1. Introduction
Poultry producers have been preferentially using cost-effective antibiotics in large quantities since 1951 [
1] to improve growth performance and prevent diseases. However, the development of microbial resistance to these antibiotics causes health issues in the food chain from animals to humans. Consequently, the European Union has banned the use of antibiotic growth promoters in poultry diets since 2006 [
2], thus motivating researchers to seek effective alternatives such as probiotics, prebiotics, herbal products, marine natural products, and organic acids [
3]. One such potential natural alternative is seaweed, from which natural marine products that contain various biologically active components can be derived [
4], as well as useful ingredients with abundant health benefits [
5]. Since the 20th century, the consumption of seaweed (marine macroalgae) products has been increasing among people seeking to improve their health and diets. Holdt and Kraan [
6] reported in 2011 that seaweeds contain various types of soluble fiber such as agar, carrageenan, and alginate, which are not present in terrestrial plants. Moreover, seaweeds are rich in nutrients, contain ample proteins, lipids, polyphenols, and polysaccharides, and possess antiviral and antifungal properties. Particularly, bioactive substances have been found in red seaweeds [
7].
Seaweeds are considered a prime feed constituent in Norway, after the processing of seaweed-based meal from kelp started in the 1960s [
8]. Seaweeds are preferentially used in livestock feed, as they contain nutrients such as chelating micro-minerals at higher concentrations than can be found in typical inorganic feed components. Additionally, they are rich in prebiotic complex carbohydrates, pigments, and polyunsaturated fatty acids, which can improve the health of consumers [
9]. Red seaweeds are especially abundant in dietary fiber, vitamins, minerals, carotenoids, phlorotannins, amino acids, and other fitness-enhancing constituents and, thus, contribute raw materials to the pharmaceutical and nutraceutical industries [
6]. The polysaccharides in seaweed make it an ideal prebiotic due to their potential for improving the gut microbiota [
10]. The marine red seaweed
Palmaria palmata (L.), otherwise known as red dulse (formerly known as
Halymenia palmata), has smooth leathery fronds and is deep red in autumn and greenish/yellow in summer. Individuals can grow up to 30–40 cm in length. This species occurs in the North Atlantic and is found in moderately exposed to exposed shores in areas subject to tidal currents. Furthermore, as
P. palmata is rich in protein, it was harvested for food and given as feed to livestock such as sheep and goats in Gotland (Sweden) and cows in Brittany (France) during the 19th and early 20th centuries [
11].
Regarding poultry production, seaweed may be able to improve the immune status of birds and reduce pathogenic microbes in the digestive tract [
12]. Previous studies have reported promising results in terms of animal growth and health with the use of seaweed dietary supplements [
13,
14]. Moreover, Dierick et al. [
13] suggested that a brown seaweed (
Ascophyllum nodosum) can be a natural replacement for antibiotics. Montserrat and Goñi [
15] evaluated the effects of red (nori,
Porphyra ternera) and brown seaweeds (wakame,
Undaria pinnatifida) and reported alterations in microbial activities in rats. According to Hoebler et al. [
16], the addition of 5% brown seaweed (
Laminaria digitate) to the diet of pigs increased propionic and butyric acid concentrations in the large intestine. Furthermore, Kulshreshtha et al. [
10] showed that red seaweeds (
Chondrus crispus and
Sarcodiotheca gaudichaudii) greatly improved the growth performance, egg efficiency, and overall gut health of laying hens. The dietary inclusion of seaweed also was shown to improve bird health and feed efficiency by increasing the abundance of beneficial gut bacteria and strengthening the innate host immune system [
17]. Each type of seaweed has its own properties, so specific research on
P. palmata needs to be conducted [
9]. Nevertheless, based on previous studies, the application of seaweed should have positive effects on livestock. To date, no experiments on seaweed supplementation in the broiler diet have been initiated. Therefore, the aims of the present study were to evaluate the effects of adding marine red seaweed (
P. palmata) to a soybean meal (SBM) based diet on the growth performance, total tract nutrient digestibility, excreta gas emissions, meat quality, intestine histomorphology, and fecal microbial counts of broiler chickens.
3. Results
Broiler FI improved linearly (
p = 0.0369) with seaweed dietary supplementation from d 14 to 28 (
Table 3). The BWG tended to be greater from d 14 to 28 (
p = 0.087) and d 28 to 42 (
p = 0.082) in the seaweed-supplemented groups, with a linear increase (
p = 0.0075) observed over the entire experimental period. Additionally, seaweed supplementation was linearly associated with the FCR from d 28 to 42 (
p = 0.028) and over the entire experimental period (
p = 0.049). Quadratic effects were not observed for any performance-related variable. The level of seaweed supplementation did not affect any of the blood-related traits after week 4. However, linear increases in albumin, creatinine, uric acid, and the WBC count due to seaweed supplementation were observed over the duration of the experiment (
Table 4).
Nutrient digestibility was not influenced by seaweed supplementation, as observed on d 14 and 28. However, the digestibility coefficients of dry matter (DM;
p = 0.037) exhibited a linear improvement in broilers fed seaweed-supplemented diets on d 42, but no significant effects were observed for GE digestibility (
Table 5). Dietary supplementation with seaweed led to a linear increase in
Lactobacillus sp. (
p = 0.035) counts, whereas
E. coli (
p = 0.027) counts were decreased on d 42 (
Table 6). To contrast, no significant effects on
Salmonella sp. counts were observed throughout the entire experiment (
Table 6). Dietary supplementation with seaweed led to linear decreases in NH
3 and H
2S emissions on d 42. Conversely, no effects on total mercaptan, acetic acid, propionate, or butyrate were observed over the entire experiment (
Table 7). The morphometric measurements of the chicken intestines can be found in
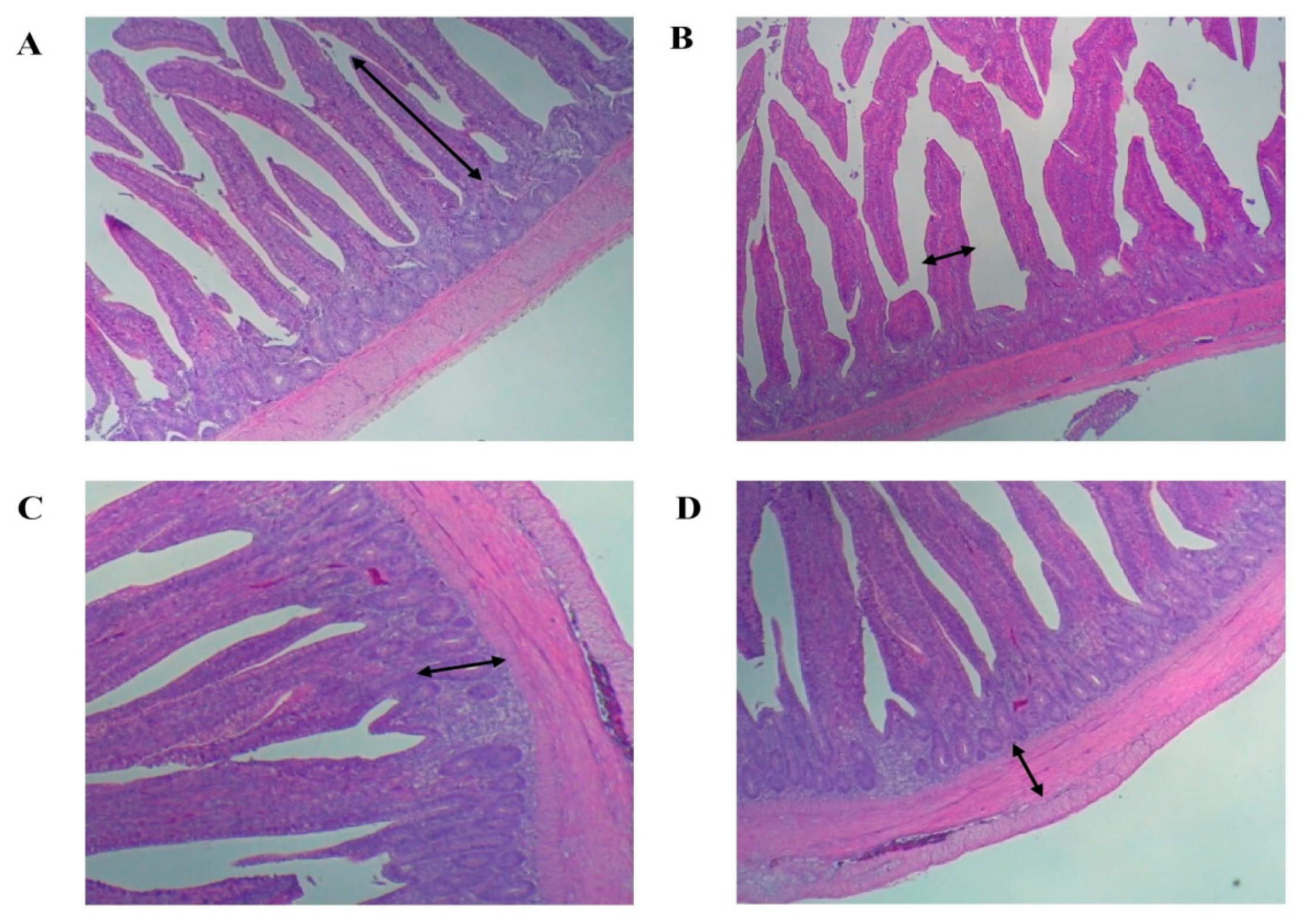
Table 8, with these measurements represented in
Figure 1. The dietary inclusion of seaweed influenced the morphology of the mucosa in the small intestine. Considering the jejunum, the average villus height and width were significantly enhanced in broilers fed seaweed-supplemented diets compared to the control broilers.
The crypt depth and villus:crypt ratio appeared to be markedly influenced by seaweed supplementation, but the differences in values among treatments were not statistically significant (
Table 8). The WHC (
p = 0.0511) and cooking loss (
p = 0.0129) decreased linearly with increasing levels of seaweed supplementation. A linear decrease in drip loss also was observed on d 7 (
p = 0.0318). Additionally, the relative weights of the breast muscle (
p = 0.033), bursa of Fabricius (
p = 0.049), and gizzard (
p = 0.018) increased linearly with increasing levels of seaweed supplementation (
Table 9).
4. Discussion
Seaweeds or macroalgae are a good source of polysaccharides, which include complex carbohydrates that cannot be hydrolyzed in the upper gastrointestinal tract. Seaweeds, thus, are considered a healthy source of dietary fiber [
24]. The polysaccharides derived from seaweeds also are considered potential prebiotics and cannot be digested in the small intestine, but they can survive bacterial fermentation in the large intestine and, hence, affect the intestinal microbiota [
14]. Abudabos et al. [
25] reported that dietary supplementation with seaweed in broilers resulted in decreased microbial abundance in the digestive tract and enhanced immune status, thereby improving the meat quality of the broilers. During the current study, dietary supplementation with red seaweed improved the growth performance of broilers in a linear manner, consistent with the findings of Wang et al. [
26], who reported that broilers fed a seaweed-supplemented diet exhibited improvements in body weight. Similarly, Choi et al. [
27] reported significant improvements in the BWG and feed intake of broilers upon supplementation with fermented seaweed; however, the feed efficiency was not affected.
Regarding terms of feed development in broiler production, the focus has expanded to include potential supplementation with natural products. Based on our results, the inclusion of seaweed in a corn/soybean meal-based diet led to significant increases in weight gain and feed efficiency, similar to the findings of Shi et al. [
28]. Evans and Critchley [
9] also showed that the dietary inclusion of seaweed enhanced bird health and productivity and improved the gut microflora. These benefits may be due to the immune-enhancing effect of red seaweeds. Concerning terms of their functional properties, polysaccharides such as fucans and alginic acid by-products in seaweeds exhibit anticoagulant, anti-inflammatory, antiviral, and antitumor activities [
29]. Accordingly, dietary supplementation with seaweed has beneficial effects on broilers [
30]. Our results also showed that the broilers fed red seaweed-supplemented diets exhibited improved nutrient digestibility, especially of DM. However, Nhlane et al. [
31] obtained opposing results, with the nutrient digestibility in broilers apparently unaffected by dietary supplementation with seaweed. To date, only a few studies have been conducted on whether the dietary inclusion of seaweed improves nutrient digestibility in broilers. Improved nutrient digestibility is associated with better absorption of feed nutrients in birds as the feed passes through the digestive tract. Despite the conflicting results, we found that the villus height in the small intestine of supplemented birds was significantly taller, which is correlated with more efficient nutrient absorption and improved growth performance. Overall, we believe that dietary supplementation with seaweeds can improve the growth performance of, and nutrient digestibility in, broiler chickens by strengthening the intestinal integrity and immune system.
One of the goals of the present study was to determine whether dietary supplementation with seaweed would improve the blood profile of broiler chickens, as assessed based on levels of albumin, creatinine, uric acid, and WBC. Kang et al. [
32] found that animals fed a dietary seaweed extract exhibited decreased oxidative stress, as shown by concentrations of glutamic oxaloacetic transaminase and glutamic pyruvic transaminase. To contrast, Lokaewmanee et al. [
33] reported that supplementation with fermented brown seaweed had no significant effects on the blood profile of broilers. Research on the effects of seaweed supplementation on poultry blood profiles is limited, so we could not perform a comparative analysis. The conflicting results from previous experiments could have been due to variation in the seaweed dosage and differences in the composition of the additive of interest and diets. White and Venkatesh [
34] determined that total cholesterol and triglycerides play major roles in humans, as they participate in animal cell membrane formation, fat transfer, and steroid hormone synthesis, and are correlated with blood sugar in the kidney.
During a recent study, Balasubramanian [
35] demonstrated that gut health is a major factor regulating bird growth performance and, hence, affects the economics of poultry production. It also was noted that the intestinal microflora profile plays an important role in gut health. During the present study, dietary supplementation with red seaweed increased the
Lactobacillus sp. count and decreased the
E. coli count in the host gut. This finding agreed with the result of Bai et al. [
36], who found decreased
E. coli and increased
Lactobacillus sp. counts in broiler chickens fed seaweed-supplemented diets. Earlier reports from Wang et al. [
37] and Leonard et al. [
38] indicated that
Laminaria japonica powder could act as a potential medicine that enhances host resistance against gastrointestinal tract pathogens in weaner pigs by strengthening their antibody immune function. The powder also helped with controlling gastrointestinal function and promoting pig growth. Cecropins can destroy gram-positive and gram-negative bacteria in the intestinal tract by permeabilizing bacterial membranes [
39]. Based on our results, we assume a similar beneficial effect from seaweed on gut microflora in poultry. Upon seaweed supplementation, seaweed components usually adhere to intestinal mucus and epithelial cells, and this attachment is believed to be key to their immunomodulatory effects [
15].
Numerous studies have explored dietary manipulation strategies to lessen the environmental hazards from emissions of noxious gases such as NH
3, H
2S, and methyl mercaptan [
40]. According to Yan and Kim [
41], fecal noxious gas emission is associated with nutrient digestibility, with decreased digestibility resulting in reductions in the amount of microbial fermentation substrates in the large intestine and fecal noxious gas emission. Considering the present research, dietary supplementation with red seaweed significantly decreased NH
3 and H
2S concentrations but did not have significant effects on the concentrations of total mercaptan, acetic acid, propionate, and butyrate in fecal gas emissions. Notably, Wang et al. [
42] stated that NH
3, H
2S, and methyl mercaptan affect the air quality and their emissions are a major problem in livestock production.
According to Offer and Knight [
43], the most significant meat quality characteristic is the WHC. During the present study, the WHC tended to decrease with the level of seaweed supplementation, with a significant linear decrease with seaweed supplementation observed for drip loss on d 7, and linear associations were found for the relative weights of the breast muscle, bursa of Fabricius, and gizzard. Hossain et al. [
44] reported that meat pH is commonly considered a direct indicator of the consistency of the muscle acid content, whereas drip loss is an indicator of meat quality [
45]. Choi et al. [
27] also found that organ weight was significantly influenced by fermented seaweed supplementation. To our knowledge, this is the first study on the meat quality of broilers fed diets supplemented with the seaweed
P. palmata. Our results showed that the dietary inclusion of
P. palmata had beneficial effects on the meat quality of broiler chickens, in terms of the WHC, drip loss, and organ weights. Nevertheless, antinutritional factors also may contribute to an increase in organ weights. It appears that seaweed supplementation might have improved the FCR and gut health (based on increases in villus height and width), thus resulting in a greater BWG compared to broilers fed the control diet. However, additional research is needed to verify our results.


 ,
, 




{kind=link}