https://doi.org/10.3390/ani11082183
Chicago/Turabian StyleWang, Youli, Yanwei Xu, Yuqin Wu, Tahir Mahmood, Jing Chen, Xiaorui Guo, Wei Wu, Bo Wang, Yuming Guo, and Jianmin Yuan. 2021. "Impact of Different Durations of Fasting on Intestinal Autophagy and Serum Metabolome in Broiler Chicken" Animals 11, no. 8: 2183. https://doi.org/10.3390/ani11082183
APA StyleWang, Y., Xu, Y., Wu, Y., Mahmood, T., Chen, J., Guo, X., Wu, W., Wang, B., Guo, Y., & Yuan, J. (2021). Impact of Different Durations of Fasting on Intestinal Autophagy and Serum Metabolome in Broiler Chicken. Animals, 11(8), 2183. https://doi.org/10.3390/ani11082183