
Health Monitoring of Laboratory Rodent Colonies—Talking about (R)evolution


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
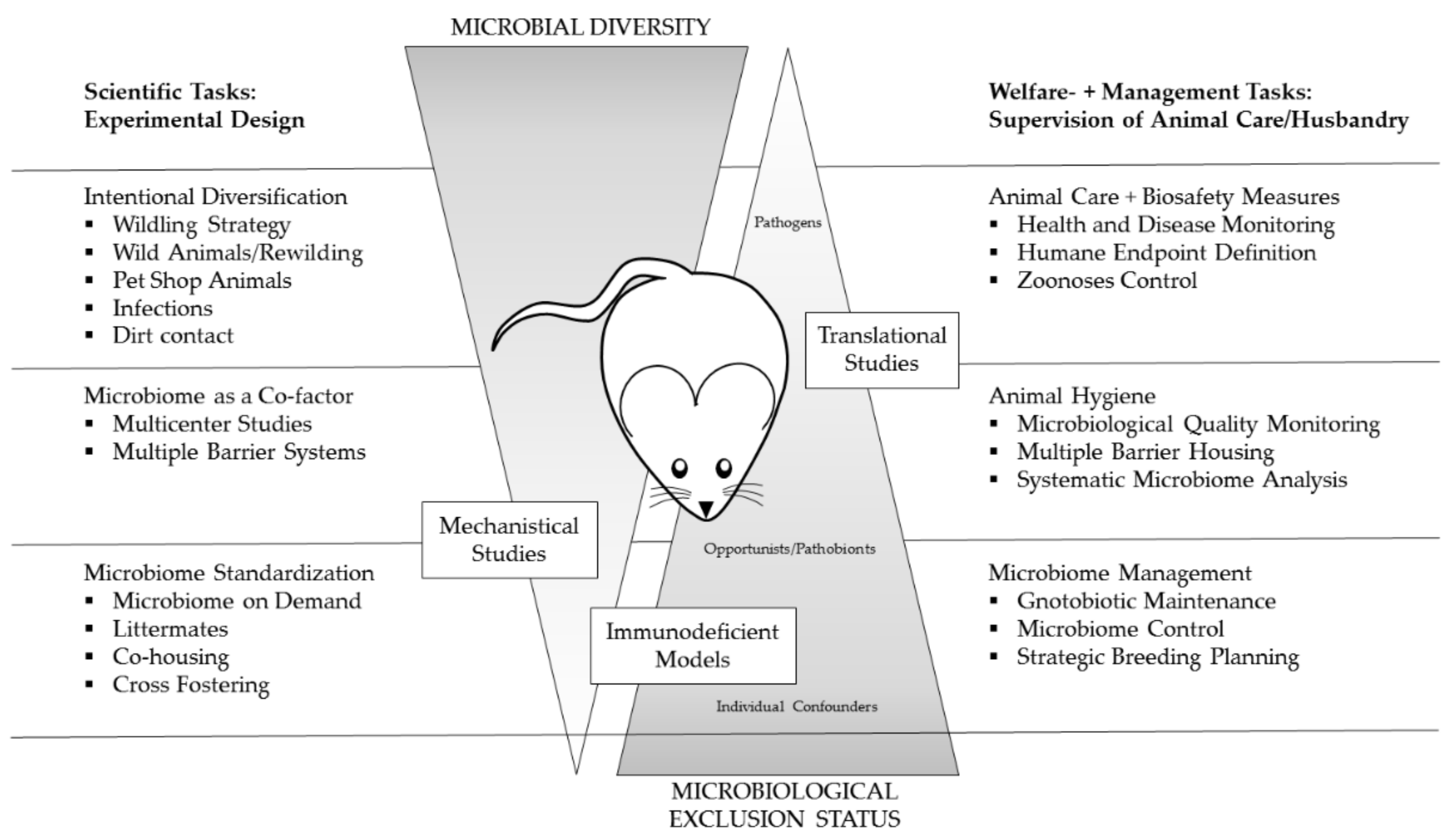
2. Scope of Health Monitoring Management
3. How to Keep Them Alive
3.1. Microbial Agents Compromising the Animal´s Life
3.2. Diagnostic Measures to Enable Disease Control
4. How to Keep Them Healthy
4.1. Agents Affecting the Animal’s Health Status
4.2. Hygienic Measures to Maintain the Health Status of Laboratory Rodents
5. How to Ensure Quality
5.1. Microbial Agents Impacting “Quality”
5.2. Measures to Ensure Quality—Standardized Health Monitoring
5.3. Key Figures of Standard Health Monitoring
5.4. Recent Developments in Standard Health Monitoring
6. How to Ensure Validity
6.1. Microbial Impact on Model Phenotypes and Research Validity
6.2. Measure to Ensure “Validity”
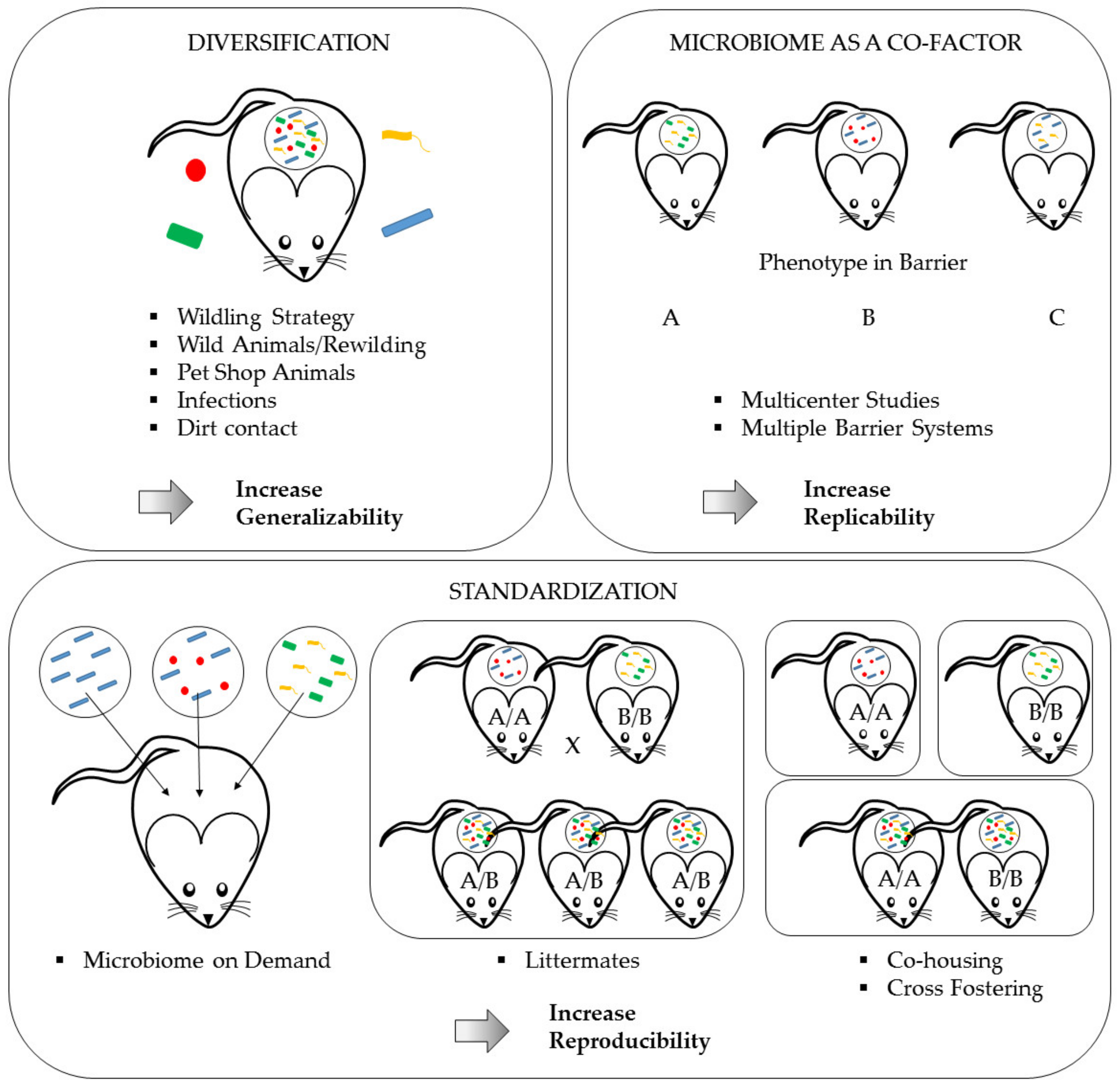
6.2.1. Standardizing the Microbiome
Littermates
Co-Housing and Cross-Fostering
Microbiome on Demand—Gnotobiology
6.2.2. Intentional Diversification of the Microbiome—Re-Wilding, Wildlings, Pet-Shop or Wild Animals
6.2.3. Microbiome as a Co-Factor
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Baker, D.G. Natural pathogens of laboratory mice, rats, and rabbits and their effects on research. Clin. Microbiol. Rev. 1998, 11, 231–266. [Google Scholar] [CrossRef]
- Weisbroth, S.H. Post-indigenous disease: Changing concepts of disease in laboratory rodents. Lab. Anim. 1996, 25, 25–33. [Google Scholar]
- Hansen, A.K.; Franklin, C. Microbiota, laboratory animals, and research. Lab. Anim. 2019, 53, 229–231. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.R.; Olekszak, H. An overview of typical infections of research mice: Health monitoring and prevention of infection. Curr. Protoc. Mouse Biol. 2015, 5, 235–245. [Google Scholar] [CrossRef]
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique; Universities Federation for Animal Welfare: Wheathampstead, UK, 1959. [Google Scholar]
- Ericsson, A.C.; Davis, J.W.; Spollen, W.; Bivens, N.; Givan, S.; Hagan, C.E.; McIntosh, M.; Franklin, C.L. Effects of vendor and genetic background on the composition of the fecal microbiota of inbred mice. PLoS ONE 2015, 10, e0116704. [Google Scholar] [CrossRef]
- Ericsson, A.C.; Gagliardi, J.; Bouhan, D.; Spollen, W.G.; Givan, S.A.; Franklin, C.L. The influence of caging, bedding, and diet on the composition of the microbiota in different regions of the mouse gut. Sci. Rep. 2018, 8, 4065. [Google Scholar] [CrossRef] [PubMed]
- Rausch, P.; Basic, M.; Batra, A.; Bischoff, S.C.; Blaut, M.; Clavel, T.; Glasner, J.; Gopalakrishnan, S.; Grassl, G.A.; Gunther, C.; et al. Analysis of factors contributing to variation in the c57bl/6j fecal microbiota across german animal facilities. Int. J. Med. Microbiol. 2016, 306, 343–355. [Google Scholar] [CrossRef]
- Bleich, A.; Fox, J.G. The mammalian microbiome and its importance in laboratory animal research. ILAR J. 2015, 56, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Bleich, A.; Hansen, A.K. Time to include the gut microbiota in the hygienic standardisation of laboratory rodents. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 81–92. [Google Scholar] [CrossRef]
- Hansen, A.K.; Hansen, C.H.; Krych, L.; Nielsen, D.S. Impact of the gut microbiota on rodent models of human disease. World J. Gastroenterol. 2014, 20, 17727–17736. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.K.; Krych, L.; Nielsen, D.S.; Hansen, C.H. A review of applied aspects of dealing with gut microbiota impact on rodent models. ILAR J. 2015, 56, 250–264. [Google Scholar] [CrossRef] [PubMed]
- Masopust, D.; Sivula, C.P.; Jameson, S.C. Of mice, dirty mice, and men: Using mice to understand human immunology. J. Immunol. 2017, 199, 383–388. [Google Scholar] [CrossRef]
- Strech, D.; Dirnagl, U. 3rs missing: Animal research without scientific value is unethical. BMJ Open Sci. 2019, 3, e000048. [Google Scholar] [CrossRef]
- Kraft, V.; Deeny, A.; Blanchet, H.; Hansen, A. Report of the felasa working group on animal health. Recommendations for the health monitoring of mouse, rat, hamster, guineapig and rabbit breeding colonies. Lab. Anim. 1994, 28, 1–12. [Google Scholar] [CrossRef]
- Nicklas, W.; Baneux, P.; Boot, R.; Decelle, T.; Deeny, A.A.; Fumanelli, M.; Illgen-Wilcke, B.; FELASA. FELASA Recommendations for the health monitoring of rodent and rabbit colonies in breeding and experimental units. Lab. Anim. 2002, 36, 20–42. [Google Scholar] [CrossRef]
- Mähler, M.; Berard, M.; Feinstein, R.; Gallagher, A.; Illgen-Wilcke, B.; Pritchett-Corning, K.; Raspa, M. Felasa recommendations for the health monitoring of mouse, rat, hamster, guinea pig and rabbit colonies in breeding and experimental units. Lab. Anim. 2014, 48, 178–192. [Google Scholar]
- Barthold, S.W.; Griffey, S.M.; Percy, D.H. Pathology of Laboratory Rodents and Rabbits; Wiley-Blackwell: Hoboken, NJ, USA, 2016; Volume 4. [Google Scholar]
- Fox, J.A.L.; Glen, O.; Kathleen, P.; Mark, W. Laboratory Animal Medicine; Academic Press: Cambridge, MA, USA, 2015; Volume 3. [Google Scholar]
- Lussier, G. Potential detrimental effects of rodent viral infections on long-term experiments. Vet. Res. Commun. 1988, 12, 199–217. [Google Scholar] [CrossRef]
- Tyzzer, E.E. A fatal disease of the japanese waltzing mouse caused by a spore-bearing bacillus (bacillus piliformis, n. Sp.). J. Med. Res. 1917, 37, 307–338. [Google Scholar] [PubMed]
- Ganaway, J.R.; Allen, A.M.; Moore, T.D. Tyzzer’s disease. Am. J. Pathol. 1971, 64, 717–730. [Google Scholar]
- Itoh, T.; Ebukuro, M.; Kagiyama, N. Inactivation of bacillus piliformis spores by heat and certain chemical disinfectants. Jikken Dobutsu 1987, 36, 239–244. [Google Scholar]
- Ganaway, J.R. Effect of heat and selected chemical disinfectants upon infectivity of spores of bacillus piliformis (tyzzer’s disease). Lab. Anim. Sci. 1980, 30, 192–196. [Google Scholar] [PubMed]
- Hansen, A.K.; Svendsen, O.; Mollegaard-Hansen, K.E. Epidemiological studies of bacillus piliformis infection and tyzzer’s disease in laboratory rats. Z. Vers. 1990, 33, 163–169. [Google Scholar]
- Smith, T. Some bacteriological and environmental factors in the pneumonias of lower animals with special reference to the guinea-pig. J. Med. Res. 1913, 29, 291–324. [Google Scholar] [PubMed]
- Collins, M.J., Jr.; Parker, J.C. Murine virus contaminants of leukemia viruses and transplantable tumors. J. Natl. Cancer Inst. 1972, 49, 1139–1143. [Google Scholar]
- Nicklas, W.; Kraft, V.; Meyer, B. Contamination of transplantable tumors, cell lines, and monoclonal antibodies with rodent viruses. Lab. Anim. Sci. 1993, 43, 296–300. [Google Scholar] [PubMed]
- Nakai, N.; Kawaguchi, C.; Nawa, K.; Kobayashi, S.; Katsuta, Y.; Watanabe, M. Detection and elimination of contaminating microorganisms in transplantable tumors and cell lines. Exp. Anim. 2000, 49, 309–313. [Google Scholar] [CrossRef][Green Version]
- Livingston, R.S. In Testing Biological—Mitigating Risk to Animals, People and Research. In Proceedings of the ESLAV-ECLAM Virtual Annual Meeting 2020, Online, 24–25 November 2020. [Google Scholar]
- Lipman, N.S.; Perkins, S.; Nguyen, H.; Pfeffer, M.; Meyer, H. Mousepox resulting from use of ectromelia virus-contaminated, imported mouse serum. Comp. Med. 2000, 50, 426–435. [Google Scholar]
- Dick, E.J., Jr.; Kittell, C.L.; Meyer, H.; Farrar, P.L.; Ropp, S.L.; Esposito, J.J.; Buller, R.M.; Neubauer, H.; Kang, Y.H.; McKee, A.E. Mousepox outbreak in a laboratory mouse colony. Lab. Anim. Sci. 1996, 46, 602–611. [Google Scholar]
- Labelle, P.; Hahn, N.E.; Fraser, J.K.; Kendall, L.V.; Ziman, M.; James, E.; Shastri, N.; Griffey, S.M. Mousepox detected in a research facility: Case report and failure of mouse antibody production testing to identify ectromelia virus in contaminated mouse serum. Comp. Med. 2009, 59, 180–186. [Google Scholar] [PubMed]
- Dykewicz, C.A.; Dato, V.M.; Fisher-Hoch, S.P.; Howarth, M.V.; Perez-Oronoz, G.I.; Ostroff, S.M.; Gary, H., Jr.; Schonberger, L.B.; McCormick, J.B. Lymphocytic choriomeningitis outbreak associated with nude mice in a research institute. JAMA 1992, 267, 1349–1353. [Google Scholar] [CrossRef] [PubMed]
- Clement, J.; LeDuc, J.W.; Lloyd, G.; Reynes, J.M.; McElhinney, L.; van Ranst, M.; Lee, H.W. Wild rats, laboratory rats, pet rats: Global seoul hantavirus disease revisited. Viruses 2019, 11, 652. [Google Scholar] [CrossRef] [PubMed]
- Umenai, T.; Lee, H.W.; Lee, P.W.; Saito, T.; Toyoda, T.; Hongo, M.; Yoshinaga, K.; Nobunaga, T.; Horiuchi, T.; Ishida, N. Korean haemorrhagic fever in staff in an animal laboratory. Lancet 1979, 1, 1314–1316. [Google Scholar] [CrossRef]
- De Souza, M.; Smith, A.L. Comparison of isolation in cell culture with conventional and modified mouse antibody production tests for detection of murine viruses. J. Clin. Microbiol. 1989, 27, 185–187. [Google Scholar] [CrossRef]
- Lewis, V.J.; Clayton, D.M. An evaluation of the mouse antibody production test for detecting three murine viruses. Lab. Anim. Sci. 1971, 21, 203–205. [Google Scholar]
- Morse, S.S. Comparative sensitivity of infectivity assay and mouse antibody production (map) test for detection of mouse thymic virus (mtlv). J. Virol. Methods 1990, 28, 15–23. [Google Scholar] [CrossRef]
- Bootz, F.; Sieber, I. Replacement of mouse and rat antibody production test; comparison of sensitivity between the in vitro and in vivo methods. ALTEX 2002, 19 (Suppl. 1), 76–86. [Google Scholar]
- Bauer, B.A.; Besch-Williford, C.L.; Riley, L.K. Comparison of the mouse antibody production (map) assay and polymerase chain reaction (pcr) assays for the detection of viral contaminants. Biologicals 2004, 32, 177–182. [Google Scholar] [CrossRef]
- Mahabir, E.; Jacobsen, K.; Brielmeier, M.; Peters, D.; Needham, J.; Schmidt, J. Mouse antibody production test: Can we do without it? J. Virol. Methods 2004, 120, 239–245. [Google Scholar] [CrossRef]
- Meehan, T.F.; Conte, N.; West, D.B.; Jacobsen, J.O.; Mason, J.; Warren, J.; Chen, C.K.; Tudose, I.; Relac, M.; Matthews, P.; et al. Disease model discovery from 3,328 gene knockouts by the international mouse phenotyping consortium. Nat. Genet. 2017, 49, 1231–1238. [Google Scholar] [CrossRef]
- Brandes, R.P.; Dueck, A.; Engelhardt, S.; Kaulich, M.; Kupatt, C.; de Angelis, M.T.; Leisegang, M.S.; le Noble, F.; Moretti, A.; Muller, O.J.; et al. Dgk and dzhk position paper on genome editing: Basic science applications and future perspective. Basic Res. Cardiol. 2021, 116, 2. [Google Scholar] [CrossRef]
- Waggie, K.S.; Hansen, C.T.; Ganaway, J.R.; Spencer, T.S. A study of mouse strains susceptibility to bacillus piliformis (tyzzer’s disease): The association of b-cell function and resistance. Lab. Anim. Sci. 1981, 31, 139–142. [Google Scholar]
- Compton, S.R.; Ball-Goodrich, L.J.; Johnson, L.K.; Johnson, E.A.; Paturzo, F.X.; Macy, J.D. Pathogenesis of enterotropic mouse hepatitis virus in immunocompetent and immunodeficient mice. Comp. Med. 2004, 54, 681–689. [Google Scholar] [PubMed]
- Van Andel, R.A.; Hook, R.R., Jr.; Franklin, C.L.; Besch-Williford, C.L.; Riley, L.K. Interleukin-12 has a role in mediating resistance of murine strains to tyzzer’s disease. Infect. Immun. 1998, 66, 4942–4946. [Google Scholar] [CrossRef]
- Binder, A.; Gartner, K.; Hedrich, H.J.; Hermanns, W.; Kirchhoff, H.; Wonigeit, K. Strain differences in sensitivity of rats to mycoplasma arthritidis isr 1 infection are under multiple gene control. Infect. Immun. 1990, 58, 1584–1590. [Google Scholar] [CrossRef]
- Brownstein, D.G.; Smith, A.L.; Jacoby, R.O.; Johnson, E.A.; Hansen, G.; Tattersall, P. Pathogenesis of infection with a virulent allotropic variant of minute virus of mice and regulation by host genotype. Lab. Investig. 1991, 65, 357–364. [Google Scholar] [PubMed]
- Fornefett, J.; Krause, J.; Klose, K.; Fingas, F.; Hassert, R.; Eisenberg, T.; Schrodl, W.; Grunwald, T.; Muller, U.; Baums, C.G. Comparative analysis of clinics, pathologies and immune responses in balb/c and c57bl/6 mice infected with streptobacillus moniliformis. Microbes Infect. 2018, 20, 101–110. [Google Scholar] [CrossRef]
- Huber, A.C.; Yolken, R.H.; Mader, L.C.; Strandberg, J.D.; Vonderfecht, S.L. Pathology of infectious diarrhea of infant rats (idir) induced by an antigenically distinct rotavirus. Vet. Pathol. 1989, 26, 376–385. [Google Scholar] [CrossRef]
- Burns, J.W.; Krishnaney, A.A.; Vo, P.T.; Rouse, R.V.; Anderson, L.J.; Greenberg, H.B. Analyses of homologous rotavirus infection in the mouse model. Virology 1995, 207, 143–153. [Google Scholar] [CrossRef]
- Ijaz, M.K.; Sabara, M.I.; Alkarmi, T.; Frenchick, P.J.; Ready, K.F.; Longson, M.; Dar, F.K.; Babiuk, L.A. Characterization of two rotaviruses differing in their in vitro and in vivo virulence. J. Vet. Med. Sci. 1993, 55, 963–971. [Google Scholar] [CrossRef] [PubMed]
- Little, L.M.; Shadduck, J.A. Pathogenesis of rotavirus infection in mice. Infect. Immun. 1982, 38, 755–763. [Google Scholar] [CrossRef]
- Vonderfecht, S.L.; Huber, A.C.; Eiden, J.; Mader, L.C.; Yolken, R.H. Infectious diarrhea of infant rats produced by a rotavirus-like agent. J. Virol. 1984, 52, 94–98. [Google Scholar] [CrossRef]
- Held, N.; Hedrich, H.J.; Bleich, A. Successful sanitation of an edim-infected mouse colony by breeding cessation. Lab. Anim. 2011, 45, 276–279. [Google Scholar] [CrossRef] [PubMed]
- Lindstrom, K.E.; Henderson, K.S.; Mayorga, M.S.; Kuiper, V.A.; Wilkerson, J.D. Contaminated shipping materials identified as the source of rotaviral infection of exported mice. J. Am. Assoc. Lab. Anim. Sci. 2018, 57, 529–533. [Google Scholar] [CrossRef]
- Broderson, J.R.; Lindsey, J.R.; Crawford, J.E. The role of environmental ammonia in respiratory mycoplasmosis of rats. Am. J. Pathol. 1976, 85, 115–130. [Google Scholar] [PubMed]
- Schoeb, T.R.; Kervin, K.C.; Lindsey, J.R. Exacerbation of murine respiratory mycoplasmosis in gnotobiotic f344/n rats by sendai virus infection. Vet. Pathol. 1985, 22, 272–282. [Google Scholar] [CrossRef]
- Ganaway, J.R.; Spencer, T.H.; Moore, T.D.; Allen, A.M. Isolation, propagation, and characterization of a newly recognized pathogen, cilia-associated respiratory bacillus of rats, an etiological agent of chronic respiratory disease. Infect. Immun. 1985, 47, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Kohyama, K.; Takakura, A.; Takenouchi, T.; Kagiyama, N. Naturally occurring car bacillus infection in a laboratory rat colony and epizootiological observations. Jikken Dobutsu 1987, 36, 387–393. [Google Scholar] [PubMed]
- Van Zwieten, M.J.; Solleveld, H.A.; Lindsey, J.R.; de Groot, F.G.; Zurcher, C.; Hollander, C.F. Respiratory disease in rats associated with a filamentous bacterium: A preliminary report. Lab. Anim. Sci. 1980, 30, 215–221. [Google Scholar]
- Brogden, K.A.; Cutlip, R.C.; Lehmkuhl, H.D. Cilia-associated respiratory bacillus in wild rats in central iowa. J. Wildl. Dis. 1993, 29, 123–126. [Google Scholar] [CrossRef]
- Ike, F.; Sakamoto, M.; Ohkuma, M.; Kajita, A.; Matsushita, S.; Kokubo, T. Filobacterium rodentium gen. Nov., sp. Nov., a member of filobacteriaceae fam. Nov. Within the phylum bacteroidetes; includes a microaerobic filamentous bacterium isolated from specimens from diseased rodent respiratory tracts. Int. J. Syst. Evol. Microbiol. 2016, 66, 150–157. [Google Scholar] [CrossRef]
- Adhikary, S.; Bisgaard, M.; Dagnaes-Hansen, F.; Christensen, H. Clonal outbreaks of [pasteurella] pneumotropica biovar heyl in two mouse colonies. Lab. Anim. 2017, 51, 613–621. [Google Scholar] [CrossRef]
- Weisbroth, S.H.; Scher, S.; Boman, I. Pasteurella pneumotropica abscess syndrome in a mouse colony. J. Am. Vet. Med. Assoc. 1969, 155, 1206–1210. [Google Scholar] [PubMed]
- Wagner, J.E.; Garrison, R.G.; Johnson, D.R.; McGuire, T.J. Spontaneous conjunctivitis and dacryoadenitis of mice. J. Am. Vet. Med. Assoc. 1969, 155, 1211–1217. [Google Scholar]
- Needham, J.R.; Cooper, J.E. An eye infection in laboratory mice associated with pasteurella pneumotropica. Lab. Anim. 1975, 9, 197–200. [Google Scholar] [CrossRef]
- Sebesteny, A. Abscesses of the bulbourethral glands of mice due to pasteurella pneumotropica. Lab. Anim. 1973, 7, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Van der Schaaf, A.; Mullink, J.W.; Nikkels, R.J.; Goudswaard, J. Pasteurella pneumotropica as a causal microorganism of multiple subcutaneous abscesses in a colony wistar rats. Z. Vers. 1970, 12, 356–362. [Google Scholar]
- Adhikary, S.; Nicklas, W.; Bisgaard, M.; Boot, R.; Kuhnert, P.; Waberschek, T.; Aalbaek, B.; Korczak, B.; Christensen, H. Rodentibacter gen. Nov. Including rodentibacter pneumotropicus comb. Nov., rodentibacter heylii sp. Nov., rodentibacter myodis sp. Nov., rodentibacter ratti sp. Nov., rodentibacter heidelbergensis sp. Nov., rodentibacter trehalosifermentans sp. Nov., rodentibacter rarus sp. Nov., rodentibacter mrazii and two genomospecies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1793–1806. [Google Scholar] [PubMed]
- Mrochen, D.M.; Grumann, D.; Schulz, D.; Gumz, J.; Trube, P.; Pritchett-Corning, K.; Johnson, S.; Nicklas, W.; Kirsch, P.; Martelet, K.; et al. Global spread of mouse-adapted staphylococcus aureus lineages cc1, cc15, and cc88 among mouse breeding facilities. Int. J. Med. Microbiol. 2018, 308, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Raafat, D.; Mrochen, D.M.; Al’Sholui, F.; Heuser, E.; Ryll, R.; Pritchett-Corning, K.R.; Jacob, J.; Walther, B.; Matuschka, F.R.; Richter, D.; et al. Molecular epidemiology of methicillin-susceptible and methicillin-resistant staphylococcus aureus in wild, captive and laboratory rats: Effect of habitat on the nasal s. Aureus population. Toxins 2020, 12, 80. [Google Scholar] [CrossRef]
- Bleich, A.; Kirsch, P.; Sahly, H.; Fahey, J.; Smoczek, A.; Hedrich, H.J.; Sundberg, J.P. Klebsiella oxytoca: Opportunistic infections in laboratory rodents. Lab. Anim. 2008, 42, 369–375. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, C.J.; Pillers, D.A.; Pang, J.; Degagne, J.M.; Kempton, J.B.; Trune, D.R. Gram-negative pathogen klebsiella oxytoca is associated with spontaneous chronic otitis media in toll-like receptor 4-deficient c3h/hej mice. Acta Otolaryngol. 2008, 128, 132–138. [Google Scholar] [CrossRef]
- Clarke, M.C.; Taylor, R.J.; Hall, G.A.; Jones, P.W. The occurrence in mice of facial and mandibular abscesses associated with staphylococcus aureus. Lab. Anim. 1978, 12, 121–123. [Google Scholar] [CrossRef]
- Bridgeford, E.C.; Fox, J.G.; Nambiar, P.R.; Rogers, A.B. Agammaglobulinemia and staphylococcus aureus botryomycosis in a cohort of related sentinel swiss webster mice. J. Clin. Microbiol. 2008, 46, 1881–1884. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fox, J.G.; Shen, Z.; Muthupalani, S.; Rogers, A.R.; Kirchain, S.M.; Dewhirst, F.E. Chronic hepatitis, hepatic dysplasia, fibrosis, and biliary hyperplasia in hamsters naturally infected with a novel helicobacter classified in the h. Bilis cluster. J. Clin. Microbiol. 2009, 47, 3673–3681. [Google Scholar] [CrossRef]
- Fox, J.G. The expanding genus of helicobacter: Pathogenic and zoonotic potential. Semin. Gastrointest. Dis. 1997, 8, 124–141. [Google Scholar] [PubMed]
- Eaton, K.A.; Opp, J.S.; Gray, B.M.; Bergin, I.L.; Young, V.B. Ulcerative typhlocolitis associated with helicobacter mastomyrinus in telomerase-deficient mice. Vet. Pathol. 2011, 48, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Shomer, N.H.; Dangler, C.A.; Schrenzel, M.D.; Fox, J.G. Helicobacter bilis-induced inflammatory bowel disease in scid mice with defined flora. Infect. Immun. 1997, 65, 4858–4864. [Google Scholar] [CrossRef] [PubMed]
- Mangerich, A.; Knutson, C.G.; Parry, N.M.; Muthupalani, S.; Ye, W.; Prestwich, E.; Cui, L.; McFaline, J.L.; Mobley, M.; Ge, Z.; et al. Infection-induced colitis in mice causes dynamic and tissue-specific changes in stress response and DNA damage leading to colon cancer. Proc. Natl. Acad. Sci. USA 2012, 109, E1820–E1829. [Google Scholar] [CrossRef]
- Haines, D.C.; Gorelick, P.L.; Battles, J.K.; Pike, K.M.; Anderson, R.J.; Fox, J.G.; Taylor, N.S.; Shen, Z.; Dewhirst, F.E.; Anver, M.R.; et al. Inflammatory large bowel disease in immunodeficient rats naturally and experimentally infected with helicobacter bilis. Vet. Pathol. 1998, 35, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Whary, M.T.; Fox, J.G. Detection, eradication, and research implications of helicobacter infections in laboratory rodents. Lab. Anim. 2006, 35, 25–27, 30–36. [Google Scholar] [CrossRef]
- Myles, M.H.; Livingston, R.S.; Franklin, C.L. Pathogenicity of helicobacter rodentium in a/jcr and scid mice. Comp. Med. 2004, 54, 549–557. [Google Scholar] [PubMed]
- Casey, K.M.; Johnson, A.L.; Hunrath, M.N.; Fraser, J.K.; McCowan, N.C.; Wasson, K.; Doty, R.A.; Griffey, S.M.; Imai, D.M. Proliferative typhlocolitis with multinucleated giant cells: A nonspecific enteropathy in immunodeficient sentinel mice. Vet. Pathol. 2019, 56, 157–168. [Google Scholar] [CrossRef]
- Zadrozny, L.M.; Brinster, L.R.; Rosenzweig, B.A.; Howard, K.E. Outbreak of opportunistic ascending pyelonephritis with numerous yeast after experimental humanization surgery in nod.Cg-prkdc(scid) il2rg(tm1wjl)/szj and nod.Cg-rag1(tm1mom) il2rg(tm1wjl)/szj immunodeficient mice. Comp. Med. 2018, 68, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Vedder, A.R.; Miedel, E.L.; Ragland, N.H.; Balasis, M.E.; Letson, C.T.; Engelman, R.W.; Padron, E. Effects of corynebacterium bovis on engraftment of patient-derived chronic-myelomonocytic leukemia cells in nsgs mice. Comp. Med. 2019, 69, 276–282. [Google Scholar] [CrossRef]
- Miedel, E.L.; Ragland, N.H.; Slate, A.R.; Engelman, R.W. Persistent corynebacterium bovis infectious hyperkeratotic dermatitis in immunocompetent epidermal-mutant dep/dep mice. Vet. Pathol. 2020, 57, 586–589. [Google Scholar] [CrossRef]
- Berard, M.; Medaille, C.; Simon, M.; Serre, S.; Pritchett-Corning, K.; Dangles-Marie, V. Ralstonia pickettii-induced ataxia in immunodeficient mice. Comp. Med. 2009, 59, 187–191. [Google Scholar] [PubMed]
- Dagnaes-Hansen, F.; Kilian, M.; Fuursted, K. Septicaemia associated with an aerococcus viridans infection in immunodeficient mice. Lab. Anim. 2004, 38, 321–325. [Google Scholar] [CrossRef]
- Shultz, L.D.; Schweitzer, P.A.; Hall, E.J.; Sundberg, J.P.; Taylor, S.; Walzer, P.D. Pneumocystis carinii pneumonia in scid/scid mice. Curr. Top. Microbiol. Immunol. 1989, 152, 243–249. [Google Scholar]
- Walzer, P.D.; Kim, C.K.; Linke, M.J.; Pogue, C.L.; Huerkamp, M.J.; Chrisp, C.E.; Lerro, A.V.; Wixson, S.K.; Hall, E.; Shultz, L.D. Outbreaks of pneumocystis carinii pneumonia in colonies of immunodeficient mice. Infect. Immun. 1989, 57, 62–70. [Google Scholar] [CrossRef]
- Henderson, K.S.; Dole, V.; Parker, N.J.; Momtsios, P.; Banu, L.; Brouillette, R.; Simon, M.A.; Albers, T.M.; Pritchett-Corning, K.R.; Clifford, C.B.; et al. Pneumocystis carinii causes a distinctive interstitial pneumonia in immunocompetent laboratory rats that had been attributed to “rat respiratory virus”. Vet. Pathol. 2012, 49, 440–452. [Google Scholar] [CrossRef]
- Dumoulin, A.; Mazars, E.; Seguy, N.; Gargallo-Viola, D.; Vargas, S.; Cailliez, J.C.; Aliouat, E.M.; Wakefield, A.E.; Dei-Cas, E. Transmission of pneumocystis carinii disease from immunocompetent contacts of infected hosts to susceptible hosts. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Macy, J.D., Jr.; Weir, E.C.; Compton, S.R.; Shlomchik, M.J.; Brownstein, D.G. Dual infection with pneumocystis carinii and pasteurella pneumotropica in b cell-deficient mice: Diagnosis and therapy. Comp. Med. 2000, 50, 49–55. [Google Scholar]
- Kawamoto, E.; Sasaki, H.; Okiyama, E.; Kanai, T.; Ueshiba, H.; Ohnishi, N.; Sawada, T.; Hayashimoto, N.; Takakura, A.; Itoh, T. Pathogenicity of pasteurella pneumotropica in immunodeficient nod/shijic-scid/jcl and immunocompetent crlj:Cd1 (icr) mice. Exp. Anim. 2011, 60, 463–470. [Google Scholar] [CrossRef][Green Version]
- Karst, S.M.; Wobus, C.E.; Lay, M.; Davidson, J.; Virgin, H.W.T. Stat1-dependent innate immunity to a norwalk-like virus. Science 2003, 299, 1575–1578. [Google Scholar] [CrossRef]
- Roediger, B.; Lee, Q.; Tikoo, S.; Cobbin, J.C.A.; Henderson, J.M.; Jormakka, M.; O’Rourke, M.B.; Padula, M.P.; Pinello, N.; Henry, M.; et al. An atypical parvovirus drives chronic tubulointerstitial nephropathy and kidney fibrosis. Cell 2018, 175, 530–543.e24. [Google Scholar] [CrossRef] [PubMed]
- Mähler, M.; Köhl, W. A serological survey to evaluate contemporary prevalence of viral agents and mycoplasma pulmonis in laboratory mice and rats in western europe. Lab. Anim. 2009, 38, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Pritchett-Corning, K.R.; Cosentino, J.; Clifford, C.B. Contemporary prevalence of infectious agents in laboratory mice and rats. Lab. Anim. 2009, 43, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Schoondermark-van de Ven, E.M.; Philipse-Bergmann, I.M.; van der Logt, J.T. Prevalence of naturally occurring viral infections, mycoplasma pulmonis and clostridium piliforme in laboratory rodents in western europe screened from 2000 to 2003. Lab. Anim. 2006, 40, 137–143. [Google Scholar] [CrossRef]
- Hayashimoto, N.; Morita, H.; Ishida, T.; Yasuda, M.; Kameda, S.; Uchida, R.; Tanaka, M.; Ozawa, M.; Sato, A.; Takakura, A.; et al. Current microbiological status of laboratory mice and rats in experimental facilities in japan. Exp. Anim. 2013, 62, 41–48. [Google Scholar] [CrossRef]
- Manjunath, S.; Kulkarni, P.G.; Nagavelu, K.; Samuel, R.J.; Srinivasan, S.; Ramasamy, N.; Hegde, N.R.; Gudde, R.S. Sero-prevalence of rodent pathogens in india. PLoS ONE 2015, 10, e0131706. [Google Scholar] [CrossRef]
- McInnes, E.F.; Rasmussen, L.; Fung, P.; Auld, A.M.; Alvarez, L.; Lawrence, D.A.; Quinn, M.E.; del Fierro, G.M.; Vassallo, B.A.; Stevenson, R. Prevalence of viral, bacterial and parasitological diseases in rats and mice used in research environments in australasia over a 5-y period. Lab. Anim. 2011, 40, 341–350. [Google Scholar] [CrossRef]
- Brownstein, D.G.; Smith, A.L.; Johnson, E.A.; Pintel, D.J.; Naeger, L.K.; Tattersall, P. The pathogenesis of infection with minute virus of mice depends on expression of the small nonstructural protein ns2 and on the genotype of the allotropic determinants vp1 and vp2. J. Virol. 1992, 66, 3118–3124. [Google Scholar] [CrossRef] [PubMed]
- Janus, L.M.; Bleich, A. Coping with parvovirus infections in mice: Health surveillance and control. Lab. Anim. 2012, 46, 14–23. [Google Scholar] [CrossRef]
- Kilham, L.; Margolis, G. Spontaneous hepatitis and cerebellar “hypoplasia” in suckling rats due to congenital infections with rat virus. Am. J. Pathol. 1966, 49, 457–475. [Google Scholar]
- Segovia, J.C.; Bueren, J.A.; Almendral, J.M. Myeloid depression follows infection of susceptible newborn mice with the parvovirus minute virus of mice (strain i). J. Virol. 1995, 69, 3229–3232. [Google Scholar] [CrossRef]
- Segovia, J.C.; Gallego, J.M.; Bueren, J.A.; Almendral, J.M. Severe leukopenia and dysregulated erythropoiesis in scid mice persistently infected with the parvovirus minute virus of mice. J. Virol. 1999, 73, 1774–1784. [Google Scholar] [CrossRef] [PubMed]
- Lamana, M.L.; Albella, B.; Bueren, J.A.; Segovia, J.C. In vitro and in vivo susceptibility of mouse megakaryocytic progenitors to strain i of parvovirus minute virus of mice. Exp. Hematol. 2001, 29, 1303–1309. [Google Scholar] [CrossRef]
- McKisic, M.D.; Paturzo, F.X.; Gaertner, D.J.; Jacoby, R.O.; Smith, A.L. A nonlethal rat parvovirus infection suppresses rat t lymphocyte effector functions. J. Immunol. 1995, 155, 3979–3986. [Google Scholar] [PubMed]
- Gripenberg-Lerche, C.; Toivanen, P. Variability in the induction of experimental arthritis: Yersinia associated arthritis in lewis rats. Scand. J. Rheumatol. 1994, 23, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Gabaldon, M.; Capdevila, C.; Zuniga, A. Effect of spontaneous pathology and thrombin on leukocyte adhesion to rat aortic endothelium. Atherosclerosis 1992, 93, 217–228. [Google Scholar] [CrossRef]
- McKisic, M.D.; Paturzo, F.X.; Smith, A.L. Mouse parvovirus infection potentiates rejection of tumor allografts and modulates t cell effector functions. Transplantation 1996, 61, 292–299. [Google Scholar] [CrossRef]
- Jacoby, R.O.; Johnson, E.A.; Ball-Goodrich, L.; Smith, A.L.; McKisic, M.D. Characterization of mouse parvovirus infection by in situ hybridization. J. Virol. 1995, 69, 3915–3919. [Google Scholar] [CrossRef]
- Janus, L.M.; Mahler, M.; Kohl, W.; Smoczek, A.; Hedrich, H.J.; Bleich, A. Minute virus of mice: Antibody response, viral shedding, and persistence of viral DNA in multiple strains of mice. Comp. Med. 2008, 58, 360–368. [Google Scholar] [PubMed]
- Janus, L.M.; Smoczek, A.; Hedrich, H.J.; Bleich, A. Risk assessment of minute virus of mice transmission during rederivation: Detection in reproductive organs, gametes, and embryos of mice after in vivo infection. Biol. Reprod. 2009, 81, 1010–1015. [Google Scholar] [CrossRef] [PubMed]
- Besselsen, D.G.; Becker, M.D.; Henderson, K.S.; Wagner, A.M.; Banu, L.A.; Shek, W.R. Temporal transmission studies of mouse parvovirus 1 in balb/c and c.B-17/icr-prkdc(scid) mice. Comp. Med. 2007, 57, 66–73. [Google Scholar] [PubMed]
- Besselsen, D.G.; Wagner, A.M.; Loganbill, J.K. Effect of mouse strain and age on detection of mouse parvovirus 1 by use of serologic testing and polymerase chain reaction analysis. Comp. Med. 2000, 50, 498–502. [Google Scholar]
- Henderson, K.S.; Pritchett-Corning, K.R.; Perkins, C.L.; Banu, L.A.; Jennings, S.M.; Francis, B.C.; Shek, W.R. A comparison of mouse parvovirus 1 infection in balb/c and c57bl/6 mice: Susceptibility, replication, shedding, and seroconversion. Comp. Med. 2015, 65, 5–14. [Google Scholar]
- Ward, J.M.; Wobus, C.E.; Thackray, L.B.; Erexson, C.R.; Faucette, L.J.; Belliot, G.; Barron, E.L.; Sosnovtsev, S.V.; Green, K.Y. Pathology of immunodeficient mice with naturally occurring murine norovirus infection. Toxicol. Pathol. 2006, 34, 708–715. [Google Scholar] [CrossRef] [PubMed]
- Cadwell, K.; Patel, K.K.; Maloney, N.S.; Liu, T.C.; Ng, A.C.; Storer, C.E.; Head, R.D.; Xavier, R.; Stappenbeck, T.S.; Virgin, H.W. Virus-plus-susceptibility gene interaction determines crohn’s disease gene atg16l1 phenotypes in intestine. Cell 2010, 141, 1135–1145. [Google Scholar] [CrossRef]
- Basic, M.; Keubler, L.M.; Buettner, M.; Achard, M.; Breves, G.; Schroder, B.; Smoczek, A.; Jorns, A.; Wedekind, D.; Zschemisch, N.H.; et al. Norovirus triggered microbiota-driven mucosal inflammation in interleukin 10-deficient mice. Inflamm. Bowel Dis. 2014, 20, 431–443. [Google Scholar] [CrossRef]
- Bolsega, S.; Basic, M.; Smoczek, A.; Buettner, M.; Eberl, C.; Ahrens, D.; Odum, K.A.; Stecher, B.; Bleich, A. Composition of the intestinal microbiota determines the outcome of virus-triggered colitis in mice. Front. Immunol. 2019, 10, 1708. [Google Scholar] [CrossRef] [PubMed]
- Patten, C.C., Jr.; Myles, M.H.; Franklin, C.L.; Livingston, R.S. Perturbations in cytokine gene expression after inoculation of c57bl/6 mice with pasteurella pneumotropica. Comp. Med. 2010, 60, 18–24. [Google Scholar] [PubMed]
- Garcia, A.; Zeng, Y.; Muthupalani, S.; Ge, Z.; Potter, A.; Mobley, M.W.; Boussahmain, C.; Feng, Y.; Wishnok, J.S.; Fox, J.G. Helicobacter hepaticus—Induced liver tumor promotion is associated with increased serum bile acid and a persistent microbial-Induced immune response. Cancer Res. 2011, 71, 2529–2540. [Google Scholar] [CrossRef]
- Hale, L.P.; Perera, D.; Gottfried, M.R.; Maggio-Price, L.; Srinivasan, S.; Marchuk, D. Neonatal co-infection with helicobacter species markedly accelerates the development of inflammation-associated colonic neoplasia in il-10(−/−) mice. Helicobacter 2007, 12, 598–604. [Google Scholar] [CrossRef]
- Kendall, L.V.; Riley, L.K.; Hook, R.R., Jr.; Besch-Williford, C.L.; Franklin, C.L. Antibody and cytokine responses to the cilium-associated respiratory bacillus in balb/c and c57bl/6 mice. Infect. Immun. 2000, 68, 4961–4967. [Google Scholar] [CrossRef][Green Version]
- Kendall, L.V.; Riley, L.K.; Hook, R.R., Jr.; Besch-Williford, C.L.; Franklin, C.L. Differential interleukin-10 and gamma interferon mrna expression in lungs of cilium-associated respiratory bacillus-infected mice. Infect. Immun. 2001, 69, 3697–3702. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Taurog, J.D.; Leary, S.L.; Cremer, M.A.; Mahowald, M.L.; Sandberg, G.P.; Manning, P.J. Infection with mycoplasma pulmonis modulates adjuvant- and collagen-induced arthritis in lewis rats. Arthritis Rheumatol. 1984, 27, 943–946. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Cuesta, J.; Vidal-Vanaclocha, F.; Mendoza, L.; Valcarcel, M.; Gallot, N.; Martinez de Tejada, G. Effect of asymptomatic natural infections due to common mouse pathogens on the metastatic progression of b16 murine melanoma in c57bl/6 mice. Clin. Exp. Metastasis 2005, 22, 549–558. [Google Scholar] [CrossRef]
- Hayashimoto, N.; Morita, H.; Ishida, T.; Uchida, R.; Tanaka, M.; Ozawa, M.; Yasuda, M.; Itoh, T. Microbiological survey of mice (mus musculus) purchased from commercial pet shops in kanagawa and tokyo, japan. Exp. Anim. 2015, 64, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Roble, G.S.; Gillespie, V.; Lipman, N.S. Infectious disease survey of mus musculus from pet stores in new york city. J. Am. Assoc. Lab. Anim. Sci. 2012, 51, 37–41. [Google Scholar]
- Hofmann, J.; Heuser, E.; Weiss, S.; Tenner, B.; Schoppmeyer, K.; Esser, J.; Klier, C.; Drewes, S.; Ulrich, R.G.; Kruger, D.H. Autochthonous ratborne seoul virus infection in woman with acute kidney injury. Emerg. Infect. Dis. 2020, 26, 3096–3099. [Google Scholar] [CrossRef] [PubMed]
- Bleich, A.; Nicklas, W. Zoonoses transmitted by mouse and rat maintained as laboratory or pet animals. Berl. Munch. Tierarztl. Wochenschr. 2008, 121, 241–255. [Google Scholar]
- Byers, K.B. Zoonotic infections from hantavirusand lymphocytic choriomeningitis virus(lcmv) associated with rodent coloniesthat were not experimentally infected. Appl. Biosaf. J. ABSA Int. 2018, 23, 143–152. [Google Scholar] [CrossRef]
- Dräger, S.; Marx, A.F.; Pigny, F.; Cherpillod, P.; Eisermann, P.; Sendi, P.; Widmer, A.F. Lymphocytic choriomeningitis virus meningitis after needlestick injury: A case report. Antimicrob. Resist. Infect. Control 2019, 8, 77. [Google Scholar] [CrossRef]
- Hayakawa, Y.; Suzuki, J.; Suzuki, M.; Sugiura, W.; Ohkusu, K. A case study of rat bite fever caused by streptobacillus moniliformis. Jpn. J. Infect. Dis. 2017, 70, 323–325. [Google Scholar] [CrossRef][Green Version]
- Hammer, A.; Wolff, D.; Geissdorfer, W.; Schrey, M.; Ziegler, R.; Steiner, H.H.; Bogdan, C. A spinal epidural abscess due to streptobacillus moniliformis infection following a rat bite: Case report. J. Neurosurg. Spine 2017, 27, 92–96. [Google Scholar] [CrossRef]
- Berset, C.M.; Caristo, M.E.; Ferrara, F.; Hardy, P.; Oropeza-Moe, M.; Waters, R. Federation of european laboratory animal science associations recommendations of best practices for the health management of ruminants and pigs used for scientific and educational purposes. Lab. Anim. 2020, 55, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Balansard, I.; Cleverley, L.; Cutler, K.L.; Spangberg, M.G.; Thibault-Duprey, K.; Langermans, J.A. Revised recommendations for health monitoring of non-human primate colonies (2018): Felasa working group report. Lab. Anim. 2019, 53, 429–446. [Google Scholar] [CrossRef]
- Hansen, A.K. Statistical aspects of health monitoring of laboratory animal colonies. Scand. J. Lab. Anim. Sci. 1993, 20, 11–14. [Google Scholar]
- Dubin, S.; Zietz, S. Sample size for animal health surveillance. Lab. Anim. 1991, 20, 29–33. [Google Scholar]
- ILAR. Committee on long-term holding of laboratoryrodents. Long-term holding of laboratory rodents. ILAR News 1976, 19, L1–L25. [Google Scholar]
- Besselsen, D.G.; Myers, E.L.; Franklin, C.L.; Korte, S.W.; Wagner, A.M.; Henderson, K.S.; Weigler, B.J. Transmission probabilities of mouse parvovirus 1 to sentinel mice chronically exposed to serial dilutions of contaminated bedding. Comp. Med. 2008, 58, 140–144. [Google Scholar] [PubMed]
- Brielmeier, M.; Mahabir, E.; Needham, J.R.; Lengger, C.; Wilhelm, P.; Schmidt, J. Microbiological monitoring of laboratory mice and biocontainment in individually ventilated cages: A field study. Lab. Anim. 2006, 40, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Compton, S.R.; Homberger, F.R.; Paturzo, F.X.; Clark, J.M. Efficacy of three microbiological monitoring methods in a ventilated cage rack. Comp. Med. 2004, 54, 382–392. [Google Scholar] [PubMed]
- Manuel, C.A.; Hsu, C.C.; Riley, L.K.; Livingston, R.S. Soiled-bedding sentinel detection of murine norovirus 4. J. Am. Assoc. Lab. Anim. Sci. 2008, 47, 31–36. [Google Scholar] [PubMed]
- Pullium, J.K.; Homberger, F.R.; Benjamin, K.A.; Dillehay, D.L.; Huerkamp, M.J. Confirmed persistent mouse hepatitis virus infection and transmission by mice with a targeted null mutation of tumor necrosis factor to sentinel mice, using short-term exposure. Comp. Med. 2003, 53, 439–443. [Google Scholar]
- Smith, P.C.; Nucifora, M.; Reuter, J.D.; Compton, S.R. Reliability of soiled bedding transfer for detection of mouse parvovirus and mouse hepatitis virus. Comp. Med. 2007, 57, 90–96. [Google Scholar]
- Whary, M.T.; Cline, J.H.; King, A.E.; Hewes, K.M.; Chojnacky, D.; Salvarrey, A.; Fox, J.G. Monitoring sentinel mice for helicobacter hepaticus, h rodentium, and h bilis infection by use of polymerase chain reaction analysis and serologic testing. Comp. Med. 2000, 50, 436–443. [Google Scholar]
- Buchheister, S.; Roegener, F.; Zschemisch, N.H.; Talbot, S.R.; Christensen, H.; Bleich, A. One for two: A novel and highly sensitive virulence factor-based quantitative polymerase chain reaction assay for the simultaneous detection of rodentibacter pneumotropicus and rodentibacter heylii in environmental sample material. Lab. Anim. 2019, 54, 239–250. [Google Scholar] [CrossRef]
- De Bruin, W.C.; van de Ven, E.M.; Hooijmans, C.R. Efficacy of soiled bedding transfer for transmission of mouse and rat infections to sentinels: A systematic review. PLoS ONE 2016, 11, e0158410. [Google Scholar] [CrossRef]
- Cundiff, D.D.; Riley, L.K.; Franklin, C.L.; Hook, R.R., Jr.; Besch-Williford, C. Failure of a soiled bedding sentinel system to detect cilia-associated respiratory bacillus infection in rats. Lab. Anim. Sci. 1995, 45, 219–221. [Google Scholar]
- Lindstrom, K.E.; Carbone, L.G.; Kellar, D.E.; Mayorga, M.S.; Wilkerson, J.D. Soiled bedding sentinels for the detection of fur mites in mice. J. Am. Assoc. Lab. Anim. Sci. 2011, 50, 54–60. [Google Scholar] [PubMed]
- Compton, S.R. Pcr and rt-pcr in the diagnosis of laboratory animal infections and in health monitoring. J. Am. Assoc. Lab. Anim. Sci. 2020, 59, 458–468. [Google Scholar] [CrossRef]
- Bauer, B.A.; Besch-Williford, C.; Livingston, R.S.; Crim, M.J.; Riley, L.K.; Myles, M.H. Influence of rack design and disease prevalence on detection of rodent pathogens in exhaust debris samples from individually ventilated caging systems. J. Am. Assoc. Lab. Anim. Sci. 2016, 55, 782–788. [Google Scholar]
- Mahabir, E.; Durand, S.; Henderson, K.S.; Hardy, P. Comparison of two prevalent individually ventilated caging systems for detection of murine infectious agents via exhaust air particles. Lab. Anim. 2019, 53, 84–88. [Google Scholar] [CrossRef]
- Niimi, K.; Maruyama, S.; Sako, N.; Miyata, K.; Yoshimoto, T.; Bileckei, B.; Henderson, K.S.; Takahashi, E. The sentinel (tm) ead(r) program can detect more microorganisms than bedding sentinel animals. Jpn. J. Vet. Res. 2018, 66, 125–129. [Google Scholar]
- Zorn, J.; Ritter, B.; Miller, M.; Kraus, M.; Northrup, E.; Brielmeier, M. Murine norovirus detection in the exhaust air of ivcs is more sensitive than serological analysis of soiled bedding sentinels. Lab. Anim. 2017, 51, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.; Ritter, B.; Zorn, J.; Brielmeier, M. Exhaust air dust monitoring is superior to soiled bedding sentinels for the detection of pasteurella pneumotropica in individually ventilated cage systems. J. Am. Assoc. Lab. Anim. Sci. 2016, 55, 775–781. [Google Scholar]
- Dubelko, A.R.; Zuwannin, M.; McIntee, S.C.; Livingston, R.S.; Foley, P.L. Pcr testing of filter material from ivc lids for microbial monitoring of mouse colonies. J. Am. Assoc. Lab. Anim. Sci. 2018, 57, 477–482. [Google Scholar] [CrossRef]
- Gerwin, P.M.; Ricart Arbona, R.J.; Riedel, E.R.; Henderson, K.S.; Lipman, N.S. Pcr testing of ivc filter tops as a method for detecting murine pinworms and fur mites. J. Am. Assoc. Lab. Anim. Sci. 2017, 56, 752–761. [Google Scholar] [PubMed]
- Miller, M.; Brielmeier, M. Environmental samples make soiled bedding sentinels dispensable for hygienic monitoring of ivc-reared mouse colonies. Lab. Anim. 2018, 52, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Pettan-Brewer, C.; Trost, R.J.; Maggio-Price, L.; Seamons, A.; Dowling, S.C. Adoption of exhaust air dust testing in spf rodent facilities. J. Am. Assoc. Lab. Anim. Sci. 2020, 59, 156–162. [Google Scholar] [CrossRef]
- Mailhiot, D.; Ostdiek, A.M.; Luchins, K.R.; Bowers, C.J.; Theriault, B.R.; Langan, G.P. Comparing mouse health monitoring between soiled-bedding sentinel and exhaust air dust surveillance programs. J. Am. Assoc. Lab. Anim. Sci. 2020, 59, 58–66. [Google Scholar] [CrossRef]
- Körner, C.; Miller, M.; Brielmeier, M. Detection of murine astrovirus and myocoptes musculinus in individually ventilated caging systems: Investigations to expose suitable detection methods for routine hygienic monitoring. PLoS ONE 2019, 14, e0221118. [Google Scholar] [CrossRef]
- Luchins, K.R.; Mailhiot, D.; Theriault, B.R.; Langan, G.P. Detection of lactate dehydrogenase elevating virus in a mouse vivarium using an exhaust air dust health monitoring program. J. Am. Assoc. Lab. Anim. Sci. 2020, 59, 328–333. [Google Scholar] [CrossRef]
- Jensen, E.S.; Allen, K.P.; Henderson, K.S.; Szabo, A.; Thulin, J.D. Pcr testing of a ventilated caging system to detect murine fur mites. J. Am. Assoc. Lab. Anim. Sci. 2013, 52, 28–33. [Google Scholar]
- Kapoor, P.; Hayes, Y.O.; Jarrell, L.T.; Bellinger, D.A.; Thomas, R.D.; Lawson, G.W.; Arkema, J.D.; Fletcher, C.A.; Nielsen, J.N. Evaluation of anthelmintic resistance and exhaust air dust pcr as a diagnostic tool in mice enzootically infected with aspiculuris tetraptera. J. Am. Assoc. Lab. Anim. Sci. 2017, 56, 273–289. [Google Scholar]
- Luchins, K.R.; Bowers, C.J.; Mailhiot, D.; Theriault, B.R.; Langan, G.P. Cost comparison of rodent soiled bedding sentinel and exhaust air dust health-monitoring programs. J. Am. Assoc. Lab. Anim. Sci. 2020, 59, 508–511. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.G.; Kim, N.; Ju, I.G.; Eo, H.; Lim, S.M.; Jang, S.E.; Kim, D.H.; Oh, M.S. Oral administration of proteus mirabilis damages dopaminergic neurons and motor functions in mice. Sci. Rep. 2018, 8, 1275. [Google Scholar] [CrossRef]
- Balish, E.; Warner, T. Enterococcus faecalis induces inflammatory bowel disease in interleukin-10 knockout mice. Am. J. Pathol. 2002, 160, 2253–2257. [Google Scholar] [CrossRef]
- Hansen, A.K.; Nielsen, D.S.; Krych, L.; Hansen, C.H.F. Bacterial species to be considered in quality assurance of mice and rats. Lab. Anim. 2019, 53, 281–291. [Google Scholar] [CrossRef]
- Shi, Y.; Mu, L. An expanding stage for commensal microbes in host immune regulation. Cell. Mol. Immunol. 2017, 14, 339–348. [Google Scholar] [CrossRef]
- Scher, J.U.; Sczesnak, A.; Longman, R.S.; Segata, N.; Ubeda, C.; Bielski, C.; Rostron, T.; Cerundolo, V.; Pamer, E.G.; Abramson, S.B.; et al. Expansion of intestinal prevotella copri correlates with enhanced susceptibility to arthritis. eLife 2013, 2, e01202. [Google Scholar] [CrossRef]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed]
- Hänninen, A.; Toivonen, R.; Poysti, S.; Belzer, C.; Plovier, H.; Ouwerkerk, J.P.; Emani, R.; Cani, P.D.; de Vos, W.M. Akkermansia muciniphila induces gut microbiota remodelling and controls islet autoimmunity in nod mice. Gut 2018, 67, 1445–1453. [Google Scholar] [CrossRef] [PubMed]
- Ganesh, B.P.; Klopfleisch, R.; Loh, G.; Blaut, M. Commensal akkermansia muciniphila exacerbates gut inflammation in salmonella typhimurium-infected gnotobiotic mice. PLoS ONE 2013, 8, e74963. [Google Scholar] [CrossRef]
- Kang, C.S.; Ban, M.; Choi, E.J.; Moon, H.G.; Jeon, J.S.; Kim, D.K.; Park, S.K.; Jeon, S.G.; Roh, T.Y.; Myung, S.J.; et al. Extracellular vesicles derived from gut microbiota, especially akkermansia muciniphila, protect the progression of dextran sulfate sodium-induced colitis. PLoS ONE 2013, 8, e76520. [Google Scholar] [CrossRef] [PubMed]
- Garland, C.D.; Lee, A.; Dickson, M.R. Segmented filamentous bacteria in the rodent small intestine: Their colonization of growing animals and possible role in host resistance tosalmonella. Microb. Ecol. 1982, 8, 181–190. [Google Scholar] [CrossRef]
- Stepankova, R.; Powrie, F.; Kofronova, O.; Kozakova, H.; Hudcovic, T.; Hrncir, T.; Uhlig, H.; Read, S.; Rehakova, Z.; Benada, O.; et al. Segmented filamentous bacteria in a defined bacterial cocktail induce intestinal inflammation in scid mice reconstituted with cd45rbhigh cd4+ t cells. Inflamm. Bowel Dis. 2007, 13, 1202–1211. [Google Scholar] [CrossRef]
- Ivanov, I.I.; Atarashi, K.; Manel, N.; Brodie, E.L.; Shima, T.; Karaoz, U.; Wei, D.; Goldfarb, K.C.; Santee, C.A.; Lynch, S.V. Induction of intestinal th17 cells by segmented filamentous bacteria. Cell 2009, 139, 485–498. [Google Scholar] [CrossRef]
- Wu, H.J.; Ivanov, I.I.; Darce, J.; Hattori, K.; Shima, T.; Umesaki, Y.; Littman, D.R.; Benoist, C.; Mathis, D. Gut-residing segmented filamentous bacteria drive autoimmune arthritis via t helper 17 cells. Immunity 2010, 32, 815–827. [Google Scholar] [CrossRef]
- Carlsson, A.H.; Yakymenko, O.; Olivier, I.; Hakansson, F.; Postma, E.; Keita, A.V.; Soderholm, J.D. Faecalibacterium prausnitzii supernatant improves intestinal barrier function in mice dss colitis. Scand. J. Gastroenterol. 2013, 48, 1136–1144. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Chain, F.; Miquel, S.; Lu, J.; Gratadoux, J.J.; Sokol, H.; Verdu, E.F.; Bercik, P.; Bermudez-Humaran, L.G.; Langella, P. The commensal bacterium faecalibacterium prausnitzii is protective in dnbs-induced chronic moderate and severe colitis models. Inflamm. Bowel Dis. 2014, 20, 417–430. [Google Scholar] [CrossRef]
- Rosique, R.M.; Bermudez-Humaran, L.G.; Chain, F.; Blennerhassett, P.; Gratadoux, J.J.; Sokol, H.; Verdu, E.F.; Bercik, P.; Langella, P. Protective and curative effect of faecalibacterium prausnitzii in a chronic dnbs-induced murine colitis. Gastroenterology 2012, 142, S392. [Google Scholar] [CrossRef]
- Zhang, M.; Qiu, X.; Zhang, H.; Yang, X.; Hong, N.; Yang, Y.; Chen, H.; Yu, C. Faecalibacterium prausnitzii inhibits interleukin-17 to ameliorate colorectal colitis in rats. PLoS ONE 2014, 9, e109146. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Round, J.L.; Kasper, D.L. A microbial symbiosis factor prevents intestinal inflammatory disease. Nature 2008, 453, 620–625. [Google Scholar] [CrossRef] [PubMed]
- Rath, H.C.; Wilson, K.H.; Sartor, R.B. Differential induction of colitis and gastritis in hla-b27 transgenic rats selectively colonized with bacteroides vulgatus or escherichia coli. Infect. Immun. 1999, 67, 2969–2974. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Rhee, K.J.; Albesiano, E.; Rabizadeh, S.; Wu, X.; Yen, H.R.; Huso, D.L.; Brancati, F.L.; Wick, E.; McAllister, F.; et al. A human colonic commensal promotes colon tumorigenesis via activation of t helper type 17 t cell responses. Nat. Med. 2009, 15, 1016–1022. [Google Scholar] [CrossRef]
- Basic, M.; Bleich, A. Gnotobiotics: Past, present and future. Lab. Anim. 2019, 53, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Simner, P.J.; Miller, S.; Carroll, K.C. Understanding the promises and hurdles of metagenomic next-generation sequencing as a diagnostic tool for infectious diseases. Clin. Infect. Dis. 2018, 66, 778–788. [Google Scholar] [CrossRef]
- Gu, W.; Miller, S.; Chiu, C.Y. Clinical metagenomic next-generation sequencing for pathogen detection. Annu. Rev. Pathol. 2019, 14, 319–338. [Google Scholar] [CrossRef] [PubMed]
- Keubler, L.M.; Buettner, M.; Hager, C.; Bleich, A. A multihit model: Colitis lessons from the interleukin-10-deficient mouse. Inflamm. Bowel Dis. 2015, 21, 1967–1975. [Google Scholar] [CrossRef] [PubMed]
- Mähler, M.; Leiter, E.H. Genetic and environmental context determines the course of colitis developing in il-10-deficient mice. Inflamm. Bowel Dis. 2002, 8, 347–355. [Google Scholar] [CrossRef]
- Hart, M.L.; Ericsson, A.C.; Franklin, C.L. Differing complex microbiota alter disease severity of the il-10(−/−) mouse model of inflammatory bowel disease. Front. Microbiol. 2017, 8, 792. [Google Scholar] [CrossRef]
- Bruesch, I.; Meier, P.; Vital, M.; Pieper, D.H.; Selke, K.; Bohlen, S.; Basic, M.; Meier, M.; Glage, S.; Hundrieser, J.; et al. Analysis of cdcs1 colitogenic effects in the hematopoietic compartment reveals distinct microbiome interaction and a new subcongenic interval active in t cells. Mucosal. Immunol. 2019, 12, 691–702. [Google Scholar] [CrossRef]
- Brinkman, B.M.; Becker, A.; Ayiseh, R.B.; Hildebrand, F.; Raes, J.; Huys, G.; Vandenabeele, P. Gut microbiota affects sensitivity to acute dss-induced colitis independently of host genotype. Inflamm. Bowel Dis. 2013, 19, 2560–2567. [Google Scholar] [CrossRef]
- Velazquez, E.M.; Nguyen, H.; Heasley, K.T.; Saechao, C.H.; Gil, L.M.; Rogers, A.W.L.; Miller, B.M.; Rolston, M.R.; Lopez, C.A.; Litvak, Y.; et al. Endogenous enterobacteriaceae underlie variation in susceptibility to salmonella infection. Nat. Microbiol. 2019, 4, 1057–1064. [Google Scholar] [CrossRef]
- Mullineaux-Sanders, C.; Collins, J.W.; Ruano-Gallego, D.; Levy, M.; Pevsner-Fischer, M.; Glegola-Madejska, I.T.; Sagfors, A.M.; Wong, J.L.C.; Elinav, E.; Crepin, V.F.; et al. Citrobacter rodentium relies on commensals for colonization of the colonic mucosa. Cell Rep. 2017, 21, 3381–3389. [Google Scholar] [CrossRef] [PubMed]
- Zackular, J.P.; Baxter, N.T.; Iverson, K.D.; Sadler, W.D.; Petrosino, J.F.; Chen, G.Y.; Schloss, P.D. The gut microbiome modulates colon tumorigenesis. mBio 2013, 4, e00692-13. [Google Scholar] [CrossRef]
- Ellekilde, M.; Krych, L.; Hansen, C.H.; Hufeldt, M.R.; Dahl, K.; Hansen, L.H.; Sorensen, S.J.; Vogensen, F.K.; Nielsen, D.S.; Hansen, A.K. Characterization of the gut microbiota in leptin deficient obese mice-correlation to inflammatory and diabetic parameters. Res. Vet. Sci. 2014, 96, 241–250. [Google Scholar] [CrossRef]
- Marietta, E.V.; Gomez, A.M.; Yeoman, C.; Tilahun, A.Y.; Clark, C.R.; Luckey, D.H.; Murray, J.A.; White, B.A.; Kudva, Y.C.; Rajagopalan, G. Low incidence of spontaneous type 1 diabetes in non-obese diabetic mice raised on gluten-free diets is associated with changes in the intestinal microbiome. PLoS ONE 2013, 8, e78687. [Google Scholar] [CrossRef]
- Ussar, S.; Griffin, N.W.; Bezy, O.; Fujisaka, S.; Vienberg, S.; Softic, S.; Deng, L.; Bry, L.; Gordon, J.I.; Kahn, C.R. Interactions between gut microbiota, host genetics and diet modulate the predisposition to obesity and metabolic syndrome. Cell Metab. 2015, 22, 516–530. [Google Scholar] [CrossRef] [PubMed]
- Brenner, D.A.; Paik, Y.H.; Schnabl, B. Role of gut microbiota in liver disease. J. Clin. Gastroenterol. 2015, 49 (Suppl. 1), S25–S27. [Google Scholar] [CrossRef]
- Hrncir, T.; Hrncirova, L.; Kverka, M.; Tlaskalova-Hogenova, H. The role of gut microbiota in intestinal and liver diseases. Lab. Anim. 2019, 53, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of lactobacillus strain regulates emotional behavior and central gaba receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef]
- Bangsgaard Bendtsen, K.M.; Krych, L.; Sorensen, D.B.; Pang, W.; Nielsen, D.S.; Josefsen, K.; Hansen, L.H.; Sorensen, S.J.; Hansen, A.K. Gut microbiota composition is correlated to grid floor induced stress and behavior in the balb/c mouse. PLoS ONE 2012, 7, e46231. [Google Scholar]
- Bercik, P.; Denou, E.; Collins, J.; Jackson, W.; Lu, J.; Jury, J.; Deng, Y.; Blennerhassett, P.; Macri, J.; McCoy, K.D.; et al. The intestinal microbiota affect central levels of brain-derived neurotropic factor and behavior in mice. Gastroenterology 2011, 141, 599–609.e3. [Google Scholar] [CrossRef]
- Hsiao, E.Y.; McBride, S.W.; Hsien, S.; Sharon, G.; Hyde, E.R.; McCue, T.; Codelli, J.A.; Chow, J.; Reisman, S.E.; Petrosino, J.F.; et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 2013, 155, 1451–1463. [Google Scholar] [CrossRef]
- Sharon, G.; Cruz, N.J.; Kang, D.W.; Gandal, M.J.; Wang, B.; Kim, Y.M.; Zink, E.M.; Casey, C.P.; Taylor, B.C.; Lane, C.J.; et al. Human gut microbiota from autism spectrum disorder promote behavioral symptoms in mice. Cell 2019, 177, 1600–1618.e17. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Menezes, J.S.; Umesaki, Y.; Mazmanian, S.K. Proinflammatory t-cell responses to gut microbiota promote experimental autoimmune encephalomyelitis. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4615–4622. [Google Scholar] [CrossRef]
- Franklin, C.L.; Ericsson, A.C. Microbiota and reproducibility of rodent models. Lab. Anim. 2017, 46, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Baker, M. Is there a reproducibility crisis? Nature 2016, 533, 452–454. [Google Scholar] [CrossRef] [PubMed]
- Kafkafi, N.; Agassi, J.; Chesler, E.J.; Crabbe, J.C.; Crusio, W.E.; Eilam, D.; Gerlai, R.; Golani, I.; Gomez-Marin, A.; Heller, R.; et al. Reproducibility and replicability of rodent phenotyping in preclinical studies. Neurosci. Biobehav. Rev. 2018, 87, 218–232. [Google Scholar] [CrossRef]
- Dirnagl, U. Rethinking research reproducibility. EMBO J. 2019, 38, e101117. [Google Scholar] [CrossRef]
- Würbel, H. Behaviour and the standardization fallacy. Nat. Genet. 2000, 26, 263. [Google Scholar] [CrossRef]
- Witjes, V.M.; Boleij, A.; Halffman, W. Reducing versus embracing variation as strategies for reproducibility: The microbiome of laboratory mice. Animals 2020, 10, 2415. [Google Scholar] [CrossRef]
- Stappenbeck, T.S.; Virgin, H.W. Accounting for reciprocal host-microbiome interactions in experimental science. Nature 2016, 534, 191–199. [Google Scholar] [CrossRef]
- Hildebrand, F.; Nguyen, T.L.; Brinkman, B.; Yunta, R.G.; Cauwe, B.; Vandenabeele, P.; Liston, A.; Raes, J. Inflammation-associated enterotypes, host genotype, cage and inter-individual effects drive gut microbiota variation in common laboratory mice. Genome Biol. 2013, 14, R4. [Google Scholar] [CrossRef] [PubMed]
- Leamy, L.J.; Kelly, S.A.; Nietfeldt, J.; Legge, R.M.; Ma, F.; Hua, K.; Sinha, R.; Peterson, D.A.; Walter, J.; Benson, A.K.; et al. Host genetics and diet, but not immunoglobulin a expression, converge to shape compositional features of the gut microbiome in an advanced intercross population of mice. Genome Biol. 2014, 15, 552. [Google Scholar] [CrossRef]
- Caruso, R.; Ono, M.; Bunker, M.E.; Nunez, G.; Inohara, N. Dynamic and asymmetric changes of the microbial communities after cohousing in laboratory mice. Cell Rep. 2019, 27, 3401–3412.e3. [Google Scholar] [CrossRef] [PubMed]
- Hansen, C.H.; Metzdorff, S.B.; Hansen, A.K. Customizing laboratory mice by modifying gut microbiota and host immunity in an early “window of opportunity”. Gut Microbes 2013, 4, 241–245. [Google Scholar] [CrossRef]
- Cahenzli, J.; Koller, Y.; Wyss, M.; Geuking, M.B.; McCoy, K.D. Intestinal microbial diversity during early-life colonization shapes long-term ige levels. Cell Host Microbe 2013, 14, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Torow, N.; Hornef, M.W. The neonatal window of opportunity: Setting the stage for life-long host-microbial interaction and immune homeostasis. J. Immunol. 2017, 198, 557–563. [Google Scholar] [CrossRef]
- Robertson, S.J.; Lemire, P.; Maughan, H.; Goethel, A.; Turpin, W.; Bedrani, L.; Guttman, D.S.; Croitoru, K.; Girardin, S.E.; Philpott, D.J. Comparison of co-housing and littermate methods for microbiota standardization in mouse models. Cell Rep. 2019, 27, 1910–1919. [Google Scholar] [CrossRef]
- Macpherson, A.J.; McCoy, K.D. Standardised animal models of host microbial mutualism. Mucosal. Immunol. 2015, 8, 476–486. [Google Scholar] [CrossRef]
- Schaedler, R.W.; Dubs, R.; Costello, R. Association of germfree mice with bacteria isolated from normal mice. J. Exp. Med. 1965, 122, 77–82. [Google Scholar] [CrossRef]
- Wymore Brand, M.; Wannemuehler, M.J.; Phillips, G.J.; Proctor, A.; Overstreet, A.M.; Jergens, A.E.; Orcutt, R.P.; Fox, J.G. The altered schaedler flora: Continued applications of a defined murine microbial community. ILAR J. 2015, 56, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Brugiroux, S.; Beutler, M.; Pfann, C.; Garzetti, D.; Ruscheweyh, H.J.; Ring, D.; Diehl, M.; Herp, S.; Lotscher, Y.; Hussain, S.; et al. Genome-guided design of a defined mouse microbiota that confers colonization resistance against salmonella enterica serovar typhimurium. Nat. Microbiol. 2016, 2, 16215. [Google Scholar] [CrossRef] [PubMed]
- Becker, N.; Kunath, J.; Loh, G.; Blaut, M. Human intestinal microbiota: Characterization of a simplified and stable gnotobiotic rat model. Gut Microbes 2011, 2, 25–33. [Google Scholar] [CrossRef]
- Uchimura, Y.; Wyss, M.; Brugiroux, S.; Limenitakis, J.P.; Stecher, B.; McCoy, K.D.; Macpherson, A.J. Complete genome sequences of 12 species of stable defined moderately diverse mouse microbiota 2. Genome Announc. 2016, 4, e00951-16. [Google Scholar] [CrossRef] [PubMed]
- Norin, E.; Midtvedt, T. Intestinal microflora functions in laboratory mice claimed to harbor a “normal” intestinal microflora. Is the spf concept running out of date? Anaerobe 2010, 16, 311–313. [Google Scholar] [CrossRef]
- Lagkouvardos, I.; Pukall, R.; Abt, B.; Foesel, B.U.; Meier-Kolthoff, J.P.; Kumar, N.; Bresciani, A.; Martinez, I.; Just, S.; Ziegler, C.; et al. The mouse intestinal bacterial collection (mibc) provides host-specific insight into cultured diversity and functional potential of the gut microbiota. Nat. Microbiol. 2016, 1, 16131, Erratum in 2016, 1, 16219. [Google Scholar] [CrossRef]
- Herp, S.; Brugiroux, S.; Garzetti, D.; Ring, D.; Jochum, L.M.; Beutler, M.; Eberl, C.; Hussain, S.; Walter, S.; Gerlach, R.G.; et al. Mucispirillum schaedleri antagonizes salmonella virulence to protect mice against colitis. Cell Host Microbe 2019, 25, 681–694.e8. [Google Scholar] [CrossRef] [PubMed]
- Streidl, T.; Karkossa, I.; Segura Munoz, R.R.; Eberl, C.; Zaufel, A.; Plagge, J.; Schmaltz, R.; Schubert, K.; Basic, M.; Schneider, K.M.; et al. The gut bacterium extibacter muris produces secondary bile acids and influences liver physiology in gnotobiotic mice. Gut Microbes 2021, 13, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Wyss, M.; Brown, K.; Thomson, C.A.; Koegler, M.; Terra, F.; Fan, V.; Ronchi, F.; Bihan, D.; Lewis, I.; Geuking, M.B.; et al. Using precisely defined in vivo microbiotas to understand microbial regulation of ige. Front. Immunol. 2019, 10, 3107. [Google Scholar] [CrossRef]
- Studer, N.; Desharnais, L.; Beutler, M.; Brugiroux, S.; Terrazos, M.A.; Menin, L.; Schurch, C.M.; McCoy, K.D.; Kuehne, S.A.; Minton, N.P.; et al. Functional intestinal bile acid 7alpha-dehydroxylation by clostridium scindens associated with protection from clostridium difficile infection in a gnotobiotic mouse model. Front. Cell Infect. Microbiol. 2016, 6, 191. [Google Scholar] [CrossRef]
- Beura, L.K.; Hamilton, S.E.; Bi, K.; Schenkel, J.M.; Odumade, O.A.; Casey, K.A.; Thompson, E.A.; Fraser, K.A.; Rosato, P.C.; Filali-Mouhim, A.; et al. Normalizing the environment recapitulates adult human immune traits in laboratory mice. Nature 2016, 532, 512–516. [Google Scholar] [CrossRef]
- Abolins, S.; King, E.C.; Lazarou, L.; Weldon, L.; Hughes, L.; Drescher, P.; Raynes, J.G.; Hafalla, J.C.R.; Viney, M.E.; Riley, E.M. The comparative immunology of wild and laboratory mice, mus musculus domesticus. Nat. Commun. 2017, 8, 14811. [Google Scholar] [CrossRef]
- Abolins, S.; Lazarou, L.; Weldon, L.; Hughes, L.; King, E.C.; Drescher, P.; Pocock, M.J.O.; Hafalla, J.C.R.; Riley, E.M.; Viney, M. The ecology of immune state in a wild mammal, mus musculus domesticus. PLoS Biol. 2018, 16, e2003538. [Google Scholar] [CrossRef] [PubMed]
- Beans, C. News feature: What happens when lab animals go wild. Proc. Natl. Acad. Sci. USA 2018, 115, 3196–3199. [Google Scholar] [CrossRef]
- Flies, A.S.; Woods, G.M. Editorial: Wild immunology-the answers are out there. Front. Immunol. 2019, 10, 126. [Google Scholar] [CrossRef]
- Weldon, L.; Abolins, S.; Lenzi, L.; Bourne, C.; Riley, E.M.; Viney, M. The gut microbiota of wild mice. PLoS ONE 2015, 10, e0134643. [Google Scholar] [CrossRef]
- Maurice, C.F.; Knowles, S.C.; Ladau, J.; Pollard, K.S.; Fenton, A.; Pedersen, A.B.; Turnbaugh, P.J. Marked seasonal variation in the wild mouse gut microbiota. ISME J. 2015, 9, 2423–2434. [Google Scholar] [CrossRef]
- Kohl, K.D.; Dearing, M.D. Wild-caught rodents retain a majority of their natural gut microbiota upon entrance into captivity. Environ. Microbiol. Rep. 2014, 6, 191–195. [Google Scholar] [CrossRef]
- Kreisinger, J.; Bastien, G.; Hauffe, H.C.; Marchesi, J.; Perkins, S.E. Interactions between multiple helminths and the gut microbiota in wild rodents. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140295. [Google Scholar] [CrossRef]
- Kreisinger, J.; Cizkova, D.; Vohanka, J.; Pialek, J. Gastrointestinal microbiota of wild and inbred individuals of two house mouse subspecies assessed using high-throughput parallel pyrosequencing. Mol. Ecol. 2014, 23, 5048–5060. [Google Scholar] [CrossRef]
- Rosshart, S.P.; Vassallo, B.G.; Angeletti, D.; Hutchinson, D.S.; Morgan, A.P.; Takeda, K.; Hickman, H.D.; McCulloch, J.A.; Badger, J.H.; Ajami, N.J.; et al. Wild mouse gut microbiota promotes host fitness and improves disease resistance. Cell 2017, 171, 1015–1028.e13. [Google Scholar] [CrossRef]
- Rosshart, S.P.; Herz, J.; Vassallo, B.G.; Hunter, A.; Wall, M.K.; Badger, J.H.; McCulloch, J.A.; Anastasakis, D.G.; Sarshad, A.A.; Leonardi, I.; et al. Laboratory mice born to wild mice have natural microbiota and model human immune responses. Science 2019, 365, eaaw4361. [Google Scholar] [CrossRef]
- Graham, A.L. Naturalizing mouse models for immunology. Nat. Immunol. 2021, 22, 111–117. [Google Scholar] [CrossRef]
- Viney, M.; Riley, E.M. The immunology of wild rodents: Current status and future prospects. Front. Immunol. 2017, 8, 1481. [Google Scholar] [CrossRef]
- Viney, M. The gut microbiota of wild rodents: Challenges and opportunities. Lab. Anim. 2019, 53, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Jirkof, P.; Abdelrahman, A.; Bleich, A.; Durst, M.; Keubler, L.; Potschka, H.; Struve, B.; Talbot, S.R.; Vollmar, B.; Zechner, D.; et al. A safe bet? Inter-laboratory variability in behaviour-based severity assessment. Lab. Anim. 2020, 54, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.; Naccache, S.N.; Samayoa, E.; Messacar, K.; Arevalo, S.; Federman, S.; Stryke, D.; Pham, E.; Fung, B.; Bolosky, W.J.; et al. Laboratory validation of a clinical metagenomic sequencing assay for pathogen detection in cerebrospinal fluid. Genome Res. 2019, 29, 831–842. [Google Scholar] [CrossRef]
- Thoendel, M.J.; Jeraldo, P.R.; Greenwood-Quaintance, K.E.; Yao, J.Z.; Chia, N.; Hanssen, A.D.; Abdel, M.P.; Patel, R. Identification of prosthetic joint infection pathogens using a shotgun metagenomics approach. Clin. Infect. Dis. 2018, 67, 1333–1338. [Google Scholar] [CrossRef]
- Munk, P.; Andersen, V.D.; de Knegt, L.; Jensen, M.S.; Knudsen, B.E.; Lukjancenko, O.; Mordhorst, H.; Clasen, J.; Agerso, Y.; Folkesson, A.; et al. A sampling and metagenomic sequencing-based methodology for monitoring antimicrobial resistance in swine herds. J. Antimicrob. Chemother. 2017, 72, 385–392. [Google Scholar] [CrossRef]
- Schlaberg, R.; Chiu, C.Y.; Miller, S.; Procop, G.W.; Weinstock, G.; The Professional Practice Committee and Committee on Laboratory Practices of the American Society for Microbiology; The Microbiology Resource Committee of the College of American Pathologists. Validation of metagenomic next-generation sequencing tests for universal pathogen detection. Arch. Pathol. Lab. Med. 2017, 141, 776–786. [Google Scholar] [CrossRef]
- Osborne, N.; Avey, M.T.; Anestidou, L.; Ritskes-Hoitinga, M.; Griffin, G. Improving animal research reporting standards: Harrp, the first step of a unified approach by iclas to improve animal research reporting standards worldwide. EMBO Rep. 2018, 19, e46069. [Google Scholar] [CrossRef]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The arrive guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar]
- Smith, A.J.; Clutton, R.E.; Lilley, E.; Hansen, K.E.A.; Brattelid, T. Prepare: Guidelines for planning animal research and testing. Lab. Anim. 2018, 52, 135–141. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buchheister, S.; Bleich, A. Health Monitoring of Laboratory Rodent Colonies—Talking about (R)evolution. Animals 2021, 11, 1410. https://doi.org/10.3390/ani11051410
Buchheister S, Bleich A. Health Monitoring of Laboratory Rodent Colonies—Talking about (R)evolution. Animals. 2021; 11(5):1410. https://doi.org/10.3390/ani11051410
Chicago/Turabian StyleBuchheister, Stephanie, and André Bleich. 2021. "Health Monitoring of Laboratory Rodent Colonies—Talking about (R)evolution" Animals 11, no. 5: 1410. https://doi.org/10.3390/ani11051410
APA StyleBuchheister, S., & Bleich, A. (2021). Health Monitoring of Laboratory Rodent Colonies—Talking about (R)evolution. Animals, 11(5), 1410. https://doi.org/10.3390/ani11051410