
Hepatitis E Virus RNA Presence in Wild Boar Carcasses at Slaughterhouses in Italy


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples Collection
2.2. RNA Extraction
2.3. Generation of HEV Standard Curve
2.4. RT-qPCR
2.5. PCR
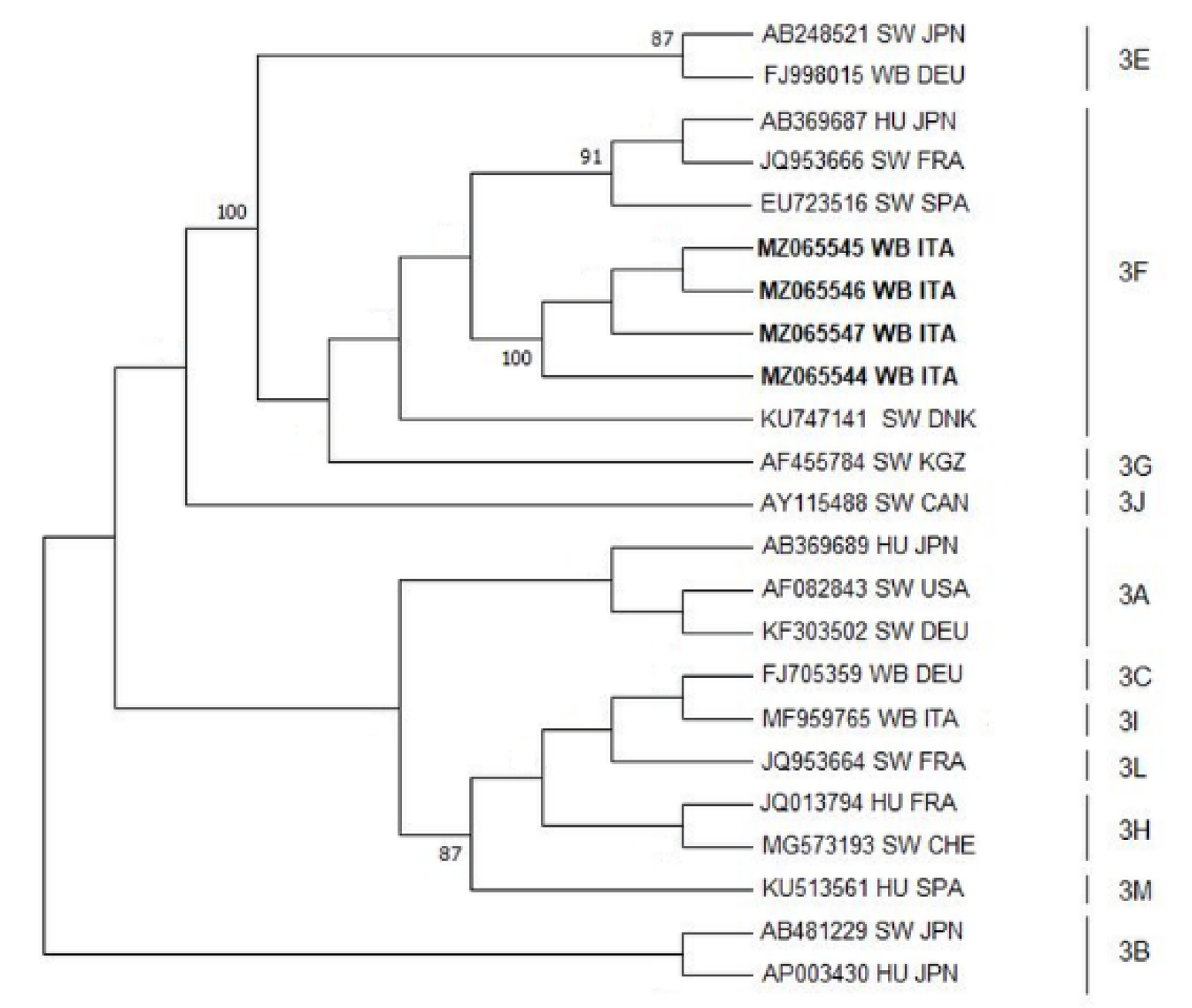
2.6. Sequence and Phylogenetic Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Rein, D.B.; Stevens, G.A.; Theaker, J.; Wittenborn, J.S.; Wiersma, S.T. The global burden of hepatitis E virus genotypes 1 and 2 in 2005. Hepatology 2012, 55, 988–997. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Izopet, J.; Nicot, F.; Simmonds, P.; Jameel, S.; Meng, X.J.; Norder, H.; Okamoto, H.; van der Poel, W.H.M.; Reuter, G.; et al. Update: Proposed reference sequences for subtypes of hepatitis E virus (species Orthohepevirus A). J. Gen. Virol. 2020, 101, 692–698. [Google Scholar] [CrossRef]
- Yin, X.; Li, X.; Feng, Z. Role of envelopment in the HEV life cycle. Viruses 2016, 8, 229. [Google Scholar] [CrossRef]
- Donnelly, M.C.; Scobie, L.; Crossan, C.L.; Dalton, H.; Hayes, P.C.; Simpson, K.J. Review article: Hepatitis E-a concise review of virology, epidemiology, clinical presentation and therapy. Aliment. Pharmacol. Ther. 2017, 46, 126–141. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, R. The Global Prevalence of Hepatitis E Virus Infection and Susceptibility: A Systematic Review; WHO Document Production Services: Geneva, Switzerland, 2010. [Google Scholar]
- Doceul, V.; Bagdassarian, E.; Demange, A.; Pavio, N. Zoonotic hepatitis E virus: Classification, animal reservoirs and transmission routes. Viruses 2016, 8, 270. [Google Scholar] [CrossRef]
- Hartl, J.; Otto, B.; Madden, R.G.; Webb, G.; Woolson, K.L.; Kriston, L.; Vettorazzi, E.; Lohse, A.W.; Dalton, H.R.; Pischke, S. Hepatitis E seroprevalence in Europe: A meta-analysis. Viruses 2016, 8, 211. [Google Scholar] [CrossRef] [PubMed]
- Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernandez Escamez, P.S.; Herman, L.; Koutsoumanis, K.; Lindqvist, R.; Nørrung, B.; et al. Public health risks associated with hepatitis E virus (HEV) as a food-borne pathogen. EFSA J. 2017, 15. [Google Scholar] [CrossRef]
- Ferri, G.; Vergara, A. Hepatitis E Virus in the Food of Animal Origin. Foodborne Pathog. Dis. 2021. [Google Scholar] [CrossRef]
- Lange, H.; Overbo, J.; Borgen, K.; Dudman, S.; Hoddevik, G.; Urdahl, A.M.; Vold, L.; Sjurseth, S.K. Hepatitis e in Norway: Seroprevalence in humans and swine. Epidemiol. Infect. 2017, 145, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, J.; Mesquita, J.R.; Pereira, S.S.; Oliveira, R.M.S.; Abreu-Silva, J.; Rodrigues, A.; Myrmel, M.; Stene-Johansen, K.; Overbo, J.; Goncalves, G.; et al. Prevalence of hepatitis E virus antibodies in workers occupationally exposed to swine in Portugal. Med. Microbiol. Immunol. 2017, 206, 77–81. [Google Scholar] [CrossRef] [PubMed]
- van Gageldonk-Lafeber, A.B.; van der Hoek, W.; Borlée, F.; Heederik, D.J.J.; Mooi, S.H.; Maassen, C.B.M.; Yzermans, C.J.; Rockx, B.; Smit, L.A.M.; Reimerink, J.H.J. Hepatitis E virus seroprevalence among the general population in a livestock-dense area in the Netherlands: A cross-sectional population-based serological survey. BMC Infect. Dis. 2017, 17, 21. [Google Scholar] [CrossRef] [PubMed]
- Di Bartolo, I.; Ponterio, E.; Castellini, L.; Ostanello, F.; Ruggeri, F.M. Viral and antibody HEV prevalence in swine at slaughterhouse in Italy. Vet. Microbiol. 2011, 149, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Berto, A.; Martelli, F.; Grierson, S.; Banks, M. Hepatitis e virus in pork food chain, United Kingdom, 2009–2010. Emerg. Infect. Dis. 2012, 18, 1358–1360. [Google Scholar] [CrossRef] [PubMed]
- Di Bartolo, I.; Diez-Valcarce, M.; Vasickova, P.; Kralik, P.; Hernandez, M.; Angeloni, G.; Ostanello, F.; Bouwknegt, M.; Rodríguez-Lázaro, D.; Pavlik, I.; et al. Hepatitis E virus in pork production chain in Czech Republic, Italy, and Spain, 2010. Emerg. Infect. Dis. 2012, 18, 1282–1289. [Google Scholar] [CrossRef] [PubMed]
- Chelli, E.; Suffredini, E.; De Santis, P.; De Medici, D.; Di Bella, S.; D’amato, S.; Gucciardi, F.; Guercio, A.; Ostanello, F.; Perrone, V.; et al. Hepatitis E Virus Occurrence in Pigs Slaughtered in Italy. Animals 2021, 11, 277. [Google Scholar] [CrossRef]
- Sooryanarain, H.; Heffron, C.L.; Hill, D.E.; Fredericks, J.; Rosenthal, B.M.; Werre, S.R.; Opriessnig, T.; Meng, X.J. Hepatitis e virus in pigs from slaughterhouses, United States, 2017–2019. Emerg. Infect. Dis. 2020, 26, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Intharasongkroh, D.; Sa-nguanmoo, P.; Tuanthap, S.; Thongmee, T.; Duang-in, A.; Klinfueng, S.; Chansaenroj, J.; Vongpunsawad, S.; Theamboonlers, A.; Payungporn, S.; et al. Hepatitis E Virus in Pork and Variety Meats Sold in Fresh Markets. Food Environ. Virol. 2017, 9, 45–53. [Google Scholar] [CrossRef]
- Leblanc, D.; Poitras, E.; Gagné, M.J.; Ward, P.; Houde, A. Hepatitis E virus load in swine organs and tissues at slaughterhouse determined by real-time RT-PCR. Int. J. Food Microbiol. 2010, 139, 206–209. [Google Scholar] [CrossRef]
- Nantel-Fortier, N.; Letellier, A.; Lachapelle, V.; Fravalo, P.; L’Homme, Y.; Brassard, J. Detection and Phylogenetic Analysis of the Hepatitis E Virus in a Canadian Swine Production Network. Food Environ. Virol. 2016, 8, 296–304. [Google Scholar] [CrossRef]
- Wichmann, O.; Schimanski, S.; Koch, J.; Kohler, M.; Rothe, C.; Plentz, A.; Jilg, W.; Stark, K. Phylogenetic and case-control study on hepatitis E virus infection in Germany. J. Infect. Dis. 2008, 198, 1732–1741. [Google Scholar] [CrossRef] [PubMed]
- Kozyra, I.; Jabłoński, A.; Bigoraj, E.; Rzeżutka, A. Wild boar as a sylvatic reservoir of hepatitis E virus in Poland: A cross-sectional population study. Viruses 2020, 12, 1113. [Google Scholar] [CrossRef] [PubMed]
- Izopet, J.; Tremeaux, P.; Marion, O.; Migueres, M.; Capelli, N.; Chapuy-Regaud, S.; Mansuy, J.M.; Abravanel, F.; Kamar, N.; Lhomme, S. Hepatitis E virus infections in Europe. J. Clin. Virol. 2019, 120, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Juarez, A.; Frias, M.; Martinez-Peinado, A.; Risalde, M.A.; Rodriguez-Cano, D.; Camacho, A.; García-Bocanegra, I.; Cuenca-Lopez, F.; Gomez-Villamandos, J.C.; Rivero, A. Familial Hepatitis E Outbreak Linked to Wild Boar Meat Consumption. Zoonoses Public Health 2017, 64, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Sommerkorn, F.M.; Schauer, B.; Schreiner, T.; Fickenscher, H.; Krumbholz, A. Performance of Hepatitis E Virus (HEV)-antibody tests: A comparative analysis based on samples from individuals with direct contact to domestic pigs or wild boar in Germany. Med. Microbiol. Immunol. 2017, 206, 277–286. [Google Scholar] [CrossRef]
- Di Pasquale, S.; De Santis, P.; La Rosa, G.; Di Domenico, K.; Iaconelli, M.; Micarelli, G.; Martini, E.; Bilei, S.; De Medici, D.; Suffredini, E. Quantification and genetic diversity of Hepatitis E virus in wild boar (Sus scrofa) hunted for domestic consumption in Central Italy. Food Microbiol. 2019, 82, 194–201. [Google Scholar] [CrossRef]
- Mazzei, M.; Nardini, R.; Verin, R.; Forzan, M.; Poli, A.; Tolari, F. Serologic and molecular survey for hepatitis E virus in wild boar (Sus scrofa) in Central Italy. New Microbes New Infect. 2015, 7, 41–47. [Google Scholar] [CrossRef]
- Dalton, H.R.; Seghatchian, J. Hepatitis E virus: Emerging from the shadows in developed countries. Transfus. Apher. Sci. 2016, 55, 271–274. [Google Scholar] [CrossRef] [PubMed]
- de Deus, N.; Peralta, B.; Pina, S.; Allepuz, A.; Mateu, E.; Vidal, D.; Ruiz-Fons, F.; Martín, M.; Gortázar, C.; Segalés, J. Epidemiological study of hepatitis E virus infection in European wild boars (Sus scrofa) in Spain. Vet. Microbiol. 2008, 129, 163–170. [Google Scholar] [CrossRef]
- Boadella, M.; Ruiz-Fons, J.F.; Vicente, J.; Martín, M.; Segalés, J.; Gortazar, C. Seroprevalence evolution of selected pathogens in iberian wild boar. Transbound. Emerg. Dis. 2012, 59, 395–404. [Google Scholar] [CrossRef]
- Lewis, H.C.; Wichmann, O.; Duizer, E. Transmission routes and risk factors for autochthonous hepatitis E virus infection in Europe: A systematic review. Epidemiol. Infect. 2010, 138, 145–166. [Google Scholar] [CrossRef] [PubMed]
- Schielke, A.; Ibrahim, V.; Czogiel, I.; Faber, M.; Schrader, C.; Dremsek, P.; Ulrich, R.G.; Johne, R. Hepatitis E virus antibody prevalence in hunters from a district in Central Germany, 2013: A cross-sectional study providing evidence for the benefit of protective gloves during disembowelling of wild boars. BMC Infect. Dis. 2015, 15, 1–8. [Google Scholar] [CrossRef]
- Sáez-Royuela, C.; Gomariz, R.P.L.T. Age Determination of European Wild Boar Wildl. Soc. Bull. 1989, 17, 326–329. [Google Scholar]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods 2006, 131, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, H.; Ling, R.; Li, H.; Harrison, T.J. The complete sequence of hepatitis E virus genotype 4 reveals an alternative strategy for translation of open reading frames 2 and 3. J. Gen. Virol. 2000, 81, 1675–1686. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Pittiglio, C.; Khomenko, S.; Beltran-Alcrudo, D. Wild boar mapping using population-density statistics: From polygons to high resolution raster maps. PLoS ONE 2018, 13, 1–19. [Google Scholar] [CrossRef]
- Mughini-Gras, L.; Angeloni, G.; Salata, C.; Vonesch, N.; D’Amico, W.; Campagna, G.; Natale, A.; Zuliani, F.; Ceglie, L.; Monne, I.; et al. Hepatitis e virus infection in North Italy: High seroprevalence in swine herds and increased risk for swine workers. Epidemiol. Infect. 2017, 145, 3375–3384. [Google Scholar] [CrossRef] [PubMed]
- Spada, E.; Pupella, S.; Pisani, G.; Bruni, R.; Chionne, P.; Madonna, E.; Villano, U.; Simeoni, M.; Fabi, S.; Marano, G.; et al. A nationwide retrospective study on prevalence of hepatitis E virus infection in Italian blood donors. Blood Transfus. 2018, 16, 413–421. [Google Scholar]



{kind=link}
{kind=link}
| Animal | Age Category | Sex | Hunting Area | Slaughtering Date | RT-qPCR Viral Load | |
|---|---|---|---|---|---|---|
| Liver | Swab | |||||
| 1 | Adult | F | Semproniano GR | 11 February 2021 | 4.28 × 102 | - |
| 2 | Adult | F | 1.52 × 101 | - | ||
| 3 | Adult | M | 1.13 × 101 | - | ||
| 4 | Adult | M | 5.37 × 101 | - | ||
| 5 | Adult | F | Orbetello GR | 19 February 2021 | - | 3.84 × 101 |
| 6 | Adult | F | - | 9.9 × 102 | ||
| 7 | Sub-adult | M | Roccalbegna GR | 25 February 2021 | 1.31 × 103 | 1.42 × 102 |
| 8 | Sub-adult | M | 6.12 × 102 | 5.16 × 103 | ||
| 9 | Adult | F | 7.33 × 103 | - | ||
| 10 | Adult | M | Vicchi PO | 31 February 2021 | 2.08 × 103 | - |
| Mean | 1.48 × 103 | 1.57 × 103 | ||||
| Standard Deviation | 2.47 × 103 | 2.42 × 103 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Forzan, M.; Pacini, M.I.; Periccioli, M.; Mazzei, M. Hepatitis E Virus RNA Presence in Wild Boar Carcasses at Slaughterhouses in Italy. Animals 2021, 11, 1624. https://doi.org/10.3390/ani11061624
Forzan M, Pacini MI, Periccioli M, Mazzei M. Hepatitis E Virus RNA Presence in Wild Boar Carcasses at Slaughterhouses in Italy. Animals. 2021; 11(6):1624. https://doi.org/10.3390/ani11061624
Chicago/Turabian StyleForzan, Mario, Maria Irene Pacini, Marcello Periccioli, and Maurizio Mazzei. 2021. "Hepatitis E Virus RNA Presence in Wild Boar Carcasses at Slaughterhouses in Italy" Animals 11, no. 6: 1624. https://doi.org/10.3390/ani11061624
APA StyleForzan, M., Pacini, M. I., Periccioli, M., & Mazzei, M. (2021). Hepatitis E Virus RNA Presence in Wild Boar Carcasses at Slaughterhouses in Italy. Animals, 11(6), 1624. https://doi.org/10.3390/ani11061624