
Effects of Starch Level and a Mixture of Sunflower and Fish Oils on Nutrient Intake and Digestibility, Rumen Fermentation, and Ruminal Methane Emissions in Dairy Cows

 , , ,
, , , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Experimental Design and Diets
2.2. Feed Intake, Milk Yield and Chemical Analysis
2.3. Rumen Fermentation
2.4. Apparent Total-Tract Digestibility
2.5. Ruminal Gas Production
2.6. Microbial Analysis
2.7. Calculations
2.8. Statistical Analysis
3. Results
3.1. Dry Matter and Nutrient Intake and Milk Yield
3.2. Apparent Total-Tract Digestibility
3.3. Rumen Fermentation
3.4. Ruminal CH4 and CO2 Emission
3.5. Rumen Microbial Ecology
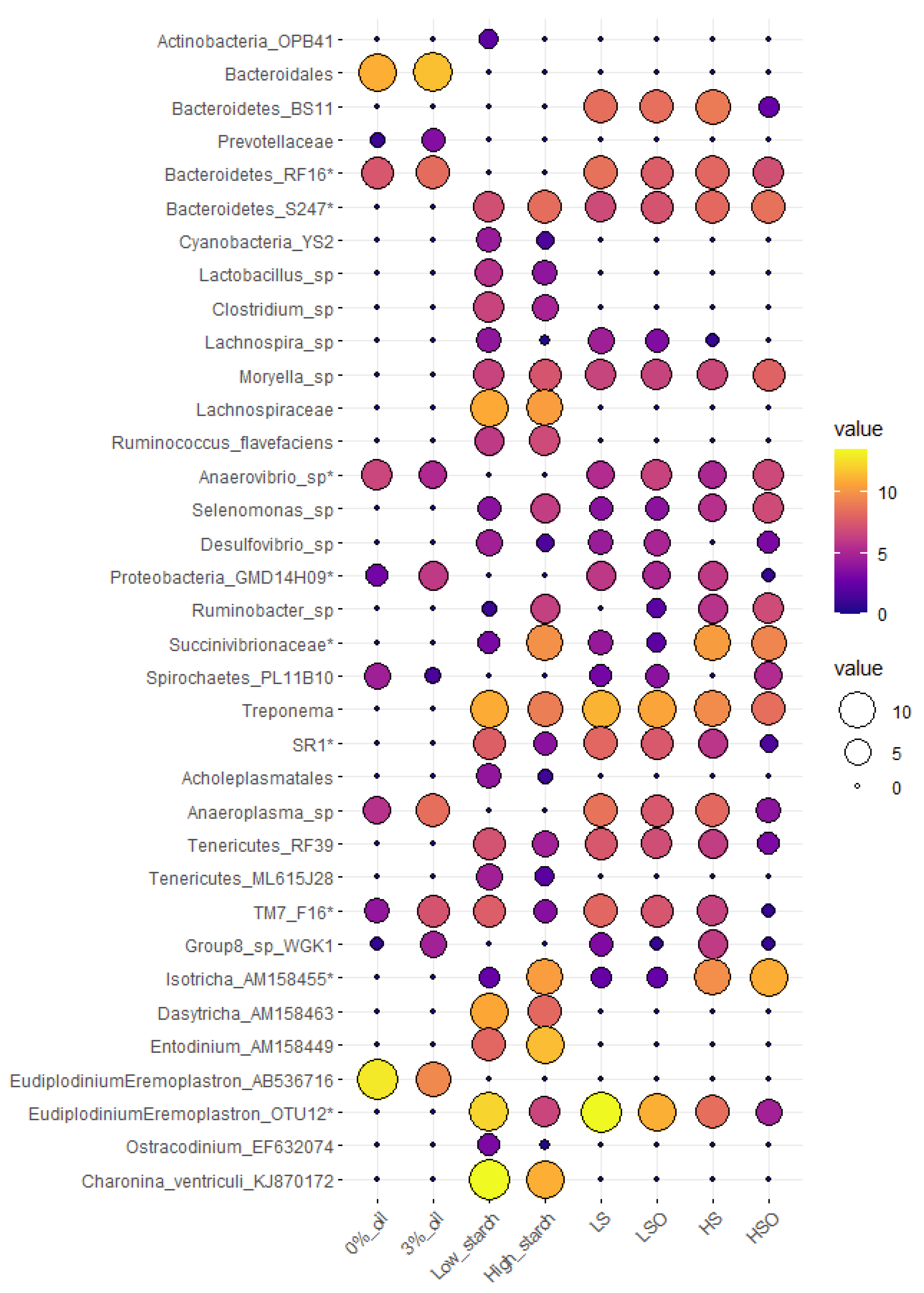
3.6. Taxa Affected by Diet, Oil and Starch
3.7. Microbiota Association with Rumen Fermentation and Methane Production Traits
4. Discussion
4.1. Dry Matter Intake, Milk Yield, and Nutrient Digestibility
4.2. Methane Production and Rumen Fermentation
4.3. Rumen Microbial Ecology
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hristov, A.N.; Oh, J.; Firkins, J.; Dijkstra, J.; Kebreab, E.; Waghorn, G.; Makkar, H.; Adesogan, A.; Yang, W.; Lee, C. Special topics—Mitigation of methane and nitrous oxide emissions from animal operations: I. A review of enteric methane mitigation options. J. Anim. Sci. 2013, 91, 5045–5069. [Google Scholar] [CrossRef]
- Grainger, C.; Beauchemin, K. Can enteric methane emissions from ruminants be lowered without lowering their production? Anim. Feed Sci. Technol. 2011, 166, 308–320. [Google Scholar] [CrossRef]
- Knapp, J.R.; Laur, G.; Vadas, P.A.; Weiss, W.P.; Tricarico, J.M. Invited review: Enteric methane in dairy cattle production: Quantifying the opportunities and impact of reducing emissions. J. Dairy Sci. 2014, 97, 3231–3261. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Morgavi, D.; Doreau, M. Methane mitigation in ruminants: From microbe to the farm scale. Animal 2010, 4, 351–365. [Google Scholar] [CrossRef] [PubMed]
- Shibata, M.; Terada, F. Factors affecting methane production and mitigation in ruminants. Anim. Sci. J. 2010, 81, 2–10. [Google Scholar] [CrossRef]
- Hatew, B.; Cone, J.; Pellikaan, W.; Podesta, S.; Bannink, A.; Hendriks, W.; Dijkstra, J. Relationship between in vitro and in vivo methane production measured simultaneously with different dietary starch sources and starch levels in dairy cattle. Anim. Feed Sci. Technol. 2015, 202, 20–31. [Google Scholar] [CrossRef]
- Bayat, A.R.; Tapio, I.; Vilkki, J.; Shingfield, K.; Leskinen, H. Plant oil supplements reduce methane emissions and improve milk fatty acid composition in dairy cows fed grass silage-based diets without affecting milk yield. J. Dairy Sci. 2018, 101, 1136–1151. [Google Scholar] [CrossRef]
- Lanier, J.S.; Corl, B.A. Challenges in enriching milk fat with polyunsaturated fatty acids. J. Anim. Sci. Biotechnol. 2015, 6, 26. [Google Scholar] [CrossRef]
- European Union. Council Directive 86/609/EEC on the Approximation of Laws, Regulations and Administrative Provisions of the Member States regarding the Protection of Animals used for Experimental and other Scientific Purposes. Off. J. 1986, L358, 1–28. [Google Scholar]
- Kairenius, P. Role of Dietary Fish Oil and Plant Oil Supplements in Ruminal Lipid Metabolism and Fish Oil-Induced Milk Fat Depression in Lactating Cows. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2020. [Google Scholar]
- MTT Agrifood Research Finland. Finnish Feed Tables and Feeding Recommendations. 2006. Available online: http://www.luke.fi/rehutaulukot (accessed on 13 August 2011).
- Shingfield, K.; Jaakkola, S.; Huhtanen, P. Effects of level of nitrogen fertilizer application and various nitrogenous supplements on milk production and nitrogen utilization of dairy cows given grass silage-based diets. Anim. Sci. 2001, 73, 541–554. [Google Scholar] [CrossRef]
- Huida, L.; Väätäinen, H.; Lampila, M. Comparison of dry matter contents in grass silages as determined by oven drying and gas chromatographic water analysis. Ann. Agric. Fenn. 1986, 25, 215–230. [Google Scholar]
- Bayat, A.R.; Kairenius, P.; Stefański, T.; Leskinen, H.; Comtet-Marre, S.; Forano, E.; Chaucheyras-Durand, F.; Shingfield, K. Effect of camelina oil or live yeasts (Saccharomyces cerevisiae) on ruminal methane production, rumen fermentation, and milk fatty acid composition in lactating cows fed grass silage diets. J. Dairy Sci. 2015, 98, 3166–3181. [Google Scholar] [CrossRef] [PubMed]
- Bayat, A.R.; Stefański, T.; Luukkonen, T.; Kairenius, P.; Leskinen, H.; Vilkki, J. Sulphur hexafluoride tracer technique for measuring methane directly from rumen of dairy cows validated with respiration chambers. In Proceedings of the International Symposium on Emission of Gas and Dust from Livestock, Saint-Malo, France, 21–24 May 2017; pp. 98–99. [Google Scholar]
- Regina, K.; Alakukku, L. Greenhouse gas fluxes in varying soils types under conventional and no-tillage practices. Soil Tillage Res. 2010, 109, 144–152. [Google Scholar] [CrossRef]
- Yu, Z.; Morrison, M. Improved extraction of PCR-quality community DNA from digesta and fecal samples. Biotechniques 2004, 36, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Tapio, I.; Shingfield, K.J.; McKain, N.; Bonin, A.; Fischer, D.; Bayat, A.R.; Vilkki, J.; Taberlet, P.; Snelling, T.J.; Wallace, R.J. Oral samples as non-invasive proxies for assessing the composition of the rumen microbial community. PLoS ONE 2016, 11, e0151220. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef]
- Seedorf, H.; Kittelmann, S.; Henderson, G.; Janssen, P.H. RIM-DB: A taxonomic framework for community structure analysis of methanogenic archaea from the rumen and other intestinal environments. PeerJ 2014, 2, e494. [Google Scholar] [CrossRef]
- Kittelmann, S.; Devente, S.R.; Kirk, M.R.; Seedorf, H.; Dehority, B.A.; Janssen, P.H. Phylogeny of intestinal ciliates, including Charonina ventriculi, and comparison of microscopy and 18S rRNA gene pyrosequencing for rumen ciliate community structure analysis. Appl. Environ. Microbiol. 2015, 81, 2433–2444. [Google Scholar] [CrossRef]
- Kriss, M. Quantitative relations of the dry matter of the food consumed, the heat production, the gaseous outgo, and the insensible loss in body weight of cattle. J. Agric. Res. 1930, 40, 283–295. [Google Scholar]
- Sjaunja, L. A Nordic proposal for an energy-corrected milk (ECM) formula. In Proceedings of the 27th Session International Committee for Recording and Productivity of Milk Animals, Paris, France, 2–6 July 1990. [Google Scholar]
- Zakrzewski, M.; Proietti, C.; Ellis, J.J.; Hasan, S.; Brion, M.-J.; Berger, B.; Krause, L. Calypso: A user-friendly web-server for mining and visualizing microbiome–environment interactions. Bioinformatics 2017, 33, 782–783. [Google Scholar] [CrossRef] [PubMed]
- Bayat, A.R.; Ventto, L.; Kairenius, P.; Stefański, T.; Leskinen, H.; Tapio, I.; Negussie, E.; Vilkki, J.; Shingfield, K. Dietary forage to concentrate ratio and sunflower oil supplement alter rumen fermentation, ruminal methane emissions, and nutrient utilization in lactating cows. Transl. Anim. Sci. 2017, 1, 277–286. [Google Scholar] [CrossRef]
- Benchaar, C.; Hassanat, F.; Martineau, R.; Gervais, R. Linseed oil supplementation to dairy cows fed diets based on red clover silage or corn silage: Effects on methane production, rumen fermentation, nutrient digestibility, N balance, and milk production. J. Dairy Sci. 2015, 98, 7993–8008. [Google Scholar] [CrossRef] [PubMed]
- Fievez, V.; Dohme, F.; Danneels, M.; Raes, K.; Demeyer, D. Fish oils as potent rumen methane inhibitors and associated effects on rumen fermentation in vitro and in vivo. Anim. Feed Sci. Technol. 2003, 104, 41–58. [Google Scholar] [CrossRef]
- Vafa, T.S.; Naserian, A.A.; Moussavi, A.R.H.; Valizadeh, R.; Mesgaran, M.D. Effect of supplementation of fish and canola oil in the diet on milk fatty acid composition in early lactating holstein cows. Asian Australas. J. Anim. Sci. 2012, 25, 311. [Google Scholar] [CrossRef] [PubMed]
- Kairenius, P.; Leskinen, H.; Toivonen, V.; Muetzel, S.; Ahvenjärvi, S.; Vanhatalo, A.; Huhtanen, P.; Wallace, R.; Shingfield, K.J. Effect of dietary fish oil supplements alone or in combination with sunflower and linseed oil on ruminal lipid metabolism and bacterial populations in lactating cows. J. Dairy Sci. 2018, 101, 3021–3035. [Google Scholar] [CrossRef]
- Shingfield, K.J.; Reynolds, C.K.; Hervás, G.; Griinari, J.M.; Grandison, A.S.; Beever, D.E. Examination of the persistency of milk fatty acid composition responses to fish oil and sunflower oil in the diet of dairy cows. J. Dairy Sci. 2006, 89, 714–732. [Google Scholar] [CrossRef]
- Thanh, L.P.; Suksombat, W. Milk yield, composition, and fatty acid profile in dairy cows fed a high-concentrate diet blended with oil mixtures rich in polyunsaturated fatty acids. Asian Australas. J. Anim. Sci. 2015, 28, 796. [Google Scholar] [CrossRef]
- Rabiee, A.; Breinhild, K.; Scott, W.; Golder, H.; Block, E.; Lean, I. Effect of fat additions to diets of dairy cattle on milk production and components: A meta-analysis and meta-regression. J. Dairy Sci. 2012, 95, 3225–3247. [Google Scholar] [CrossRef] [PubMed]
- Weld, K.; Armentano, L. The effects of adding fat to diets of lactating dairy cows on total-tract neutral detergent fiber digestibility: A meta-analysis. J. Dairy Sci. 2017, 100, 1766–1779. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Ferlay, A.; Mosoni, P.; Rochette, Y.; Chilliard, Y.; Doreau, M. Increasing linseed supply in dairy cow diets based on hay or corn silage: Effect on enteric methane emission, rumen microbial fermentation, and digestion. J. Dairy Sci. 2016, 99, 3445–3456. [Google Scholar] [CrossRef]
- Shingfield, K.J.; Ahvenjärvi, S.; Toivonen, V.; Vanhatalo, A.; Huhtanen, P.; Griinari, J.M. Effect of incremental levels of sunflower-seed oil in the diet on ruminal lipid metabolism in lactating cows. Br. J. Nutr. 2008, 99, 971–983. [Google Scholar] [CrossRef]
- Pirondini, M.; Colombini, S.; Mele, M.; Malagutti, L.; Rapetti, L.; Galassi, G.; Crovetto, G. Effect of dietary starch concentration and fish oil supplementation on milk yield and composition, diet digestibility, and methane emissions in lactating dairy cows. J. Dairy Sci. 2015, 98, 357–372. [Google Scholar] [CrossRef]
- Philippeau, C.; Lettat, A.; Martin, C.; Silberberg, M.; Morgavi, D.; Ferlay, A.; Berger, C.; Nozière, P. Effects of bacterial direct-fed microbials on ruminal characteristics, methane emission, and milk fatty acid composition in cows fed high-or low-starch diets. J. Dairy Sci. 2017, 100, 2637–2650. [Google Scholar] [CrossRef]
- Hatew, B.; Podesta, S.; Van Laar, H.; Pellikaan, W.; Ellis, J.; Dijkstra, J.; Bannink, A. Effects of dietary starch content and rate of fermentation on methane production in lactating dairy cows. J. Dairy Sci. 2015, 98, 486–499. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.S. Effects of diet on short-term regulation of feed intake by lactating dairy cattle. J. Dairy Sci. 2000, 83, 1598–1624. [Google Scholar] [CrossRef]
- Voelker, J.; Allen, M. Pelleted beet pulp substituted for high-moisture corn: 1. Effects on feed intake, chewing behavior, and milk production of lactating dairy cows. J. Dairy Sci. 2003, 86, 3542–3552. [Google Scholar] [CrossRef]
- Allen, M.; Bradford, B.; Oba, M. Board-invited review: The hepatic oxidation theory of the control of feed intake and its application to ruminants. J. Anim. Sci. 2009, 87, 3317–3334. [Google Scholar] [CrossRef]
- Beckman, J.; Weiss, W.P. Nutrient digestibility of diets with different fiber to starch ratios when fed to lactating dairy cows. J. Dairy Sci. 2005, 88, 1015–1023. [Google Scholar] [CrossRef]
- Kliem, K.E.; Humphries, D.J.; Kirton, P.; Givens, D.I.; Reynolds, C.K. Differential effects of oilseed supplements on methane production and milk fatty acid concentrations in dairy cows. Animal 2019, 13, 309–317. [Google Scholar] [CrossRef]
- Muñoz, C.; Sánchez, R.; Peralta, A.; Espíndola, S.; Yan, T.; Morales, R.; Ungerfeld, E. Effects of feeding unprocessed oilseeds on methane emission, nitrogen utilization efficiency and milk fatty acid profile of lactating dairy cows. Anim. Feed Sci. Technol. 2019, 249, 18–30. [Google Scholar] [CrossRef]
- Ramin, M.; Huhtanen, P. Development of equations for predicting methane emissions from ruminants. J. Dairy Sci. 2013, 96, 2476–2493. [Google Scholar] [CrossRef]
- Beauchemin, K.; Kreuzer, M.; O’mara, F.; McAllister, T. Nutritional management for enteric methane abatement: A review. Aust. J. Exp. Agric. 2008, 48, 21–27. [Google Scholar] [CrossRef]
- Beauchemin, K.; McGinn, S.; Benchaar, C.; Holtshausen, L. Crushed sunflower, flax, or canola seeds in lactating dairy cow diets: Effects on methane production, rumen fermentation, and milk production. J. Dairy Sci. 2009, 92, 2118–2127. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K. The effect of dietary fats on methane emissions, and its other effects on digestibility, rumen fermentation and lactation performance in cattle: A meta-analysis. Livest. Sci. 2013, 155, 244–254. [Google Scholar] [CrossRef]
- Maia, M.R.; Chaudhary, L.C.; Figueres, L.; Wallace, R.J. Metabolism of polyunsaturated fatty acids and their toxicity to the microflora of the rumen. Antonie Van Leeuwenhoek 2007, 91, 303–314. [Google Scholar] [CrossRef]
- Hobson, P.N.; Stewart, C.S. The Rumen Microbial Ecosystem; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Shelton, J.L.; Akob, D.M.; McIntosh, J.C.; Fierer, N.; Spear, J.R.; Warwick, P.D.; McCray, J.E. Environmental drivers of differences in microbial community structure in crude oil reservoirs across a methanogenic gradient. Front. Microbiol. 2016, 7, 1535. [Google Scholar] [CrossRef] [PubMed]
- Cieslak, A.; Miltko, R.; Belzecki, G.; Szumacher-Strabel, M.; Potkanski, A.; Kwiatkowska, E.; Michalowski, T. Effect of vegetable oils on the methane concentration and population density of the rumen ciliate, Eremoplastron dilobum, grown in vitro. J. Anim. Feed Sci. 2006, 15, 15. [Google Scholar] [CrossRef]
- Ormerod, K.L.; Wood, D.L.; Lachner, N.; Gellatly, S.L.; Daly, J.N.; Parsons, J.D.; Dal’Molin, C.G.; Palfreyman, R.W.; Nielsen, L.K.; Cooper, M.A. Genomic characterization of the uncultured Bacteroidales family S24-7 inhabiting the guts of homeothermic animals. Microbiome 2016, 4, 36. [Google Scholar] [CrossRef] [PubMed]
- Piao, H.; Lachman, M.; Malfatti, S.; Sczyrba, A.; Knierim, B.; Auer, M.; Tringe, S.G.; Mackie, R.I.; Yeoman, C.J.; Hess, M. Temporal dynamics of fibrolytic and methanogenic rumen microorganisms during in situ incubation of switchgrass determined by 16S rRNA gene profiling. Front. Microbiol. 2014, 5, 307. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Wang, Y.; Li, Y.; Cheng, Y.; Zhu, W. Temporal changes of the bacterial community colonizing wheat straw in the cow rumen. Anaerobe 2018, 50, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tapio, I.; Snelling, T.J.; Strozzi, F.; Wallace, R.J. The ruminal microbiome associated with methane emissions from ruminant livestock. J. Anim. Sci. Biotechnol. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Belanche, A.; de la Fuente, G.; Newbold, C.J. Effect of progressive inoculation of fauna-free sheep with holotrich protozoa and total-fauna on rumen fermentation, microbial diversity and methane emissions. FEMS Microbiol. Ecol. 2015, 91, fiu026. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Diet 1 | ||||
|---|---|---|---|---|
| LS | LSO | HS | HSO | |
| Inclusion rate (g/kg DM) | ||||
| Grass silage | 550 | 550 | 550 | 550 |
| Rolled barley | 0 | 0 | 85 | 77 |
| Ground wheat | 0 | 0 | 255 | 231.5 |
| Sugar beet pulp | 255 | 231.5 | 0 | 0 |
| Barley feed | 85 | 77 | 0 | 0 |
| Rapeseed meal, solvent-extracted | 80 | 80 | 80 | 80 |
| Urea | 0 | 1.5 | 0 | 1.5 |
| Oil mixture 2 | 0 | 30 | 0 | 30 |
| Vitamin and mineral Pre-mix 3 | 30 | 30 | 30 | 30 |
| Chemical composition (g/kg DM unless stated) | ||||
| DM (g/kg as fed) | 536 | 539 | 528 | 532 |
| OM | 926 | 928 | 941 | 942 |
| CP | 159 | 160 | 165 | 165 |
| FA | 23.5 | 51.4 | 22.9 | 50.8 |
| NDF | 430 | 417 | 344 | 339 |
| pdNDF | 348 | 337 | 276 | 271 |
| WSC | 44.8 | 42.3 | 27.5 | 26.6 |
| Starch | 16.1 | 15.0 | 202 | 184 |
| GE (MJ/kg DM) | 18.3 | 18.9 | 18.4 | 19.0 |
| Intake (kg/d Unless Stated) | Treatment 1 | SEM | p-Value 2 | |||||
|---|---|---|---|---|---|---|---|---|
| LS | LSO | HS | HSO | S | O | S × O | ||
| Silage DM | 13.1 | 11.9 | 13.4 | 11.9 | 0.26 | 0.36 | <0.001 | 0.56 |
| Concentrate DM | 9.9 | 8.5 | 10.1 | 8.5 | 0.19 | 0.57 | <0.001 | 0.35 |
| Oil | - | 0.60 | - | 0.61 | 0.006 | - | <0.001 | - |
| DM | 23.0 | 21.0 | 23.5 | 21.0 | 0.43 | 0.43 | <0.001 | 0.45 |
| OM | 20.7 | 18.9 | 21.5 | 19.2 | 0.39 | 0.087 | <0.001 | 0.38 |
| CP | 3.66 | 3.35 | 3.86 | 3.44 | 0.062 | 0.013 | <0.001 | 0.27 |
| NDF | 10.0 | 8.80 | 8.22 | 7.23 | 0.174 | <0.001 | <0.001 | 0.51 |
| pdNDF | 8.06 | 7.11 | 6.58 | 5.78 | 0.140 | <0.001 | <0.001 | 0.49 |
| WSC | 1.01 | 0.87 | 0.64 | 0.55 | 0.027 | <0.001 | 0.007 | 0.43 |
| Starch | 0.36 | 0.31 | 4.57 | 3.72 | 0.036 | <0.001 | <0.001 | <0.001 |
| GE intake (MJ/d) | 421 | 396 | 434 | 400 | 7.8 | 0.18 | 0.002 | 0.42 |
| FA intake (g/d) | ||||||||
| 12:0 | 0.44 | 0.74 | 0.46 | 0.75 | 0.012 | 0.12 | <0.001 | 0.63 |
| 14:0 | 2.1 | 16.1 | 2.0 | 16.1 | 0.21 | 0.93 | <0.001 | 0.89 |
| 16:0 | 88 | 131 | 86 | 128 | 2.2 | 0.067 | <0.001 | 0.81 |
| cis-9 16:1 | 3.2 | 18.1 | 3.3 | 18.2 | 0.24 | 0.78 | <0.001 | 0.95 |
| 18:0 | 7.0 | 27.3 | 7.3 | 27.5 | 0.37 | 0.42 | <0.001 | 0.95 |
| cis-9 18:1 | 74 | 188 | 73 | 186 | 2.7 | 0.38 | <0.001 | 0.95 |
| cis-11 18:1 | 13.1 | 20.0 | 13.0 | 19.8 | 0.34 | 0.47 | <0.001 | 0.75 |
| cis-9, cis-12 18:2 | 167 | 373 | 171 | 375 | 5.4 | 0.41 | <0.001 | 0.73 |
| 20:0 | 5.85 | 6.83 | 5.85 | 6.77 | 0.136 | 0.70 | <0.001 | 0.69 |
| 18:3n–3 3 | 145 | 136 | 145 | 134 | 3.1 | 0.65 | 0.003 | 0.64 |
| 22:0 | 4.31 | 6.90 | 4.07 | 6.67 | 0.114 | 0.015 | <0.001 | 0.99 |
| cis-11 22:1 | 0.02 | 2.17 | 0.01 | 2.17 | 0.028 | 0.91 | <0.001 | 0.82 |
| 20:5n–3 | 0.0 | 31.5 | 0.0 | 31.6 | 0.41 | 0.85 | <0.001 | 0.85 |
| 24:0 | 3.61 | 4.18 | 3.00 | 3.63 | 0.076 | <0.001 | <0.001 | 0.55 |
| 22:5n–3 | 0.00 | 3.63 | 0.00 | 3.64 | 0.048 | 0.85 | <0.001 | 0.85 |
| 26:0 | 2.92 | 2.76 | 3.04 | 2.81 | 0.064 | 0.072 | 0.002 | 0.48 |
| 22:6n–3 | 0.0 | 20.2 | 0.0 | 20.3 | 0.26 | 0.85 | <0.001 | 0.85 |
| Unidentified | 0.92 | 2.90 | 1.09 | 3.04 | 0.041 | 0.002 | <0.001 | 0.59 |
| Σ 25–30 | 11.7 | 10.7 | 11.9 | 10.8 | 0.25 | 0.37 | 0.001 | 0.56 |
| SFA | 127 | 211 | 124 | 207 | 3.4 | 0.10 | <0.001 | 0.85 |
| MUFA | 118 | 267 | 113 | 263 | 3.9 | 0.11 | <0.001 | 0.98 |
| PUFA | 294 | 568 | 300 | 570 | 8.8 | 0.43 | <0.001 | 0.72 |
| Total FA | 539 | 1049 | 537 | 1042 | 16.0 | 0.69 | <0.001 | 0.81 |
| Treatment 1 | SEM | p-Value 2 | ||||||
|---|---|---|---|---|---|---|---|---|
| LS | LSO | HS | HSO | S | O | S × O | ||
| Yield | ||||||||
| Milk (kg/d) | 30.9 | 31.3 | 31.1 | 30.4 | 1.55 | 0.73 | 0.85 | 0.55 |
| ECM (kg/d) | 30.2 | 30.0 | 31.3 | 27.8 | 1.87 | 0.58 | 0.11 | 0.14 |
| Fat (g/d) | 1242 | 1230 | 1266 | 1093 | 85.2 | 0.26 | 0.087 | 0.12 |
| Protein (g/d) | 1021 | 981 | 1079 | 924 | 51.0 | 0.99 | 0.030 | 0.15 |
| Lactose (g/d) | 1322 | 1374 | 1383 | 1358 | 101.3 | 0.71 | 0.82 | 0.52 |
| Milk composition (g/kg) | ||||||||
| Fat | 40.1 | 39.4 | 40.5 | 35.8 | 1.16 | 0.13 | 0.024 | 0.072 |
| Protein | 33.1 | 31.5 | 34.7 | 30.6 | 0.94 | 0.68 | 0.008 | 0.15 |
| Lactose | 42.7 | 43.9 | 44.3 | 44.3 | 1.45 | 0.35 | 0.54 | 0.52 |
| ECM/DMI | 1.31 | 1.43 | 1.33 | 1.32 | 0.075 | 0.42 | 0.31 | 0.28 |
| Treatment 1 | SEM | p-Value 2 | ||||||
|---|---|---|---|---|---|---|---|---|
| LS | LSO | HS | HSO | S | O | S × O | ||
| Digestibility (g/kg or otherwise stated) | ||||||||
| DM | 691 | 699 | 712 | 720 | 3.1 | 0.001 | 0.13 | 0.98 |
| OM | 706 | 714 | 726 | 733 | 3.4 | 0.002 | 0.15 | 0.96 |
| CP | 674 | 682 | 682 | 719 | 7.3 | 0.063 | 0.058 | 0.20 |
| NDF | 608 | 615 | 562 | 563 | 5.9 | <0.001 | 0.61 | 0.76 |
| pdNDF | 751 | 761 | 701 | 704 | 7.2 | <0.001 | 0.55 | 0.76 |
| Starch | 858 | 805 | 927 | 928 | 14.3 | 0.002 | 0.20 | 0.18 |
| GE (kJ/MJ) | 680 | 692 | 699 | 715 | 3.4 | 0.003 | 0.019 | 0.66 |
| Treatment 1 | SEM | p-Value 2 | ||||||
|---|---|---|---|---|---|---|---|---|
| LS | LSO | HS | HSO | S | O | S × O | ||
| pH | 6.79 | 6.68 | 6.64 | 6.67 | 0.061 | 0.24 | 0.56 | 0.32 |
| Ammonia N (mmol/L) | 5.60 | 6.16 | 7.67 | 9.17 | 0.306 | <0.001 | <0.001 | 0.027 |
| Total VFA (mmol/L) | 100 | 100 | 110 | 102 | 2.6 | 0.056 | 0.21 | 0.18 |
| Molar proportions (mmol/mol) | ||||||||
| Acetate | 681 | 681 | 653 | 637 | 5.8 | <0.001 | 0.23 | 0.20 |
| Propionate | 179 | 181 | 186 | 203 | 9.8 | 0.18 | 0.37 | 0.50 |
| Butyrate | 99.3 | 101 | 111 | 112 | 4.41 | 0.016 | 0.77 | 0.96 |
| Isobutyrate | 7.77 | 6.97 | 9.41 | 9.09 | 0.695 | 0.014 | 0.35 | 0.67 |
| Valerate | 15.3 | 14.1 | 17.5 | 17.5 | 0.47 | <0.001 | 0.28 | 0.21 |
| Isovalerate | 11.4 | 9.81 | 15.2 | 14.4 | 1.76 | 0.004 | 0.26 | 0.68 |
| Caproate | 6.93 | 6.35 | 8.25 | 7.85 | 0.544 | 0.034 | 0.38 | 0.87 |
| Molar ratio | ||||||||
| Acetate:Propionate | 3.81 | 3.77 | 3.56 | 3.17 | 0.193 | 0.070 | 0.31 | 0.41 |
| Treatment 1 | SEM | p-Value 2 | ||||||
|---|---|---|---|---|---|---|---|---|
| LS | LSO | HS | HSO | S | O | S × O | ||
| Enteric CH4 | ||||||||
| g/d | 551 | 471 | 553 | 478 | 39.4 | 0.89 | 0.051 | 0.93 |
| g/kg OMI | 26.8 | 25.0 | 25.9 | 25.0 | 1.95 | 0.79 | 0.46 | 0.80 |
| g/kg OMD | 38.2 | 35.1 | 36.9 | 33.1 | 2.53 | 0.46 | 0.15 | 0.87 |
| g/kg milk | 17.8 | 15.1 | 17.8 | 15.9 | 0.74 | 0.64 | 0.015 | 0.58 |
| g/kg ECM | 18.2 | 15.7 | 17.8 | 17.4 | 0.70 | 0.37 | 0.067 | 0.16 |
| % of GEI | 7.25 | 6.56 | 7.04 | 6.59 | 0.521 | 0.86 | 0.26 | 0.80 |
| Enteric CO2 | ||||||||
| g/d | 6109 | 4488 | 4937 | 4846 | 544.8 | 0.48 | 0.16 | 0.21 |
| g/kg OMD | 424 | 335 | 330 | 336 | 34.7 | 0.23 | 0.27 | 0.22 |
| g/kg milk | 198 | 144 | 159 | 158 | 12.8 | 0.38 | 0.074 | 0.086 |
| g/kg ECM | 202 | 149 | 160 | 173 | 12.2 | 0.46 | 0.15 | 0.036 |
| Diversity | Treatment 1 | SEM | p-Value 2 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Estimate | LS | LSO | HS | HSO | S | O | S × O | ||
| Bacteria | Shannon | 2.21 | 2.22 | 2.18 | 2.02 | 0.104 | 0.23 | 0.45 | 0.38 |
| Simpson | 0.728 | 0.732 | 0.738 | 0.662 | 0.035 | 0.29 | 0.21 | 0.17 | |
| Richness | 68.5 | 68.4 | 63.3 | 62.3 | 2.019 | 0.014 | 0.73 | 0.78 | |
| Evenness | 0.519 | 0.524 | 0.522 | 0.486 | 0.022 | 0.36 | 0.42 | 0.30 | |
| Archaea | Shannon | 1.17 | 1.18 | 0.96 | 1.01 | 0.135 | 0.13 | 0.79 | 0.86 |
| Simpson | 0.594 | 0.633 | 0.515 | 0.536 | 0.059 | 0.15 | 0.60 | 0.87 | |
| Richness | 6.81 | 6.33 | 7.69 | 5.61 | 0.723 | 0.89 | 0.064 | 0.21 | |
| Evenness | 0.581 | 0.589 | 0.437 | 0.545 | 0.056 | 0.10 | 0.28 | 0.34 | |
| Ciliate protozoa | Shannon | 2.15 | 2.26 | 2.23 | 2.12 | 0.088 | 0.80 | 0.85 | 0.22 |
| Simpson | 0.840 | 0.854 | 0.855 | 0.836 | 0.009 | 0.89 | 0.74 | 0.071 | |
| Richness | 25.9 | 27.7 | 26.0 | 27.5 | 1.626 | 0.98 | 0.31 | 0.93 | |
| Evenness | 0.628 | 0.648 | 0.663 | 0.613 | 0.014 | 0.97 | 0.24 | 0.022 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Darabighane, B.; Tapio, I.; Ventto, L.; Kairenius, P.; Stefański, T.; Leskinen, H.; Shingfield, K.J.; Vilkki, J.; Bayat, A.-R. Effects of Starch Level and a Mixture of Sunflower and Fish Oils on Nutrient Intake and Digestibility, Rumen Fermentation, and Ruminal Methane Emissions in Dairy Cows. Animals 2021, 11, 1310. https://doi.org/10.3390/ani11051310
Darabighane B, Tapio I, Ventto L, Kairenius P, Stefański T, Leskinen H, Shingfield KJ, Vilkki J, Bayat A-R. Effects of Starch Level and a Mixture of Sunflower and Fish Oils on Nutrient Intake and Digestibility, Rumen Fermentation, and Ruminal Methane Emissions in Dairy Cows. Animals. 2021; 11(5):1310. https://doi.org/10.3390/ani11051310
Chicago/Turabian StyleDarabighane, Babak, Ilma Tapio, Laura Ventto, Piia Kairenius, Tomasz Stefański, Heidi Leskinen, Kevin J. Shingfield, Johanna Vilkki, and Ali-Reza Bayat. 2021. "Effects of Starch Level and a Mixture of Sunflower and Fish Oils on Nutrient Intake and Digestibility, Rumen Fermentation, and Ruminal Methane Emissions in Dairy Cows" Animals 11, no. 5: 1310. https://doi.org/10.3390/ani11051310
APA StyleDarabighane, B., Tapio, I., Ventto, L., Kairenius, P., Stefański, T., Leskinen, H., Shingfield, K. J., Vilkki, J., & Bayat, A.-R. (2021). Effects of Starch Level and a Mixture of Sunflower and Fish Oils on Nutrient Intake and Digestibility, Rumen Fermentation, and Ruminal Methane Emissions in Dairy Cows. Animals, 11(5), 1310. https://doi.org/10.3390/ani11051310